
12.1 : Les expériences de Mendel et les lois des probabilités
- Page ID
- 189710

Compétences à développer
- Décrire les raisons scientifiques du succès des travaux expérimentaux de Mendel
- Décrire les résultats attendus des croisements monohybrides impliquant des allèles dominants et récessifs
- Appliquez les règles de somme et de produit pour calculer les probabilités
Johann Gregor Mendel (1822-1884) (Figure\(\PageIndex{1}\)) a toujours été un apprenant, un enseignant, un scientifique et un homme de foi. Jeune adulte, il a rejoint l'abbaye augustinienne de Saint-Thomas à Brno, dans l'actuelle République tchèque. Soutenu par le monastère, il a enseigné la physique, la botanique et les sciences naturelles aux niveaux secondaire et universitaire. En 1856, il a entamé une recherche de dix ans sur les modèles d'héritage des abeilles domestiques et des plantes, avant de choisir les plants de pois comme principal système modèle (un système doté de caractéristiques pratiques utilisé pour étudier un phénomène biologique spécifique à appliquer à d'autres systèmes). En 1865, Mendel a présenté les résultats de ses expériences sur près de 30 000 plants de pois à la Société d'histoire naturelle locale. Il a démontré que les traits sont transmis fidèlement des parents à la progéniture indépendamment des autres traits et selon des modèles dominants et récessifs. En 1866, il a publié son travail, Experiments in Plant Hybridization, 1 dans les actes de la Société d'histoire naturelle de Brünn.

Les travaux de Mendel sont passés pratiquement inaperçus auprès de la communauté scientifique qui croyait, à tort, que le processus d'héritage impliquait un mélange de traits parentaux qui produisait une apparence physique intermédiaire chez la progéniture ; ce processus hypothétique semblait correct en raison de ce que nous connaissons aujourd'hui sous le nom de variation continue. La variation continue résulte de l'action de nombreux gènes pour déterminer une caractéristique telle que la taille humaine. Les enfants semblent être un « mélange » des traits de leurs parents lorsque nous examinons des caractéristiques qui présentent des variations continues. La théorie de l'hérédité mixte affirmait que les traits parentaux d'origine avaient été perdus ou absorbés par le mélange chez la progéniture, mais nous savons maintenant que ce n'est pas le cas. Mendel a été le premier chercheur à l'observer. Au lieu d'utiliser des caractéristiques continues, Mendel a travaillé avec des caractères hérités dans des classes distinctes (en particulier, les fleurs violettes par rapport aux fleurs blanches) ; c'est ce que l'on appelle la variation discontinue. Le choix de ces types de traits par Mendel lui a permis de constater expérimentalement que les traits n'étaient pas mélangés chez la progéniture et qu'ils n'étaient pas absorbés, mais qu'ils conservaient leur caractère distinct et pouvaient être transmis. En 1868, Mendel est devenu abbé du monastère et a échangé ses activités scientifiques contre ses fonctions pastorales. Il n'a pas été reconnu pour ses contributions scientifiques extraordinaires de son vivant. En fait, ce n'est qu'en 1900 que ses travaux ont été redécouverts, reproduits et revitalisés par des scientifiques sur le point de découvrir les bases chromosomiques de l'hérédité.
Système de modèles de Mendel
Le travail fondateur de Mendel a été réalisé en utilisant le pois de jardin, Pisum sativum, pour étudier l'héritage. Cette espèce s'autoféconde naturellement, de sorte que le pollen rencontre des ovules dans les fleurs individuelles. Les pétales de fleurs restent hermétiquement fermés jusqu'à la pollinisation, empêchant ainsi la pollinisation par d'autres plantes. Il en résulte des plants de pois hautement consanguins, ou « véritables reproducteurs ». Ce sont des plantes qui produisent toujours une progéniture qui ressemble au parent. En expérimentant des plants de pois reproducteurs authentiques, Mendel a évité l'apparition de caractères inattendus chez la progéniture qui pourraient apparaître si les plantes n'étaient pas réellement reproductrices. Le pois de jardin atteint également sa maturité en une saison, ce qui signifie que plusieurs générations peuvent être évaluées sur une période relativement courte. Enfin, de grandes quantités de pois de jardin ont pu être cultivées simultanément, ce qui a permis à Mendel de conclure que ses résultats n'étaient pas le fruit du hasard.
