
12.3 : Lois relatives à l'héritage
- Page ID
- 189692

Compétences à développer
- Expliquer la loi de ségrégation et l'assortiment indépendant de Mendel en termes de génétique et d'événements de méiose
- Utilisez la méthode bifurquée et les règles de probabilité pour calculer la probabilité de génotypes et de phénotypes issus de multiples croisements de gènes
- Expliquer l'effet de la liaison et de la recombinaison sur les génotypes des gamètes
- Expliquer les résultats phénotypiques des effets épistatiques entre gènes
Mendel a généralisé les résultats de ses expériences sur les plants de pois en quatre postulats, dont certains sont parfois appelés « lois », qui décrivent la base de l'hérédité dominante et récessive chez les organismes diploïdes. Comme vous l'avez appris, il existe des extensions plus complexes du mendélisme qui ne présentent pas les mêmes rapports phénotypiques F 2 (3:1). Néanmoins, ces lois résument les bases de la génétique classique.
Paires de facteurs unitaires, ou gènes
Mendel a d'abord suggéré que les facteurs unitaires appariés de l'hérédité étaient transmis fidèlement de génération en génération par la dissociation et la réassociation de facteurs appariés au cours de la gamétogenèse et de la fécondation, respectivement. Après avoir croisé des pois aux traits contrastés et découvert que le caractère récessif avait refait surface chez la génération F 2, Mendel a déduit que les facteurs héréditaires devaient être hérités sous forme d'unités distinctes. Cette découverte contredisait la croyance de l'époque selon laquelle les traits parentaux étaient mélangés chez la progéniture.
Les allèles peuvent être dominants ou récessifs
La loi de dominance de Mendel stipule que chez un hétérozygote, un trait masquera la présence d'un autre trait pour la même caractéristique. Au lieu que les deux allèles contribuent à un phénotype, l'allèle dominant sera exprimé exclusivement. L'allèle récessif restera « latent » mais sera transmis à la progéniture de la même manière que l'allèle dominant. Le caractère récessif ne sera exprimé que par les descendants qui possèdent deux copies de cet allèle (Figure\(\PageIndex{1}\)), et ces descendants se reproduisent correctement lorsqu'ils se croisent eux-mêmes.
Depuis les expériences de Mendel sur des plants de pois, d'autres chercheurs ont découvert que la loi de la dominance n'est pas toujours vraie. Au contraire, on a découvert l'existence de plusieurs modèles d'héritage différents.

Ségrégation égale des allèles
Constatant que des plants de pois reproducteurs présentant des caractères contrastés donnaient naissance à des générations F 1 qui exprimaient toutes le caractère dominant et à des générations F 2 exprimant les caractères dominants et récessifs dans un rapport de 3:1, Mendel a proposé la loi de ségrégation. Cette loi stipule que les facteurs unitaires appariés (gènes) doivent se séparer également en gamètes, de sorte que la progéniture ait une probabilité égale d'hériter de l'un ou l'autre facteur. Pour la génération F 2 d'un croisement monohybride, les trois combinaisons possibles de génotypes suivantes pourraient résulter : homozygote dominant, hétérozygote ou homozygote récessif. Comme les hétérozygotes peuvent provenir de deux voies différentes (recevant un allèle dominant et un allèle récessif de l'un ou l'autre des parents) et parce que les hétérozygotes et les individus dominants homozygotes sont phénotypiquement identiques, la loi soutient le ratio phénotypique de 3:1 observé par Mendel. La ségrégation égale des allèles est la raison pour laquelle nous pouvons appliquer le carré de Punnett pour prédire avec précision la progéniture de parents dont les génotypes sont connus. La base physique de la loi de ségrégation de Mendel est la première division de la méiose, dans laquelle les chromosomes homologues avec leurs différentes versions de chaque gène sont séparés en noyaux filles. Le rôle de la ségrégation méiotique des chromosomes dans la reproduction sexuée n'a pas été compris par la communauté scientifique du vivant de Mendel.
