
13.2 : Bases chromosomiques des maladies héréditaires
- Page ID
- 189669

Compétences à développer
- Décrire comment un caryogramme est créé
- Expliquer comment la non-disjonction entraîne des troubles du nombre de chromosomes
- Comparez les troubles causés par l'aneuploïdie
- Décrire comment les erreurs dans la structure des chromosomes se produisent lors d'inversions et de translocations
Des troubles héréditaires peuvent survenir lorsque les chromosomes se comportent anormalement au cours de la méiose. Les troubles chromosomiques peuvent être divisés en deux catégories : les anomalies du nombre de chromosomes et les réarrangements structuraux chromosomiques. Étant donné que même de petits segments de chromosomes peuvent couvrir de nombreux gènes, les troubles chromosomiques sont typiquement dramatiques et souvent mortels.
Identification des chromosomes
L'isolement et l'observation microscopique des chromosomes constituent la base de la cytogénétique et constituent la principale méthode par laquelle les cliniciens détectent les anomalies chromosomiques chez l'homme. Un caryotype est le nombre et l'apparence des chromosomes, y compris leur longueur, leur motif de bandes et la position du centromère. Pour obtenir une vue du caryotype d'un individu, les cytologues photographient les chromosomes, puis copient et collent chaque chromosome dans un tableau, ou caryogramme, également connu sous le nom d'idéogramme (Figure\(\PageIndex{1}\)).
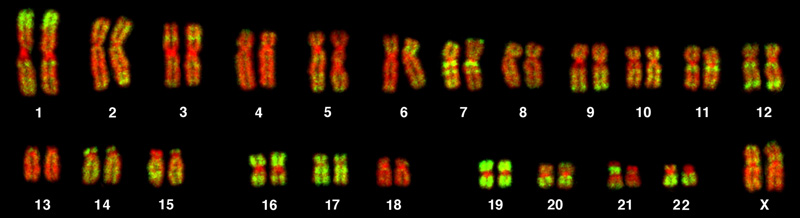
Chez une espèce donnée, les chromosomes peuvent être identifiés par leur nombre, leur taille, leur position centromérique et leur structure de bandes. Dans un caryotype humain, les autosomes ou « chromosomes corporels » (tous les chromosomes non sexuels) sont généralement organisés par ordre de taille approximatif, du plus grand (chromosome 1) au plus petit (chromosome 22). Les chromosomes X et Y ne sont pas des autosomes. Cependant, le chromosome 21 est en fait plus court que le chromosome 22. Cela a été découvert après la désignation du syndrome de Down sous le nom de trisomie 21, reflétant la façon dont cette maladie résulte de la possession d'un chromosome 21 supplémentaire (trois au total). Ne voulant pas changer le nom de cette maladie importante, le chromosome 21 a conservé sa numérotation, bien qu'il ait décrit l'ensemble de chromosomes le plus court. Les « bras » chromosomiques faisant saillie de chaque extrémité du centromère peuvent être désignés comme courts ou longs, selon leur longueur relative. Le bras court est abrégé p (pour « petite »), tandis que le bras long est abrégé q (car il suit « p » par ordre alphabétique). Chaque bras est ensuite subdivisé et désigné par un numéro. À l'aide de ce système de dénomination, les localisations sur les chromosomes peuvent être décrites de manière cohérente dans la littérature scientifique.
Lien professionnel : les généticiens utilisent des caryogrammes pour identifier les aberrations chromosomiques
Bien que Mendel soit considéré comme le « père de la génétique moderne », il a réalisé ses expériences avec aucun des outils que les généticiens d'aujourd'hui utilisent régulièrement. L'une de ces techniques cytologiques puissantes est le caryotypage, une méthode qui permet d'identifier les traits caractérisés par des anomalies chromosomiques à partir d'une seule cellule. Pour observer le caryotype d'une personne, ses cellules (comme les globules blancs) sont d'abord prélevées à partir d'un échantillon de sang ou d'un autre tissu. En laboratoire, les cellules isolées sont stimulées pour commencer à se diviser activement. Une substance chimique appelée colchicine est ensuite appliquée sur les cellules pour arrêter les chromosomes condensés en métaphase. Les cellules sont ensuite amenées à gonfler à l'aide d'une solution hypotonique afin que les chromosomes se dispersent. Enfin, l'échantillon est conservé dans un fixateur et appliqué sur une lame.