Croix mendéliennes
Mendel a réalisé des hybridations, qui impliquent l'accouplement de deux individus reproducteurs réels qui ont des traits différents. Dans le cas du pois, qui est naturellement autogame, cela se fait en transférant manuellement le pollen de l'anthère d'un plant de pois mature d'une variété vers le stigmate d'un plant de pois mature distinct de la seconde variété. Chez les plantes, le pollen transporte les gamètes mâles (spermatozoïdes) vers le stigmate, un organe collant qui piège le pollen et permet au sperme de se déplacer le long du pistil jusqu'aux gamètes femelles (ovules) situés en dessous. Pour empêcher le plant de pois qui recevait du pollen de s'autoféconder et de fausser ses résultats, Mendel a minutieusement retiré toutes les anthères des fleurs de la plante avant qu'elles n'aient eu la chance de mûrir.
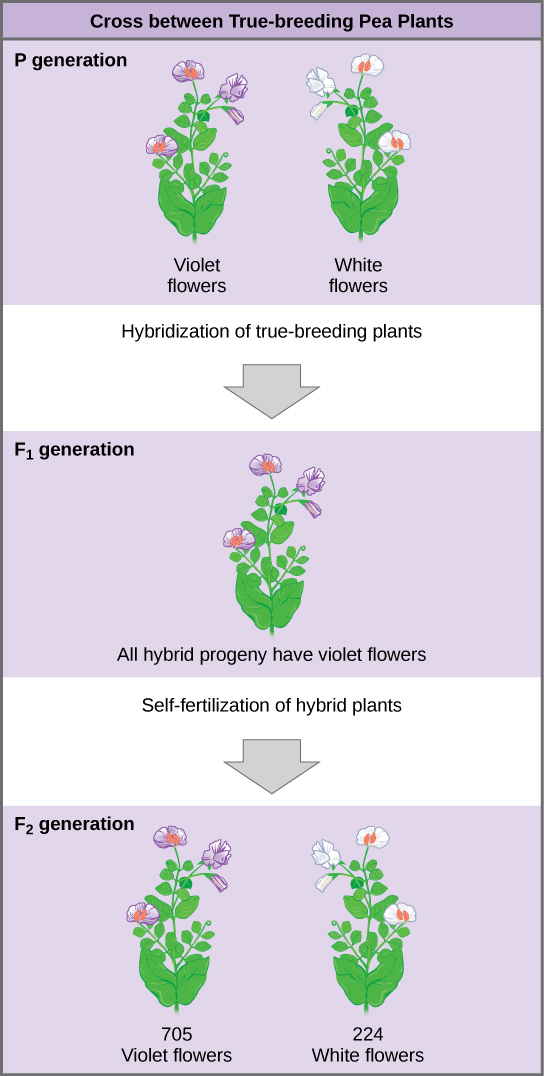
Les plantes utilisées dans les croisements de première génération étaient appelées plantes P 0, ou plantes parentales de première génération (Figure\(\PageIndex{2}\)). Mendel a collecté les graines appartenant aux plantes P 0 issues de chaque croisement et les a cultivées la saison suivante. Ces descendants étaient appelés la génération F 1, ou la première génération filiale (filiale = progéniture, fille ou fils). Une fois que Mendel a examiné les caractéristiques des plantes de la génération F1, il les a autorisées à s'autofertiliser naturellement. Il a ensuite collecté et cultivé les graines des plantes F1 pour produire la génération F 2, ou deuxième génération filiale. Les expériences de Mendel se sont étendues au-delà de la génération F 2 aux générations F 3 et F 4, etc., mais c'est le rapport des caractéristiques des générations P 0 −F 1 −F 2 qui a été le plus intrigant et est devenu la base de Les postulats de Mendel.