Assortiment indépendant
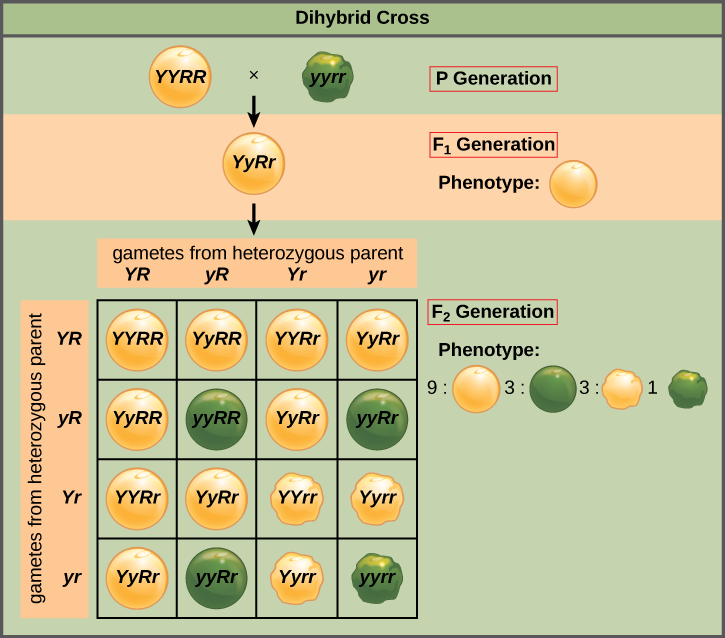
La loi de l'assortiment indépendant de Mendel stipule que les gènes ne s'influencent pas mutuellement en ce qui concerne le tri des allèles en gamètes, et que toutes les combinaisons possibles d'allèles pour chaque gène sont également susceptibles de se produire. L'assortiment indépendant de gènes peut être illustré par le croisement dihybride, un croisement entre deux parents reproducteurs authentiques qui expriment des traits différents pour deux caractéristiques. Tenez compte des caractéristiques de la couleur et de la texture des graines de deux plants de pois, l'un à graines vertes ridées (yyrr) et l'autre à graines jaunes et rondes (YYRR). Comme chaque parent est homozygote, la loi de ségrégation indique que les gamètes de la plante verte/ridée sont tous de l'an, et que les gamètes de la plante jaune/ronde sont tous YR. Par conséquent, les descendants de la génération F 1 sont tous des yYrR (Figure\(\PageIndex{2}\)).
Exercice
Chez les plants de pois, les fleurs violettes (P) dominent les fleurs blanches (p) et les pois jaunes (Y) dominent les pois verts (y). Quels sont les génotypes et phénotypes possibles pour un croisement entre des plants de pois pPyy et PpyY ? Combien de carrés avez-vous besoin pour effectuer une analyse au carré de Punnett de cette croix ?
- Réponse
-
Les génotypes possibles sont pPyy, pPyy, pPyy et pPyy. Les deux premiers génotypes donneraient lieu à des plantes à fleurs violettes et à pois jaunes, tandis que les deux derniers génotypes donneraient lieu à des plantes à fleurs blanches avec des pois jaunes, pour un ratio de 1:1 de chaque phénotype. Vous n'avez besoin que d'un carré de Punnett 2 × 2 (quatre carrés au total) pour effectuer cette analyse, car deux des allèles sont homozygotes.
Pour la génération F 2, la loi de ségrégation exige que chaque gamète reçoive un allèle R ou un allèle r en même temps qu'un allèle Y ou un allèle y. La loi de l'assortiment indépendant stipule qu'un gamète dans lequel un allèle r est trié serait également susceptible de contenir un allèle Y ou un allèle y. Ainsi, quatre gamètes tout aussi probables peuvent se former lorsque l'hétérozygote YyRR est auto-croisé, comme suit : YR, Yr, yR et yr. La disposition de ces gamètes en haut et à gauche d'un carré de Punnett 4 × 4 (Figure\(\PageIndex{2}\)) nous donne 16 combinaisons génotypiques tout aussi probables. À partir de ces génotypes, nous déduisons un ratio phénotypique de 9 rond/jaune/3 rond/vert/3 ridés/jaune/1 ridé/vert (Figure\(\PageIndex{2}\)). Ce sont les ratios de progéniture auxquels nous nous attendons, en supposant que nous ayons effectué les croisements avec un échantillon suffisamment important.
En raison de l'indépendance de l'assortiment et de la dominance, le rapport phénotypique dihybride 9:3:3:1 peut être réduit en deux rapports de 3:1, caractéristiques de tout croisement monohybride qui suit un schéma dominant et récessif. En négligeant la couleur des graines et en ne tenant compte que de la texture des graines du croisement dihybride ci-dessus, nous pouvons nous attendre à ce que les trois quarts des descendants de la génération F 2 soient ronds et qu'un quart soient froissés. De même, en isolant uniquement la couleur des graines, nous supposerions que les trois quarts des descendants F 2 seraient jaunes et un quart seraient verts. Le tri des allèles pour la texture et la couleur étant des événements indépendants, nous pouvons appliquer la règle du produit. Par conséquent, la proportion de descendants F 2 ronds et jaunes devrait être de (3/4) × (3/4) = 9/16, et la proportion de descendants ridés et verts devrait être de (1/4) × (1/4) = 1/16. Ces proportions sont identiques à celles obtenues à l'aide d'un carré de Punnett. La progéniture jaune ronde, verte et ridée peut également être calculée en utilisant la règle du produit, car chacun de ces génotypes comprend un phénotype dominant et un phénotype récessif. Par conséquent, la proportion de chacun est calculée comme suit : (3/4) × (1/4) = 3/16.