Le généticien colore ensuite les chromosomes avec l'un des nombreux colorants afin de mieux visualiser les motifs de bandes distincts et reproductibles de chaque paire de chromosomes. Après coloration, les chromosomes sont examinés au microscope à fond clair. La teinture Giemsa est un choix de teinture courant. La coloration de Giemsa produit environ 400 à 800 bandes (d'ADN étroitement enroulé et de protéines condensées) disposées le long des 23 paires de chromosomes ; un généticien expérimenté peut identifier chaque bande. Outre les modèles de bandes, les chromosomes sont identifiés en fonction de leur taille et de leur localisation centromérique. Pour obtenir la représentation classique du caryotype dans laquelle les paires homologues de chromosomes sont alignées par ordre numérique du plus long au plus court, le généticien obtient une image numérique, identifie chaque chromosome et organise manuellement les chromosomes selon ce schéma (Figure\(\PageIndex{1}\)).
Dans sa forme la plus élémentaire, le caryogramme peut révéler des anomalies génétiques dans lesquelles un individu possède trop ou trop peu de chromosomes par cellule. Le syndrome de Down, identifié par une troisième copie du chromosome 21, et le syndrome de Turner, caractérisé par la présence d'un seul chromosome X chez la femme au lieu des deux normaux, en sont des exemples. Les généticiens peuvent également identifier de grandes délétions ou insertions d'ADN. Par exemple, le syndrome de Jacobsen, qui implique des traits faciaux distinctifs ainsi que des malformations cardiaques et hémorragiques, est identifié par une délétion sur le chromosome 11. Enfin, le caryotype peut identifier les translocations, qui se produisent lorsqu'un segment du matériel génétique se brise d'un chromosome et se rattache à un autre chromosome ou à une partie différente du même chromosome. Les translocations sont impliquées dans certains cancers, notamment la leucémie myéloïde chronique.
Du vivant de Mendel, l'héritage était un concept abstrait qui ne pouvait être déduit qu'en effectuant des croisements et en observant les traits exprimés par la progéniture. En observant un caryogramme, les généticiens d'aujourd'hui peuvent visualiser la composition chromosomique d'un individu afin de confirmer ou de prédire des anomalies génétiques chez les enfants, même avant la naissance.
Troubles du nombre de chromosomes
De tous les troubles chromosomiques, les anomalies du nombre de chromosomes sont celles qui sont le plus clairement identifiables à partir d'un caryogramme. Les troubles du nombre de chromosomes incluent la duplication ou la perte de chromosomes entiers, ainsi que des modifications du nombre d'ensembles complets de chromosomes. Elles sont causées par la non-disjonction, qui se produit lorsque des paires de chromosomes homologues ou de chromatides sœurs ne se séparent pas au cours de la méiose. Une synapse mal alignée ou incomplète, ou un dysfonctionnement de l'appareil fusiforme qui facilite la migration des chromosomes, peut entraîner la non-disjonction. Le risque de non-disjonction augmente avec l'âge des parents.
La non-disjonction peut survenir au cours de la méiose I ou II, avec des résultats différents (Figure\(\PageIndex{2}\)). Si les chromosomes homologues ne se séparent pas au cours de la méiose I, il en résulte deux gamètes dépourvus du chromosome en question et deux gamètes comportant deux copies du chromosome. Si les chromatides sœurs ne se séparent pas pendant la méiose II, il en résulte un gamète dépourvu de ce chromosome, deux gamètes normaux avec une copie du chromosome et un gamète avec deux copies du chromosome.
