
11.1 : Le processus de la méiose
- Page ID
- 189662

Compétences à développer
- Décrire le comportement des chromosomes lors de la méiose
- Décrire les événements cellulaires pendant la méiose
- Expliquer les différences entre la méiose et la mitose
- Expliquer les mécanismes de la méiose qui génèrent des variations génétiques entre les produits de la méiose
La reproduction sexuée nécessite la fécondation, c'est-à-dire l'union de deux cellules provenant de deux organismes individuels. Si ces deux cellules contiennent chacune un ensemble de chromosomes, alors la cellule résultante contient deux ensembles de chromosomes. Les cellules haploïdes contiennent un ensemble de chromosomes. Les cellules contenant deux ensembles de chromosomes sont dites diploïdes. Le nombre d'ensembles de chromosomes dans une cellule est appelé niveau de ploïdie. Si le cycle de reproduction doit se poursuivre, la cellule diploïde doit d'une manière ou d'une autre réduire son nombre d'ensembles chromosomiques avant que la fécondation ne puisse se reproduire, sinon le nombre d'ensembles chromosomiques doublera continuellement à chaque génération. Ainsi, en plus de la fécondation, la reproduction sexuée comprend une division nucléaire qui réduit le nombre d'ensembles de chromosomes.
La plupart des animaux et des plantes sont diploïdes et contiennent deux ensembles de chromosomes. Dans chaque cellule somatique de l'organisme (toutes les cellules d'un organisme multicellulaire à l'exception des gamètes ou des cellules reproductrices), le noyau contient deux copies de chaque chromosome, appelées chromosomes homologues. Les cellules somatiques sont parfois appelées cellules « corporelles ». Les chromosomes homologues sont des paires appariées contenant les mêmes gènes à des emplacements identiques sur leur longueur. Les organismes diploïdes héritent d'une copie de chaque chromosome homologue de chaque parent ; ensemble, ils sont considérés comme un ensemble complet de chromosomes. Les cellules haploïdes, qui contiennent une seule copie de chaque chromosome homologue, ne se trouvent que dans des structures qui donnent naissance à des gamètes ou à des spores. Les spores sont des cellules haploïdes qui peuvent produire un organisme haploïde ou fusionner avec une autre spore pour former une cellule diploïde. Tous les animaux et la plupart des plantes produisent des ovules et des spermatozoïdes, ou gamètes. Certaines plantes et tous les champignons produisent des spores.