La loi de l'assortiment indépendant indique également qu'un croisement entre des parents jaunes froissés (YYRR) et des parents ronds verts (YYRR) produirait les mêmes descendants F 1 et F 2 que ceux du croisement YYRR x yyrr.
La base physique de la loi de l'assortiment indépendant réside également dans la méiose I, dans laquelle les différentes paires homologues s'alignent selon des orientations aléatoires. Chaque gamète peut contenir n'importe quelle combinaison de chromosomes paternel et maternel (et donc les gènes qui s'y trouvent) car l'orientation des tétrades sur le plan de la métaphase est aléatoire.
Méthode Forked-Line
Lorsque plus de deux gènes sont pris en compte, la méthode du carré de Punnett devient peu maniable. Par exemple, l'examen d'un croisement impliquant quatre gènes nécessiterait une grille 16 × 16 contenant 256 cases. Il serait extrêmement fastidieux de saisir manuellement chaque génotype. Pour les croisements plus complexes, les méthodes de la ligne bifurquée et des probabilités sont préférables.
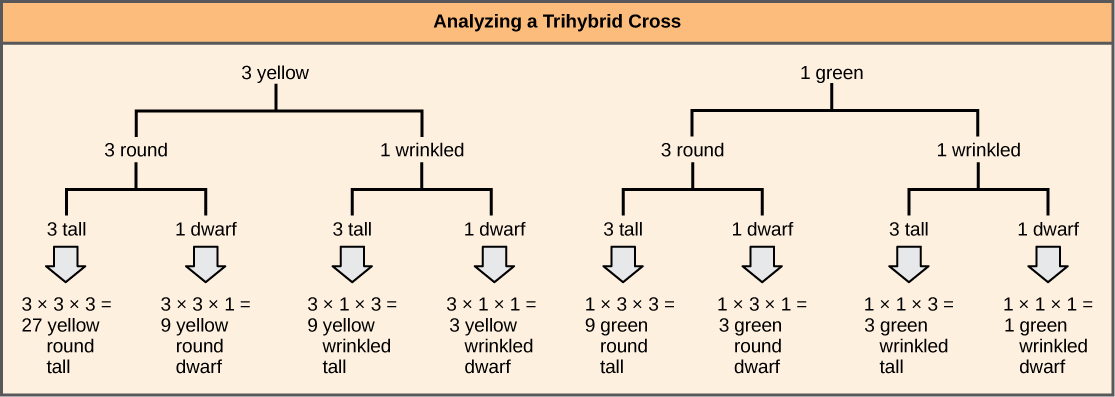
Pour préparer un diagramme bifurqué pour un croisement entre des hétérozygotes F 1 résultant d'un croisement entre les parents AABBCC et aabbcc, nous créons d'abord des rangées égales au nombre de gènes considérés, puis nous séparons les allèles de chaque rangée sur des lignes bifurquées conformément à probabilités de croisements monohybrides individuels (Figure\(\PageIndex{3}\)). Nous multiplions ensuite les valeurs le long de chaque chemin bifurqué pour obtenir les probabilités de progéniture F 2. Notez que ce processus est une version schématique de la règle du produit. Les valeurs le long de chaque voie bifurquée peuvent être multipliées car chaque gène s'assortit indépendamment. Pour un croisement trihybride, le rapport phénotypique F 2 est de 27:9:9:9:3:3:3:1.
Méthode des probabilités
Alors que la méthode de la ligne bifurquée est une approche schématique permettant de suivre les probabilités lors d'un croisement, la méthode des probabilités donne les proportions de descendants susceptibles de présenter chaque phénotype (ou génotype) sans assistance visuelle supplémentaire. Les deux méthodes utilisent la règle du produit et considèrent les allèles de chaque gène séparément. Plus tôt, nous avons examiné les proportions phénotypiques d'un croisement trihybride à l'aide de la méthode de la ligne bifurquée ; nous allons maintenant utiliser la méthode des probabilités pour examiner les proportions génotypiques d'un croisement avec encore plus de gènes.
Pour un croisement trihybride, l'écriture de la méthode de la ligne bifurquée est fastidieuse, mais pas aussi fastidieuse que l'utilisation de la méthode Punnett-square. Pour démontrer pleinement la puissance de la méthode des probabilités, nous pouvons toutefois envisager des calculs génétiques spécifiques. Par exemple, dans le cas d'un croisement tétrahybride entre des individus hétérozygotes pour les quatre gènes, et dans lequel les quatre gènes sont triés indépendamment et selon un schéma dominant et récessif, quelle proportion de la progéniture devrait être homozygote récessive pour les quatre allèles ? Plutôt que d'écrire tous les génotypes possibles, nous pouvons utiliser la méthode des probabilités. Nous savons que pour chaque gène, la fraction de descendance récessive homozygote sera de 1/4. En multipliant cette fraction pour chacun des quatre gènes, (1/4) × (1/4) × (1/4) × (1/4) × (1/4), nous déterminons que 1/256 de la progéniture sera récessive quadruplement homozygote.