Art Connection
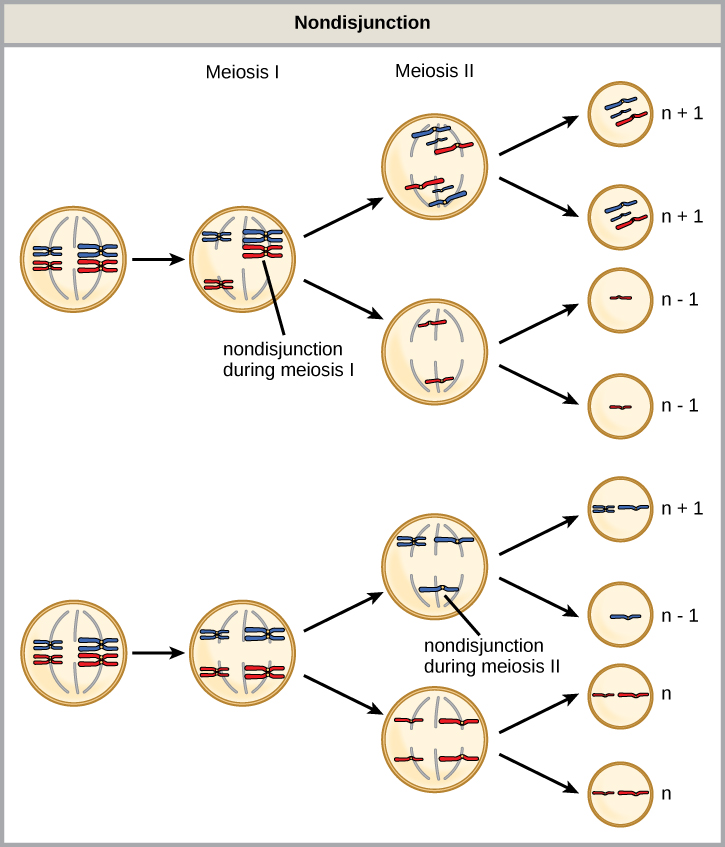
Laquelle des affirmations suivantes concernant la non-disjonction est vraie ?
- La non-disjonction ne produit que des gamètes avec des chromosomes n+1 ou n—1.
- L'absence de disjonction pendant la méiose II entraîne la production de 50 % de gamètes normaux.
- L'absence de disjonction au cours de la méiose I entraîne la production de 50 % de gamètes normaux.
- La non-disjonction donne toujours naissance à quatre types de gamètes différents.
Aneuploïdie
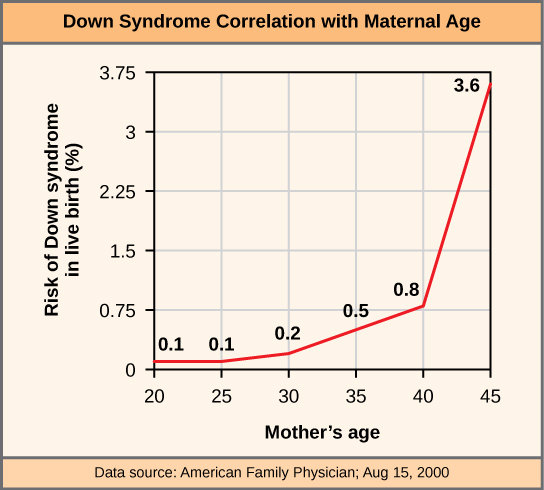
Un individu possédant le nombre approprié de chromosomes pour son espèce est appelé euploïde ; chez l'homme, l'euploïdie correspond à 22 paires d'autosomes et à une paire de chromosomes sexuels. Un individu présentant une erreur dans le nombre de chromosomes est décrit comme aneuploïde, terme qui inclut la monosomie (perte d'un chromosome) ou la trisomie (gain d'un chromosome étranger). Les zygotes humains monosomiques dépourvus d'une copie d'un autosome ne parviennent toujours pas à se développer jusqu'à la naissance en raison de leur absence de gènes essentiels. Cela souligne l'importance du « dosage génétique » chez l'homme. La plupart des trisomies autosomiques ne se développent pas non plus à la naissance ; toutefois, la duplication de certains des plus petits chromosomes (13, 15, 18, 21 ou 22) peut donner naissance à des descendants qui survivent de plusieurs semaines à de nombreuses années. Les personnes trisomiques souffrent d'un autre type de déséquilibre génétique : un excès de dose génique. Les individus possédant un chromosome supplémentaire peuvent synthétiser une grande quantité de produits géniques codés par ce chromosome. Cette dose supplémentaire (150 %) de gènes spécifiques peut entraîner un certain nombre de problèmes fonctionnels et empêche souvent le développement. La trisomie la plus fréquente chez les naissances viables est celle du chromosome 21, qui correspond au syndrome de Down. Les personnes atteintes de cette maladie héréditaire se caractérisent par une petite taille et des doigts rabougris, des distinctions faciales, notamment un crâne large et une langue volumineuse, et des retards de développement importants. L'incidence du syndrome de Down est corrélée à l'âge de la mère ; les femmes âgées sont plus susceptibles de tomber enceintes avec des fœtus porteurs du génotype de la trisomie 21 (Figure\(\PageIndex{3}\)).