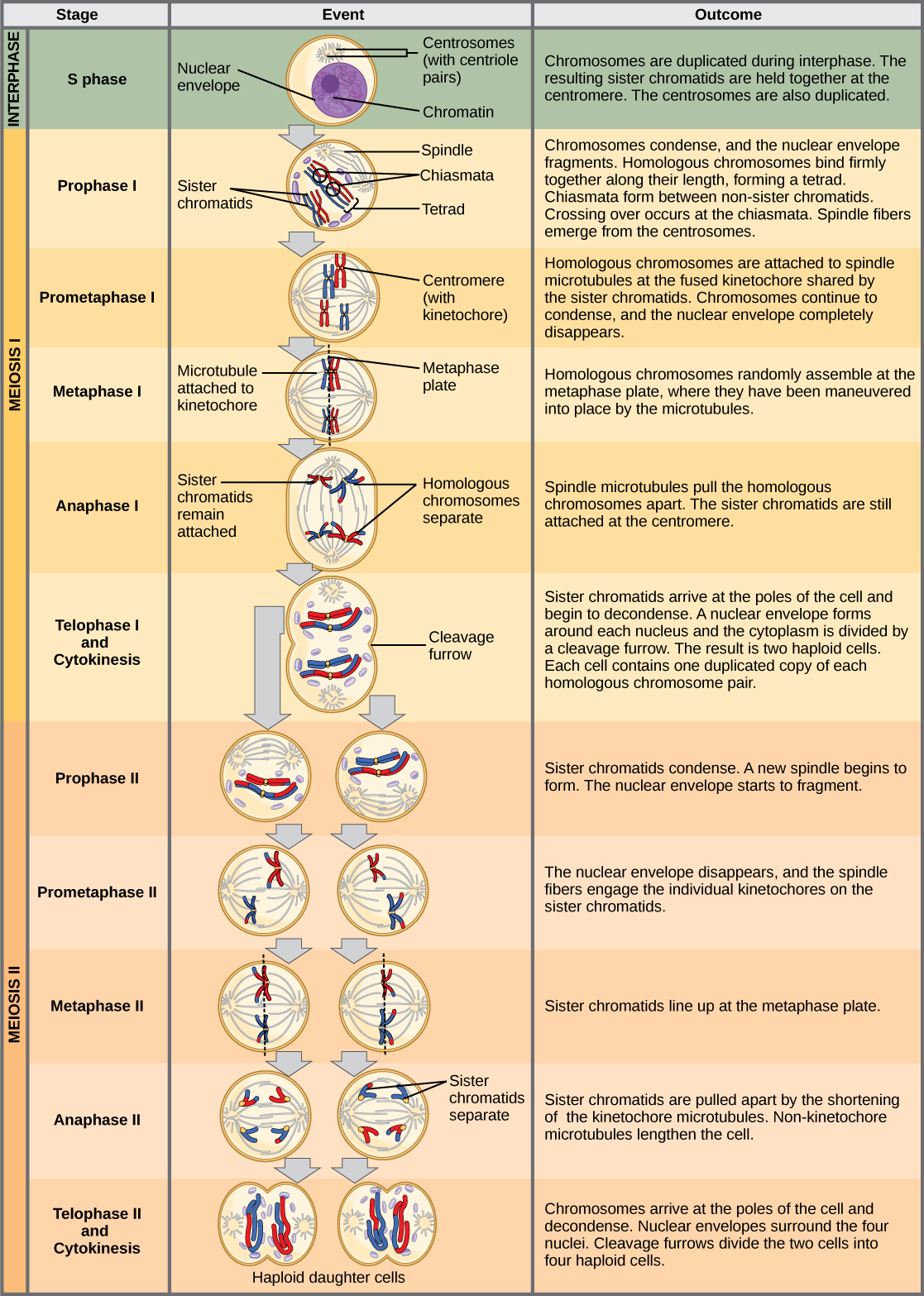
La division nucléaire qui forme les cellules haploïdes, appelée méiose, est liée à la mitose. Comme vous l'avez appris, la mitose est la partie du cycle de reproduction cellulaire qui donne naissance à des noyaux filles identiques qui sont également génétiquement identiques au noyau parent d'origine. En mitose, le noyau parent et le noyau fille ont le même niveau de ploïdie, c'est-à-dire diploïde pour la plupart des plantes et des animaux. La méiose utilise bon nombre des mêmes mécanismes que la mitose. Cependant, le noyau de départ est toujours diploïde et les noyaux qui se forment à la fin d'une division cellulaire méiotique sont haploïdes. Pour obtenir cette réduction du nombre de chromosomes, la méiose consiste en un cycle de duplication des chromosomes et en deux cycles de division nucléaire. Comme les événements qui se produisent au cours de chacun des stades de division sont analogues à ceux de la mitose, les mêmes noms de stade sont attribués. Toutefois, comme il y a deux cycles de division, le processus principal et les étapes sont désignés par un « I » ou un « II ». Ainsi, la méiose I est le premier cycle de division méiotique et comprend la prophase I, la prométaphase I, etc. La méiose II, dans laquelle a lieu le deuxième cycle de division méiotique, comprend la prophase II, la prométaphase II, etc.
Méiose I
La méiose est précédée d'une interphase composée des phases G 1, S et G 2, qui sont presque identiques aux phases précédant la mitose. La phase G 1, également appelée première phase lacunaire, est la première phase de l'interphase et se concentre sur la croissance cellulaire. La phase S est la deuxième phase de l'interphase, au cours de laquelle l'ADN des chromosomes est répliqué. Enfin, la phase G 2, également appelée deuxième phase interstitielle, est la troisième et dernière phase de l'interphase ; au cours de cette phase, la cellule subit les derniers préparatifs pour la méiose.
Lors de la duplication de l'ADN en phase S, chaque chromosome est répliqué pour produire deux copies identiques, appelées chromatides sœurs, qui sont maintenues ensemble au centromère par des protéines de cohésine. La cohésine maintient les chromatides ensemble jusqu'à l'anaphase II. Les centrosomes, qui sont les structures qui organisent les microtubules du fuseau méiotique, se répliquent également. Cela prépare la cellule à entrer dans la prophase I, la première phase méiotique.