Pour le même croisement tétrahybride, quelle est la proportion attendue de descendants présentant le phénotype dominant aux quatre loci ? Nous pouvons répondre à cette question en utilisant des proportions phénotypiques, mais faisons-le à la dure, en utilisant des proportions génotypiques. La question demande la proportion de descendants qui sont 1) homozygotes dominants en A ou hétérozygotes en A, et 2) homozygotes en B ou hétérozygotes en B, et ainsi de suite. Le fait de noter le « ou » et le « et » dans chaque cas indique clairement où appliquer les règles relatives à la somme et au produit. La probabilité d'une dominante homozygote en A est de 1/4 et la probabilité d'un hétérozygote en A est de 1/2. La probabilité de l'homozygote ou de l'hétérozygote est de 1/4 + 1/2 = 3/4 selon la règle de la somme. La même probabilité peut être obtenue de la même manière pour chacun des autres gènes, de sorte que la probabilité d'un phénotype dominant en A et B et en C et D est, selon la règle du produit, égale à 3/4 × 3/4 × 3/4 × 3/4 × 3/4, soit 27/64. Si vous ne savez pas comment combiner les probabilités, revenez à la méthode de la ligne bifurquée pour le clarifier.
Règles pour la fertilisation multihybride
Prédire les génotypes et les phénotypes des descendants issus de croisements donnés est la meilleure façon de tester vos connaissances en génétique mendélienne. Dans le cas d'un croisement multihybride qui obéit à un assortiment indépendant et suit un schéma dominant et récessif, plusieurs règles généralisées existent ; vous pouvez utiliser ces règles pour vérifier vos résultats lorsque vous effectuez des calculs génétiques (Tableau\(\PageIndex{1}\)). Pour appliquer ces règles, vous devez d'abord déterminer n, le nombre de paires de gènes hétérozygotes (le nombre de gènes séparant deux allèles chacun). Par exemple, un croisement entre des hétérozygotes aAbB et aAbB a un n de 2. En revanche, un croisement entre aAbB et aAbB a un n de 1 car A n'est pas hétérozygote.
| Règle générale | Nombre de paires de gènes hétérozygotes |
|---|---|
| Nombre de gamètes F 1 différents | 2 n |
| Nombre de génotypes F 2 différents | 3 n |
| Compte tenu de l'hérédité dominante et récessive, du nombre de phénotypes F 2 différents | 2 n |
Les gènes liés violent la loi de l'assortiment indépendant
Bien que toutes les caractéristiques du pois de Mendel se soient comportées selon la loi de l'assortiment indépendant, nous savons maintenant que certaines combinaisons d'allèles ne sont pas héritées indépendamment les unes des autres. Les gènes situés sur des chromosomes non homologues distincts seront toujours triés indépendamment. Cependant, chaque chromosome contient des centaines ou des milliers de gènes, organisés linéairement sur les chromosomes, comme des perles sur une chaîne. La ségrégation des allèles en gamètes peut être influencée par la liaison, selon laquelle les gènes situés physiquement à proximité les uns des autres sur le même chromosome sont plus susceptibles d'être hérités par paire. Cependant, en raison du processus de recombinaison, ou « croisement », il est possible que deux gènes d'un même chromosome se comportent indépendamment, ou comme s'ils n'étaient pas liés. Pour comprendre cela, examinons les bases biologiques de la liaison et de la recombinaison des gènes.
Les chromosomes homologues possèdent les mêmes gènes dans le même ordre linéaire. Les allèles peuvent différer selon les paires de chromosomes homologues, mais pas les gènes auxquels ils correspondent. En vue de la première division de la méiose, les chromosomes homologues se répliquent et se synapsent. Comme les gènes des homologues s'alignent les uns sur les autres. À ce stade, des segments de chromosomes homologues échangent des segments linéaires de matériel génétique (Figure\(\PageIndex{4}\)). Ce processus est appelé recombinaison, ou croisement, et il s'agit d'un processus génétique courant. Comme les gènes sont alignés lors de la recombinaison, l'ordre des gènes n'est pas modifié. La recombinaison a plutôt pour résultat que les allèles maternels et paternels sont combinés sur le même chromosome. Sur un chromosome donné, plusieurs événements de recombinaison peuvent se produire, provoquant un brassage important des allèles.