Lien vers l'apprentissage
Visualisez l'ajout d'un chromosome qui mène au syndrome de Down dans cette simulation vidéo.
Polyploïdie
Un individu dont le nombre de chromosomes est supérieur au nombre correct (deux pour les espèces diploïdes) est appelé polyploïde. Par exemple, la fécondation d'un ovule diploïde anormal avec un spermatozoïde haploïde normal produirait un zygote triploïde. Les animaux polyploïdes sont extrêmement rares, avec seulement quelques exemples parmi les vers plats, les crustacés, les amphibiens, les poissons et les lézards. Les animaux polyploïdes sont stériles parce que la méiose ne peut se dérouler normalement et produit plutôt des cellules-filles aneuploïdes qui ne peuvent pas produire de zygotes viables. Dans de rares cas, les animaux polyploïdes peuvent se reproduire de manière asexuée par haplodiploïdie, au cours de laquelle un œuf non fécondé se divise mitotiquement pour produire une progéniture. En revanche, la polyploïdie est très courante dans le règne végétal, et les plantes polyploïdes ont tendance à être plus grandes et plus robustes que les euploïdes de leur espèce (Figure\(\PageIndex{4}\)).

Non-disjonction des chromosomes sexuels chez l'homme
Les humains présentent des effets délétères spectaculaires avec les trisomies et les monosomies autosomiques. Par conséquent, il peut sembler contre-intuitif que les femmes et les hommes puissent fonctionner normalement, même s'ils portent des nombres différents du chromosome X. Plutôt qu'un gain ou une perte d'autosomes, les variations du nombre de chromosomes sexuels sont associées à des effets relativement légers. Cela est dû en partie à un processus moléculaire appelé inactivation de l'X. Au début du développement, lorsque les embryons de mammifères femelles ne contiennent que quelques milliers de cellules (contre des billions chez le nouveau-né), un chromosome X de chaque cellule s'inactive en se condensant étroitement pour former une structure quiescente (dormante) appelée corps de Barr. La probabilité qu'un chromosome X (d'origine maternelle ou paternelle) soit inactivé dans chaque cellule est aléatoire, mais une fois l'inactivation survenue, toutes les cellules dérivées de ce chromosome X inactif ou corps de Barr apparaîtront. Par ce processus, les femelles compensent leur double dose génétique de chromosome X. Chez les chats dits « écailles de tortue », l'inactivation du X embryonnaire est observée sous forme de panachure de couleur (Figure\(\PageIndex{5}\)). Les femelles hétérozygotes pour un gène de couleur de pelage lié à l'X exprimeront l'une des deux couleurs de pelage différentes sur différentes régions de leur corps, correspondant au chromosome X inactivé dans la cellule progénitrice de la cellule embryonnaire de cette région.

Une personne porteuse d'un nombre anormal de chromosomes X inactivera tous les chromosomes X sauf un dans chacune de ses cellules. Cependant, même les chromosomes X inactivés continuent d'exprimer quelques gènes, et les chromosomes X doivent se réactiver pour que les ovaires féminins puissent mûrir correctement. Par conséquent, les anomalies du chromosome X sont généralement associées à de légers défauts mentaux et physiques, ainsi qu'à la stérilité. Si le chromosome X est totalement absent, l'individu ne se développera pas in utero.
Plusieurs erreurs dans le nombre de chromosomes sexuels ont été caractérisées. Les individus dotés de trois chromosomes X, appelés Triplo-X, sont phénotypiquement des femmes mais présentent des retards de développement et une fertilité réduite. Le génotype XXY, qui correspond à un type de syndrome de Klinefelter, correspond à des individus phénotypiquement masculins présentant de petits testicules, des seins élargis et une pilosité réduite. Il existe des types plus complexes de syndrome de Klinefelter dans lesquels l'individu possède jusqu'à cinq chromosomes X. Dans tous les types, tous les chromosomes X, sauf un, sont inactivés pour compenser la dose génétique excessive. Cela se traduit par la présence de plusieurs corps de Barr dans chaque noyau cellulaire. Le syndrome de Turner, caractérisé par un génotype X0 (c'est-à-dire un seul chromosome sexuel), correspond à une personne phénotypiquement féminine présentant une petite taille, une peau palmée dans la région du cou, des troubles auditifs et cardiaques et une stérilité.