Les caractéristiques du pois de jardin ont révélé les bases de l'hérédité
Dans sa publication de 1865, Mendel a rapporté les résultats de ses croisements impliquant sept caractéristiques différentes, chacune présentant deux traits contrastés. Un trait est défini comme une variation de l'apparence physique d'une caractéristique héréditaire. Les caractéristiques comprenaient la hauteur de la plante, la texture des graines, la couleur des graines, la couleur de la fleur, la taille des gousses de pois, la couleur des gousses de pois et En ce qui concerne la caractéristique de la couleur des fleurs, par exemple, les deux traits contrastés étaient le blanc et le violet. Pour examiner pleinement chaque caractéristique, Mendel a généré un grand nombre de plantes F 1 et F 2, communiquant les résultats de 19 959 plantes F 2 uniquement. Ses conclusions étaient cohérentes.
Quels résultats Mendel a-t-il trouvés dans ses croix pour la couleur des fleurs ? Mendel a d'abord confirmé qu'il possédait des plantes dont la couleur des fleurs était blanche ou violette. Indépendamment du nombre de générations examinées par Mendel, tous les descendants auto-croisés de parents à fleurs blanches avaient des fleurs blanches, et tous les descendants auto-croisés de parents à fleurs violettes avaient des fleurs violettes. De plus, Mendel a confirmé que, hormis la couleur des fleurs, les plants de pois étaient physiquement identiques.
Une fois ces validations terminées, Mendel a appliqué le pollen d'une plante à fleurs violettes sur le stigmate d'une plante à fleurs blanches. Après avoir cueilli et semé les graines issues de ce croisement, Mendel a découvert que 100 pour cent des hybrides de la génération F1 avaient des fleurs violettes. La sagesse populaire de l'époque aurait prédit que les fleurs hybrides seraient violettes pâles ou que les plantes hybrides auraient un nombre égal de fleurs blanches et violettes. En d'autres termes, on s'attendait à ce que les traits parentaux contrastés se fondent dans la progéniture. Les résultats de Mendel ont plutôt démontré que le caractère de la fleur blanche de la génération F1 avait complètement disparu.
Fait important, Mendel n'y a pas arrêté ses expérimentations. Il a laissé les plantes F 1 s'autofertiliser et a découvert que, sur les plantes de la génération F 2, 705 avaient des fleurs violettes et 224 des fleurs blanches. Il s'agissait d'un ratio de 3,15 fleurs violettes pour une fleur blanche, soit environ 3:1. Lorsque Mendel a transféré le pollen d'une plante à fleurs violettes au stigmate d'une plante à fleurs blanches et vice versa, il a obtenu à peu près le même ratio quel que soit le parent, mâle ou femelle, qui a contribué à quel caractère. C'est ce que l'on appelle une croix réciproque, une croix appariée dans laquelle les traits respectifs du mâle et de la femelle d'une croix deviennent les traits respectifs de la femme et de l'homme de l'autre croix. Pour les six autres caractéristiques étudiées par Mendel, les générations F 1 et F 2 se sont comportées de la même manière que pour la couleur des fleurs. L'un des deux caractères disparaîtrait complètement de la génération F 1 pour réapparaître dans la génération F 2 dans un rapport d'environ 3:1 (tableau\(\PageIndex{1}\)).