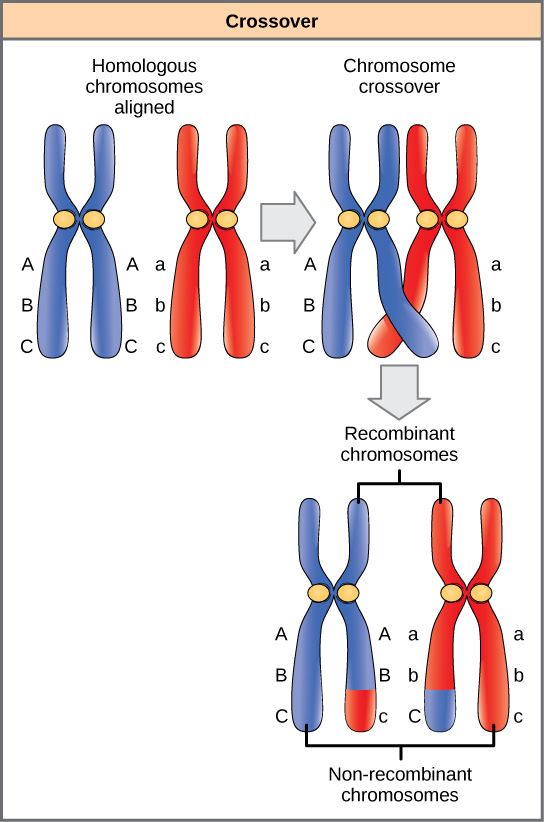
Lorsque deux gènes sont situés à proximité sur le même chromosome, ils sont considérés comme liés et leurs allèles ont tendance à être transmis conjointement par méiose. Pour illustrer cela, imaginez un croisement dihybride impliquant la couleur des fleurs et la hauteur de la plante dans lequel les gènes se trouvent côte à côte sur le chromosome. Si un chromosome homologue possède des allèles pour les plantes hautes et les fleurs rouges, et que l'autre chromosome possède des gènes pour les plantes courtes et les fleurs jaunes, alors, lorsque les gamètes se forment, les allèles grands et rouges se combinent pour former un gamète et les allèles courts et jaunes entrent dans d'autres gamètes. Ces génotypes sont appelés génotypes parentaux parce qu'ils ont été hérités intacts des parents de l'individu producteur de gamètes. Mais contrairement à ce qui se passerait si les gènes se trouvaient sur des chromosomes différents, il n'y aura pas de gamètes avec des allèles longs et jaunes et aucun gamète avec des allèles courts et rouges. Si vous créez le carré de Punnett avec ces gamètes, vous verrez que la prédiction mendélienne classique d'un résultat 9:3:3:1 d'un croisement dihybride ne s'applique pas. À mesure que la distance entre deux gènes augmente, la probabilité d'un ou de plusieurs croisements entre eux augmente et les gènes se comportent davantage comme s'ils se trouvaient sur des chromosomes distincts. Les généticiens ont utilisé la proportion de gamètes recombinants (ceux qui ne ressemblent pas aux parents) pour mesurer la distance entre les gènes d'un chromosome. À partir de ces informations, ils ont construit des cartes détaillées des gènes sur les chromosomes d'organismes bien étudiés, y compris les humains.
La publication phare de Mendel ne fait aucune mention du lien, et de nombreux chercheurs se sont demandé s'il avait rencontré un lien, mais ont choisi de ne pas publier ces croix par crainte qu'elles n'invalident son postulat d'assortiment indépendant. Le pois de jardin possède sept chromosomes, et certains ont suggéré que son choix de sept caractéristiques n'était pas le fruit du hasard. Cependant, même si les gènes qu'il a examinés n'étaient pas localisés sur des chromosomes distincts, il est possible qu'il n'ait tout simplement pas observé de liaison en raison des effets de brassage importants de la recombinaison.
Connexion entre les méthodes scientifiques : vérification de l'hypothèse d'un assortiment indépendant
Pour mieux apprécier la quantité de travail et d'ingéniosité nécessaires aux expériences de Mendel, passez par l'un des croisements dihybrides de Mendel.
Question : Quels seront les résultats d'un croisement dihybride ?
Contexte : Considérez que les plants de pois mûrissent en une seule saison de croissance et que vous avez accès à un grand jardin dans lequel vous pouvez cultiver des milliers de plants de pois. Il existe plusieurs plantes reproducteurs authentiques présentant les paires de caractères suivantes : des plantes hautes avec des gousses gonflées et des plantes naines avec des gousses resserrées. Avant que les plants soient arrivés à maturité, vous devez retirer les organes producteurs de pollen des plants hauts ou gonflés lors de vos croisements afin d'empêcher l'autofécondation. Lors de la maturation des plantes, les plantes sont croisées manuellement en transférant le pollen des plantes naines ou resserrées vers les stigmates des plantes hautes ou gonflées.