Duplications et suppressions
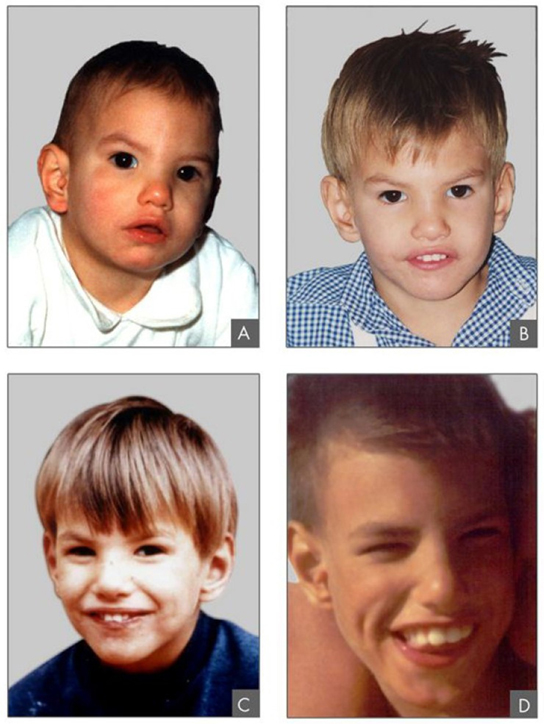
Outre la perte ou le gain d'un chromosome entier, un segment chromosomique peut être dupliqué ou perdu. Les duplications et les délétions produisent souvent des descendants qui survivent mais présentent des anomalies physiques et mentales. Les segments chromosomiques dupliqués peuvent fusionner avec des chromosomes existants ou peuvent être libres dans le noyau. Le cri-du-Chat (du français pour « cri du chat ») est un syndrome associé à des anomalies du système nerveux et à des caractéristiques physiques identifiables qui résultent de la délétion de la majeure partie du 5p (le petit bras du chromosome 5) (Figure\(\PageIndex{6}\)). Les nourrissons porteurs de ce génotype émettent un cri aigu caractéristique sur lequel le nom de la maladie est basé.
Réarrangements structuraux chromosomiques
Les cytologues ont caractérisé de nombreux réarrangements structuraux dans les chromosomes, mais les inversions et les translocations chromosomiques sont les plus courantes. Les deux sont identifiés au cours de la méiose par l'appariement adaptatif de chromosomes réarrangés avec leurs anciens homologues afin de maintenir un alignement génique approprié. Si les gènes portés par deux homologues ne sont pas orientés correctement, un événement de recombinaison peut entraîner la perte de gènes d'un chromosome et le gain de gènes sur l'autre. Cela produirait des gamètes aneuploïdes.
Inversions chromosomiques
Une inversion chromosomique est le détachement, la rotation à 180° et la réinsertion d'une partie d'un chromosome. Des inversions peuvent survenir dans la nature à la suite d'un cisaillement mécanique ou de l'action d'éléments transposables (séquences d'ADN spéciales capables de faciliter le réarrangement des segments chromosomiques à l'aide d'enzymes qui coupent et collent les séquences d'ADN). À moins qu'elles ne perturbent une séquence génétique, les inversions ne font que modifier l'orientation des gènes et sont susceptibles d'avoir des effets plus légers que les erreurs aneuploïdes. Cependant, une modification de l'orientation des gènes peut entraîner des changements fonctionnels, car les régulateurs de l'expression génique peuvent être déplacés hors de leur position par rapport à leurs cibles, ce qui entraîne des niveaux aberrants de produits géniques.
Une inversion peut être péricentrique et inclure le centromère, ou paracentrique et se produire à l'extérieur du centromère (). Une inversion péricentrique asymétrique par rapport au centromère peut modifier la longueur relative des bras chromosomiques, rendant ces inversions facilement identifiables.