| Caractéristique | Traits contrastés du P 0 | F 1 Caractéristiques de la progéniture | F 2 Caractéristiques de la progéniture | Ratios de traits F 2 |
|---|---|---|---|---|
| Couleur de la fleur | Violet contre blanc | 100 pour cent violet |
|
3. 15:1 |
| Position de la fleur | Axial ou terminal | 100 % axial |
|
3. 14:1 |
| Hauteur de la plante | Grand contre nain | 100 pour cent de haut |
|
2. 84:1 |
| Texture des graines | Rond ou froissé | 100 pour cent rond |
|
2. 96:1 |
| Couleur de la graine | Jaune contre vert | 100 % jaune |
|
3. 01:1 |
| Texture des gousses | Gonflé ou resserré | 100 % gonflé |
|
2. 95:1 |
| Couleur de la gousse | Vert contre jaune | 100 % vert |
|
2. 82:1 |
Après avoir compilé ses résultats pour plusieurs milliers de plantes, Mendel a conclu que les caractéristiques pouvaient être divisées en traits exprimés et en traits latents. Il les a appelés, respectivement, traits dominants et récessifs. Les traits dominants sont ceux qui sont hérités tels quels lors d'une hybridation. Les traits récessifs deviennent latents ou disparaissent chez les descendants d'une hybridation. Le caractère récessif réapparaît cependant chez la descendance de la progéniture hybride. Un exemple de trait dominant est le trait de fleur violette. Pour cette même caractéristique (couleur des fleurs), les fleurs de couleur blanche sont un trait récessif. Le fait que le caractère récessif soit réapparu dans la génération F 2 signifie que les caractères sont restés séparés (et non mélangés) chez les plantes de la génération F 1. Mendel a également suggéré que les plantes possédaient deux copies du caractère caractéristique de la couleur de la fleur et que chaque parent transmettait l'une de ses deux copies à sa progéniture, où elles se réunissaient. De plus, l'observation physique d'un trait dominant peut signifier que la composition génétique de l'organisme comprend deux versions dominantes de la caractéristique ou qu'il comprend une version dominante et une version récessive. À l'inverse, l'observation d'un trait récessif signifiait que l'organisme n'avait aucune version dominante de cette caractéristique.
Alors pourquoi Mendel a-t-il obtenu à plusieurs reprises des ratios de 3:1 dans ses croix ? Pour comprendre comment Mendel a déduit les mécanismes de base de l'héritage qui conduisent à de tels ratios, nous devons d'abord revoir les lois des probabilités.
Notions de base
Les probabilités sont des mesures mathématiques de probabilité. La probabilité empirique d'un événement est calculée en divisant le nombre de fois que l'événement se produit par le nombre total d'opportunités pour que l'événement se produise. Il est également possible de calculer des probabilités théoriques en divisant le nombre de fois qu'un événement devrait se produire par le nombre de fois qu'il pourrait se produire. Les probabilités empiriques proviennent d'observations, comme celles de Mendel. Les probabilités théoriques découlent de la connaissance de la manière dont les événements se produisent et de l'hypothèse que les probabilités des résultats individuels sont égales. Une probabilité de un pour un événement indique qu'il est garanti qu'il se produira, tandis qu'une probabilité de zéro indique qu'il est garanti qu'il ne se produira pas. Un exemple d'événement génétique est la graine ronde produite par un plant de pois. Dans son expérience, Mendel a démontré que la probabilité que l'événement « graine ronde » se produise était celle de la progéniture F1 de parents reproducteurs authentiques, dont l'un possède des graines rondes et l'autre des graines froissées. Lorsque les plants F 1 ont ensuite été autocroisés, la probabilité qu'une progéniture F 2 donnée ait des graines rondes était maintenant de trois sur quatre. En d'autres termes, dans une grande population de descendants F 2 choisis au hasard, 75 pour cent devaient avoir des graines rondes, tandis que 25 pour cent devaient avoir des graines froissées. À l'aide d'un grand nombre de croisements, Mendel a pu calculer des probabilités et les utiliser pour prédire les résultats d'autres croisements.
La règle du produit et la règle de la somme
Mendel a démontré que les caractéristiques des plants de pois qu'il a étudiées étaient transmises sous forme d'unités discrètes du parent à la progéniture. Comme nous le verrons, Mendel a également déterminé que différentes caractéristiques, telles que la couleur et la texture des graines, étaient transmises indépendamment les unes des autres et pouvaient être prises en compte dans des analyses de probabilité distinctes. Par exemple, le croisement entre une plante aux graines vertes et froissées et une plante aux graines jaunes et rondes a tout de même produit une progéniture qui avait un ratio de 3:1 de graines vertes/jaunes (sans tenir compte de la texture des graines) et un ratio de 3:1 de graines rondes/ridées (sans tenir compte de la couleur des graines). Les caractéristiques de couleur et de texture ne s'influençaient pas mutuellement.