Hypothèse : Les deux paires de traits seront triées indépendamment selon les lois mendéliennes. Lorsque les parents reproducteurs sont croisés, tous les descendants F1 sont grands et ont des gousses gonflées, ce qui indique que les traits grands et gonflés dominent les traits nains et restreints, respectivement. Un autocroisement des hétérozygotes F 1 donne 2 000 descendants F 2.
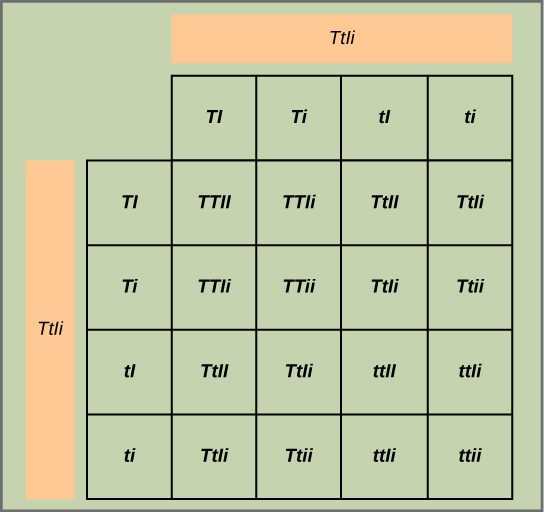
Testez l'hypothèse : étant donné que chaque paire de traits est triée indépendamment, les ratios taille:nain et gonflé:resserré devraient chacun être de 3:1. La paire de traits hauteur/nain est appelée t/t, et la paire de traits gonflés/resserrés est désignée I/i. Chaque membre de la génération F 1 possède donc un génotype du TTiI. Construisez une grille analogue à la Figure\(\PageIndex{2}\), dans laquelle vous croisez deux individus TTiI. Chaque individu peut donner quatre combinaisons de deux traits : TI, Ti, Ti ou ti, ce qui signifie qu'il existe 16 possibilités de génotypes de progéniture. Comme les allèles T et I sont dominants, toute personne possédant un ou deux de ces allèles exprimera les phénotypes hauts ou gonflés, respectivement, qu'elle possède également un allèle t ou i. Seuls les individus qui sont tt ou ii exprimeront respectivement les allèles nains et restreints. Comme le montre la figure\(\PageIndex{5}\), vous prédisez que vous observerez les proportions suivantes pour la progéniture : grand/gonflé : taille/resserré/nain/gonflé : nain/rétréci dans un rapport de 9:3:3:1. Remarquez sur la grille que lorsque vous considérez isolément les paires de traits hauteur/nain et gonflé/resserré, elles sont chacune héritées selon un ratio de 3:1.
Testez l'hypothèse : vous croisez les plantes naines et les plantes hautes, puis vous croisez vous-même la progéniture. Pour de meilleurs résultats, cette opération est répétée avec des centaines, voire des milliers de plants de pois. Quelles précautions particulières doivent être prises lors des croisements et lors de la culture des plantes ?
Analysez vos données : Vous observez les phénotypes de plantes suivants dans la génération F 2 : 2 706 hautes/gonflées, 930 hautes/resserrées, 888 naines/gonflées et 300 naines/resserrées. Réduisez ces résultats à un ratio et déterminez s'ils sont conformes aux lois mendéliennes.
Formez une conclusion : les résultats étaient-ils proches du ratio phénotypique de 9:3:3:1 attendu ? Les résultats confirment-ils la prédiction ? Que pourrait-on observer si l'on utilisait beaucoup moins de plantes, étant donné que les allèles se séparent aléatoirement en gamètes ? Imaginez que vous cultivez autant de plants de pois et considérez la possibilité d'erreurs expérimentales. Par exemple, que se passerait-il s'il y avait beaucoup de vent un jour ?
Épistase
Les études de Mendel sur les plants de pois impliquaient que la somme du phénotype d'un individu était contrôlée par des gènes (ou, comme il les appelait, des facteurs unitaires), de sorte que chaque caractéristique était contrôlée de manière distincte et complète par un seul gène. En fait, les caractéristiques observables uniques sont presque toujours sous l'influence de plusieurs gènes (chacun comportant deux allèles ou plus) agissant à l'unisson. Par exemple, au moins huit gènes contribuent à la couleur des yeux chez l'homme.