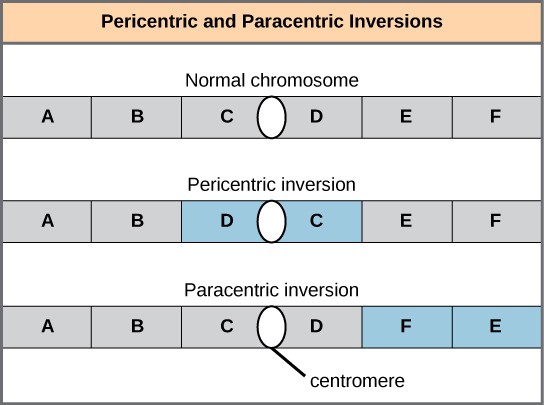
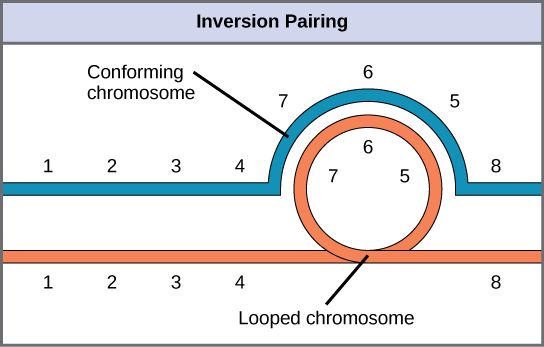
Lorsqu'un chromosome homologue subit une inversion mais pas l'autre, l'individu est décrit comme un hétérozygote d'inversion. Pour maintenir la synapse point par point pendant la méiose, un homologue doit former une boucle et l'autre homologue doit se mouler autour de celle-ci. Bien que cette topologie puisse garantir que les gènes sont correctement alignés, elle oblige également les homologues à s'étirer et peut être associée à des régions de synapses imprécises (Figure\(\PageIndex{8}\)).
Evolution Connection : l'inversion du chromosome 18
Les réarrangements structuraux des chromosomes ne produisent pas tous des individus non viables, altérés ou stériles. Dans de rares cas, un tel changement peut entraîner l'évolution d'une nouvelle espèce. En fait, une inversion péricentrique du chromosome 18 semble avoir contribué à l'évolution des humains. Cette inversion n'est pas présente chez nos plus proches parents génétiques, les chimpanzés. Les humains et les chimpanzés diffèrent cytogénétiquement par des inversions péricentriques sur plusieurs chromosomes et par la fusion de deux chromosomes distincts chez les chimpanzés qui correspondent au chromosome 2 chez l'homme.
On pense que l'inversion péricentrique du chromosome 18 s'est produite chez les premiers humains à la suite de leur divergence par rapport à un ancêtre commun aux chimpanzés il y a environ cinq millions d'années. Les chercheurs qui ont caractérisé cette inversion ont suggéré qu'environ 19 000 bases nucléotidiques ont été dupliquées sur le 18p, et que la région dupliquée s'est inversée et réinsérée sur le chromosome 18 d'un être humain ancestral.
Une comparaison des gènes humains et des chimpanzés dans la région de cette inversion indique que deux gènes, ROCK1 et USP14, qui sont adjacents au chromosome 17 du chimpanzé (qui correspond au chromosome 18 humain) sont positionnés plus loin sur le chromosome 18 humain. Cela suggère que l'un des points critiques d'inversion s'est produit entre ces deux gènes. Il est intéressant de noter que les humains et les chimpanzés expriment l'USP14 à des niveaux distincts dans des types de cellules spécifiques, notamment les cellules corticales et les fibroblastes. Peut-être que l'inversion du chromosome 18 chez un être humain ancestral a repositionné des gènes spécifiques et réinitialisé leurs niveaux d'expression d'une manière utile. Comme ROCK1 et USP14 codent pour des enzymes cellulaires, une modification de leur expression pourrait altérer la fonction cellulaire. On ne sait pas comment cette inversion a contribué à l'évolution des hominidés, mais elle semble être un facteur significatif de la divergence entre les humains et les autres primates. 1
Translocations
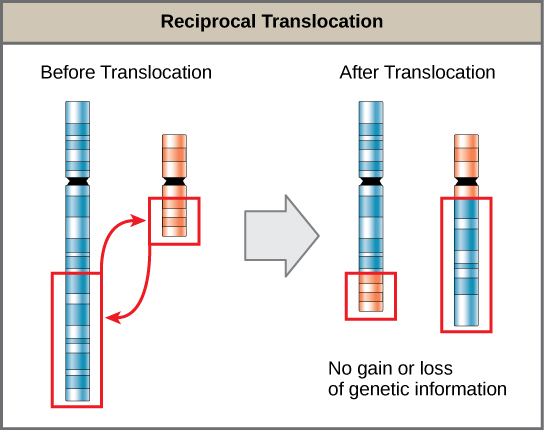
Une translocation se produit lorsqu'un segment d'un chromosome se dissocie et se rattache à un chromosome différent et non homologue. Les translocations peuvent être bénignes ou avoir des effets dévastateurs selon la façon dont la position des gènes est modifiée par rapport aux séquences régulatrices. Des translocations spécifiques ont notamment été associées à plusieurs cancers et à la schizophrénie. Les translocations réciproques résultent de l'échange de segments chromosomiques entre deux chromosomes non homologues, de telle sorte qu'il n'y a ni gain ni perte d'informations génétiques (Figure\(\PageIndex{9}\)).