La règle de probabilité du produit peut être appliquée à ce phénomène de transmission indépendante de caractéristiques. La règle du produit stipule que la probabilité que deux événements indépendants se produisent simultanément peut être calculée en multipliant les probabilités individuelles que chaque événement se produise seul. Pour démontrer la règle du produit, imaginez que vous lancez une matrice à six faces (D) et que vous retournez un centime (P) en même temps. Le dé peut lancer n'importe quel nombre compris entre 1 et 6 (D #), tandis que la pièce de un cent peut faire tourner les têtes (P H) ou les queues (P T). Le résultat du lancement du dé n'a aucun effet sur le résultat du retournement de la pièce d'un cent et vice versa. Cette action a 12 résultats possibles (tableau\(\PageIndex{2}\)), et chaque événement devrait se produire avec la même probabilité.
| Die à rouler | Retourner Penny |
|---|---|
| D 1 | P H |
| D 1 | JUSQU'À 4 |
| D 2 | P H |
| D 2 | JUSQU'À 4 |
| D 3 | P H |
| D 3 | JUSQU'À 4 |
| D 4 | P H |
| D 4 | JUSQU'À 4 |
| D 5 | P H |
| D 5 | JUSQU'À 4 |
| D 6 | P H |
| D 6 | JUSQU'À 4 |
Sur les 12 résultats possibles, le dé a une probabilité de 2/12 (ou 1/6) de lancer un deux, et le penny a une probabilité de 6/12 (ou 1/2) de monter des têtes. Selon la règle du produit, la probabilité que vous obteniez le résultat combiné 2 et les têtes est la suivante : (D 2) x (P H) = (1/6) x (1/2) ou 1/12 (tableau ci-dessus). Remarquez le mot « et » dans la description de la probabilité. Le « et » indique que vous devez appliquer la règle du produit. Par exemple, considérez comment la règle du produit est appliquée au croisement dihybride : la probabilité de présenter les deux traits dominants chez la descendance F 2 est le produit des probabilités d'avoir le trait dominant pour chaque caractéristique, comme indiqué ici :
D'autre part, la règle de la somme des probabilités est appliquée lorsque l'on considère deux résultats qui s'excluent mutuellement et qui peuvent être obtenus par plus d'une voie. La règle de la somme stipule que la probabilité de survenance d'un événement ou de l'autre événement, de deux événements s'excluant mutuellement, est la somme de leurs probabilités individuelles. Remarquez le mot « ou » dans la description de la probabilité. Le « ou » indique que vous devez appliquer la règle de la somme. Dans ce cas, imaginons que vous retournez un centime (P) et un quart (Q). Quelle est la probabilité qu'une pièce tombe en tête et qu'une pièce arrive en queue ? Ce résultat peut être obtenu dans deux cas : le cent peut être des têtes (P H) et le quart peut être des queues (Q T), ou le quart peut être des têtes (Q H) et le cent peut être des queues (P T). Dans les deux cas, le résultat est atteint. Par la règle de la somme, nous calculons la probabilité d'obtenir une tête et une queue comme
\[\mathrm{[(P_H) × (Q_T)] + [(Q_H) × (P_T)] = [(1/2) × (1/2)] + [(1/2) × (1/2)] = 1/2.}\nonumber\]
Vous devez également noter que nous avons utilisé la règle du produit pour calculer la probabilité de P H et de Q T, ainsi que la probabilité de P T et de Q H, avant de les additionner. Encore une fois, la règle de la somme peut être appliquée pour montrer la probabilité de n'avoir qu'un seul trait dominant dans la génération F 2 d'un croisement dihybride :
\[\frac{3}{16} + \frac{3}{4} = \frac{15}{16}\nonumber\]
| Règle relative au produit | Règle de somme |
|---|---|
| Pour les événements indépendants A et B, la probabilité (P) qu'ils se produisent tous les deux (A et B) est de (P A × P B) | Pour les événements A et B qui s'excluent mutuellement, la probabilité (P) qu'au moins un événement se produise (A ou B) est (P A + P B) |
Pour utiliser les lois de probabilité dans la pratique, il est nécessaire de travailler avec des échantillons de grande taille, car les échantillons de petite taille sont sujets à des écarts dus au hasard. Les grandes quantités de plants de pois examinées par Mendel lui ont permis de calculer les probabilités des caractères apparaissant dans sa génération F 2. Comme vous l'apprendrez, cette découverte signifiait que lorsque les traits parentaux étaient connus, les traits de la progéniture pouvaient être prédits avec précision avant même la fécondation.