Dans certains cas, plusieurs gènes peuvent contribuer à certains aspects d'un phénotype commun sans que leurs produits géniques n'interagissent directement. Dans le cas du développement d'organes, par exemple, les gènes peuvent être exprimés de manière séquentielle, chaque gène augmentant la complexité et la spécificité de l'organe. Les gènes peuvent fonctionner de manière complémentaire ou synergique, de sorte que deux gènes ou plus doivent être exprimés simultanément pour affecter un phénotype. Les gènes peuvent également s'opposer les uns aux autres, un gène modifiant l'expression d'un autre.
Dans l'épistase, l'interaction entre les gènes est antagoniste, de sorte qu'un gène masque ou interfère avec l'expression d'un autre. « Épistase » est un mot composé de racines grecques qui signifie « debout ». Les allèles masqués ou réduits au silence sont dits hypostatiques par rapport aux allèles épistatiques responsables du masquage. Souvent, la base biochimique de l'épistase est une voie génétique dans laquelle l'expression d'un gène dépend de la fonction d'un gène qui le précède ou le suit dans la voie.
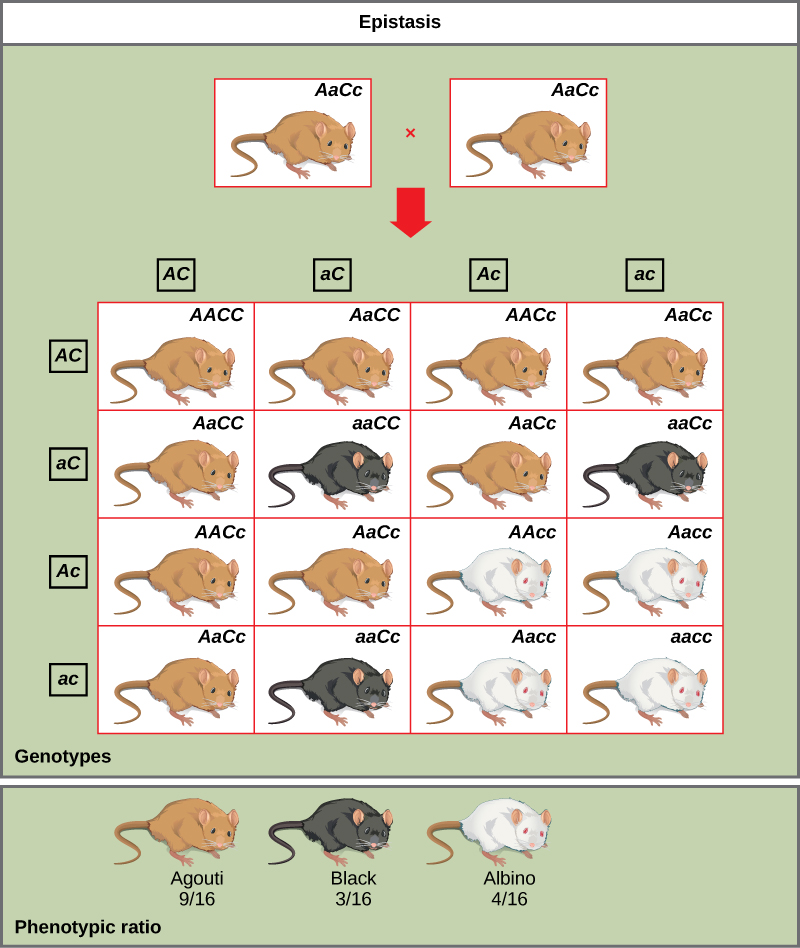
La pigmentation chez la souris est un exemple d'épistase. La couleur du pelage sauvage, agouti (AA), domine la fourrure unie (aa). Cependant, un gène distinct (C) est nécessaire à la production de pigments. Une souris présentant un allèle c récessif à ce locus ne peut pas produire de pigment et est albinos, quel que soit l'allèle présent au locus A (Figure\(\PageIndex{6}\)). Par conséquent, les génotypes Aacc, Aacc et aacc produisent tous le même phénotype albinos. Un croisement entre des hétérozygotes pour les deux gènes (aAcc x aAcc) produirait une progéniture avec un ratio phénotypique de 9 agouti:3 couleur solide:4 albinos (Figure\(\PageIndex{6}\)). Dans ce cas, le gène C est épistatique par rapport au gène A.
L'épistase peut également se produire lorsqu'un allèle dominant masque l'expression d'un gène distinct. La couleur des fruits des courges d'été s'exprime de cette manière. L'expression récessive homozygote du gène W (ww) associée à l'expression homozygote dominante ou hétérozygote du gène Y (YY ou Yy) génère des fruits jaunes, et le génotype wwyy produit des fruits verts. Toutefois, si une copie dominante du gène W est présente sous forme homozygote ou hétérozygote, la courge d'été produira des fruits blancs quels que soient les allèles Y. Un croisement entre des hétérozygotes blancs des deux gènes (WWyy × Wwyy) produirait une progéniture avec un ratio phénotypique de 12 blanc:3 jaune/1 vert.