Résumé
Le nombre, la taille, la forme et le schéma de bandes des chromosomes les rendent facilement identifiables sur un caryogramme et permettent d'évaluer de nombreuses anomalies chromosomiques. Les troubles du nombre chromosomique, ou aneuploïdies, sont généralement mortels pour l'embryon, bien que quelques génotypes trisomiques soient viables. En raison de l'inactivation du chromosome X, les aberrations des chromosomes sexuels ont généralement des effets phénotypiques plus légers. Les aneuploïdies incluent également les cas où des segments d'un chromosome sont dupliqués ou supprimés. Les structures chromosomiques peuvent également être réarrangées, par exemple par inversion ou translocation. Ces deux aberrations peuvent entraîner des effets phénotypiques problématiques. Comme elles forcent les chromosomes à adopter des topologies non naturelles au cours de la méiose, les inversions et les translocations sont souvent associées à une baisse de la fertilité en raison de la probabilité de non-disjonction.
Connexions artistiques
Figure\(\PageIndex{2}\) : Laquelle des affirmations suivantes concernant la non-disjonction est vraie ?
- La non-disjonction ne produit que des gamètes avec des chromosomes n+1 ou n—1.
- L'absence de disjonction pendant la méiose II entraîne la production de 50 % de gamètes normaux.
- L'absence de disjonction au cours de la méiose I entraîne la production de 50 % de gamètes normaux.
- La non-disjonction donne toujours naissance à quatre types de gamètes différents.
- Réponse
-
B.
Notes
- 1 Violaine Goidts et al., « Duplication segmentaire associée à l'inversion spécifique à l'homme du chromosome 18 : un autre exemple de l'impact des duplications segmentaires sur l'évolution du caryotype et du génome chez les primates », Human Genetics. 115 (2004) :116-122
Lexique
- aneuploïde
- personne présentant une erreur dans le nombre de chromosomes ; y compris les délétions et les duplications de segments chromosomiques
- autosome
- n'importe lequel des chromosomes non sexuels
- inversion chromosomique
- détachement, rotation à 180° et réinsertion d'un bras chromosomique
- euploïde
- individu possédant le nombre de chromosomes approprié à son espèce
- caryogramme
- image photographique d'un caryotype
- caryotype
- nombre et apparence des chromosomes d'un individu ; y compris la taille, les motifs de bandes et la position du centromère
- monosomie
- génotype par ailleurs diploïde dans lequel un chromosome est absent
- non-disjonction
- incapacité des homologues synapsés à se séparer complètement et à migrer vers des pôles séparés lors de la première division cellulaire de la méiose
- paracentrique
- inversion qui se produit à l'extérieur du centromère
- péricentrique
- inversion impliquant le centromère
- polyploïde
- individu avec un nombre incorrect d'ensembles de chromosomes
- translocation
- processus par lequel un segment d'un chromosome se dissocie et se rattache à un chromosome différent non homologue
- trisomie
- génotype autrement diploïde dans lequel un chromosome entier est dupliqué
- Inactivation de X
- condensation des chromosomes X dans les corps de Barr au cours du développement embryonnaire chez les femelles pour compenser la double dose génétique