Résumé
En travaillant sur des plants de pois de jardin, Mendel a découvert que les croisements entre parents qui différaient par un trait produisaient une progéniture F1 qui exprimait tous les traits d'un parent. Les traits observables sont appelés dominants et les traits non exprimés sont décrits comme récessifs. Lorsque les descendants de l'expérience de Mendel se sont auto-croisés, la progéniture F 2 a présenté le trait dominant ou le trait récessif dans un rapport de 3:1, confirmant que le trait récessif avait été transmis fidèlement par le parent P 0 d'origine. Les croisements réciproques ont généré des rapports de descendance F 1 et F 2 identiques. En examinant la taille des échantillons, Mendel a montré que ses croisements se comportaient de manière reproductible selon les lois des probabilités et que les traits étaient hérités en tant qu'événements indépendants.
Deux règles de probabilité peuvent être utilisées pour déterminer les proportions attendues de descendants de caractères différents issus de différents croisements. Pour déterminer la probabilité que deux événements indépendants ou plus se produisent simultanément, appliquez la règle du produit et multipliez les probabilités des événements individuels. L'utilisation du mot « et » suggère l'application appropriée de la règle du produit. Pour déterminer la probabilité que deux événements ou plus se produisent en combinaison, appliquez la règle de somme et additionnez leurs probabilités individuelles. L'utilisation du mot « ou » suggère l'application appropriée de la règle de la somme.
Notes
- 1 Johann Gregor Mendel, Versuche über Pflanzenhybriden Verhandlungen des naturforschenden Vereines in Brünn, Bd. IV für das Jahr, 1865 Abhandlungen, 3—47. [pour la traduction en anglais, voir www.MendelWeb.org/ Mendel.Plain.html]
Lexique
- théorie du mélange de l'héritage
- modèle héréditaire hypothétique dans lequel les traits parentaux se mélangent chez la progéniture pour produire une apparence physique intermédiaire
- variation continue
- modèle d'héritage dans lequel un personnage présente une gamme de valeurs de caractère avec de petites gradations plutôt que de grands écarts entre elles
- variation discontinue
- modèle d'héritage dans lequel les traits sont distincts et sont transmis indépendamment les uns des autres
- dominant
- trait qui confère la même apparence physique, qu'un individu possède deux copies du trait ou une copie du trait dominant et une copie du trait récessif
- F 1
- première génération filiale issue d'un croisement ; descendance de la génération parentale
- F 2
- deuxième génération filiale produite lorsque les individus F1 se croisent eux-mêmes ou sont fécondés les uns avec les autres
- hybridation
- processus d'accouplement de deux individus qui diffèrent dans le but d'atteindre une certaine caractéristique chez leur progéniture
- système de modèles
- espèce ou système biologique utilisé pour étudier un phénomène biologique spécifique à appliquer à d'autres espèces différentes
- P 0
- génération parentale croisée
- règle du produit
- la probabilité que deux événements indépendants se produisent simultanément peut être calculée en multipliant les probabilités individuelles que chaque événement se produise seul
- récessive
- trait qui apparaît « latent » ou non exprimé lorsque l'individu possède également un trait dominant pour cette même caractéristique ; lorsqu'il est présent sous forme de deux copies identiques, le trait récessif est exprimé
- croix réciproque
- croix appariée dans laquelle les traits respectifs du mâle et de la femelle d'une croix deviennent les traits respectifs de la femelle et du mâle dans l'autre croix
- règle de somme
- la probabilité qu'au moins l'un des deux événements s'excluant mutuellement se produise est la somme de leurs probabilités individuelles
- trait
- variation de l'apparence physique d'une caractéristique héréditaire