Enfin, l'épistase peut être réciproque de telle sorte que l'un ou l'autre gène, lorsqu'il est présent sous sa forme dominante (ou récessive), exprime le même phénotype. Chez la capsella bursa-pastoris (capsella bursa-pastoris), la caractéristique de la forme des graines est contrôlée par deux gènes qui entretiennent une relation épistatique dominante. Lorsque les gènes A et B sont tous deux homozygotes récessifs (aabb), les graines sont ovoïdes. Si l'allèle dominant de l'un ou l'autre de ces gènes est présent, on obtient des graines triangulaires. C'est-à-dire que tous les génotypes possibles autres que l'aabb produisent des graines triangulaires, et un croisement entre des hétérozygotes pour les deux gènes (aAbB x aAbB) produirait une progéniture avec un ratio phénotypique de 15 triangulaires/1 ovoïde.
Lorsque vous étudiez des problèmes génétiques, gardez à l'esprit que toute caractéristique qui aboutit à un ratio phénotypique de 16 est typique d'une interaction entre deux gènes. Rappelons le modèle d'hérédité phénotypique du croisement dihybride de Mendel, qui considérait deux gènes sans interaction : 9:3:3:1. De même, nous pouvons nous attendre à ce que les paires de gènes en interaction présentent également des ratios exprimés en 16 parties. Notez que nous supposons que les gènes qui interagissent ne sont pas liés ; ils continuent de s'agencer indépendamment en gamètes.
Résumé
Mendel a postulé que les gènes (caractéristiques) sont hérités sous forme de paires d'allèles (traits) qui se comportent selon un schéma dominant et récessif. Les allèles se séparent en gamètes de telle sorte que chaque gamète est également susceptible de recevoir l'un ou l'autre des deux allèles présents chez un individu diploïde. De plus, les gènes sont répartis en gamètes indépendamment les uns des autres. C'est-à-dire que les allèles ne sont généralement pas plus susceptibles de se séparer en un gamète avec un allèle particulier d'un autre gène. Un croisement dihybride présente un assortiment indépendant lorsque les gènes en question se trouvent sur des chromosomes différents ou éloignés les uns des autres sur le même chromosome. Pour les croisements impliquant plus de deux gènes, utilisez la ligne bifurquée ou des méthodes de probabilité pour prédire les génotypes et les phénotypes de la progéniture plutôt que le carré de Punnett.
Bien que les chromosomes se trient indépendamment en gamètes au cours de la méiose, la loi d'assortiment indépendant de Mendel fait référence aux gènes et non aux chromosomes, et un seul chromosome peut porter plus de 1 000 gènes. Lorsque les gènes sont situés à proximité sur le même chromosome, leurs allèles ont tendance à être hérités ensemble. Il en résulte des ratios de progéniture qui violent la loi de Mendel sur l'assortiment indépendant. Cependant, la recombinaison permet d'échanger du matériel génétique sur des chromosomes homologues de telle sorte que les allèles maternels et paternels peuvent être recombinés sur le même chromosome. C'est pourquoi les allèles d'un chromosome donné ne sont pas toujours hérités ensemble. La recombinaison est un événement aléatoire qui se produit n'importe où sur un chromosome. Par conséquent, les gènes très éloignés sur le même chromosome sont susceptibles de continuer à s'agencer indépendamment en raison des événements de recombinaison qui se sont produits dans l'espace chromosomique intermédiaire.
Qu'ils soient triés indépendamment ou non, les gènes peuvent interagir au niveau des produits géniques de telle sorte que l'expression d'un allèle pour un gène masque ou modifie l'expression d'un allèle pour un gène différent. C'est ce qu'on appelle l'épistase.
Lexique
- dihybride
- résultat d'un croisement entre deux parents reproducteurs authentiques qui expriment des traits différents pour deux caractéristiques
- épistase
- interaction antagoniste entre gènes de telle sorte qu'un gène masque ou interfère avec l'expression d'un autre
- loi de domination
- chez un hétérozygote, un trait masquera la présence d'un autre trait pour la même caractéristique
- loi de l'assortiment indépendant
- les gènes ne s'influencent pas mutuellement en ce qui concerne le tri des allèles en gamètes ; toutes les combinaisons possibles d'allèles sont également susceptibles de se produire
- loi de ségrégation
- les facteurs unitaires appariés (c'est-à-dire les gènes) se séparent également en gamètes, de sorte que la progéniture a la même probabilité d'hériter d'une combinaison de facteurs
- lien
- phénomène dans lequel les allèles situés à proximité les uns des autres sur le même chromosome sont plus susceptibles d'être hérités ensemble