Prophase I
Au début de la prophase I, avant que les chromosomes ne soient clairement visibles au microscope, les chromosomes homologues sont attachés à leur extrémité à l'enveloppe nucléaire par des protéines. Lorsque l'enveloppe nucléaire commence à se décomposer, les protéines associées aux chromosomes homologues rapprochent les deux. Rappelons qu'en mitose, les chromosomes homologues ne s'apparient pas. En mitose, les chromosomes homologues s'alignent bout à bout de sorte que, lorsqu'ils se divisent, chaque cellule fille reçoit une chromatide sœur des deux membres de la paire homologue. Le complexe synaptonémal, un réseau de protéines entre les chromosomes homologues, se forme d'abord à des endroits spécifiques, puis s'étend sur toute la longueur des chromosomes. L'appariement serré des chromosomes homologues est appelé synapsis. Dans la synapse, les gènes des chromatides des chromosomes homologues sont exactement alignés les uns avec les autres. Le complexe synaptonémal favorise l'échange de segments chromosomiques entre des chromatides homologues non sœurs, un processus appelé croisement. Le croisement peut être observé visuellement après l'échange sous forme de chiasmas (singulier = chiasma) (Figure\(\PageIndex{1}\)).
Chez des espèces telles que les humains, même si les chromosomes sexuels X et Y ne sont pas homologues (la plupart de leurs gènes diffèrent), ils possèdent une petite région d'homologie qui permet aux chromosomes X et Y de s'apparier pendant la prophase I. Un complexe synaptonémal partiel ne se développe qu'entre les régions d'homologie.
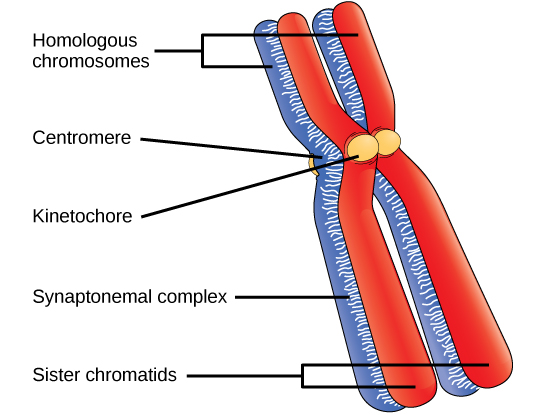
À intervalles réguliers le long du complexe synaptonémal se trouvent de grands assemblages protéiques appelés nodules de recombinaison. Ces assemblages marquent les points des chiasmas ultérieurs et médient le processus en plusieurs étapes de croisement (ou de recombinaison génétique) entre les chromatides non sœurs. Près du nodule de recombinaison de chaque chromatide, l'ADN bicaténaire est clivé, les extrémités coupées sont modifiées et une nouvelle connexion est établie entre les chromatides non sœurs. Au fur et à mesure que la prophase I progresse, le complexe synaptonémal commence à se décomposer et les chromosomes commencent à se condenser. Lorsque le complexe synaptonémal disparaît, les chromosomes homologues restent attachés les uns aux autres au niveau du centromère et des chiasmas. Les chiasmas subsistent jusqu'à l'anaphase I. Le nombre de chiasmas varie en fonction de l'espèce et de la longueur du chromosome. Il doit y avoir au moins un chiasma par chromosome pour séparer correctement les chromosomes homologues au cours de la méiose I, mais il peut y en avoir jusqu'à 25. Après le croisement, le complexe synaptonémal se décompose et la liaison de cohésine entre les paires homologues est également supprimée. À la fin de la prophase I, les paires sont maintenues ensemble uniquement au niveau des chiasmes (Figure\(\PageIndex{2}\)) et sont appelées tétrades parce que les quatre chromatides sœurs de chaque paire de chromosomes homologues sont désormais visibles.
Les événements de croisement sont la première source de variation génétique des noyaux produits par la méiose. Un seul événement de croisement entre des chromatides homologues non sœurs entraîne un échange réciproque d'ADN équivalent entre un chromosome maternel et un chromosome paternel. Maintenant, lorsque cette chromatide sœur est déplacée dans une cellule gamétique, elle transportera de l'ADN d'un parent de l'individu et de l'ADN de l'autre parent. La chromatide recombinante sœur possède une combinaison de gènes maternels et paternels qui n'existaient pas avant le croisement. De multiples croisements dans un bras du chromosome ont le même effet, échangeant des segments d'ADN pour créer des chromosomes recombinants.
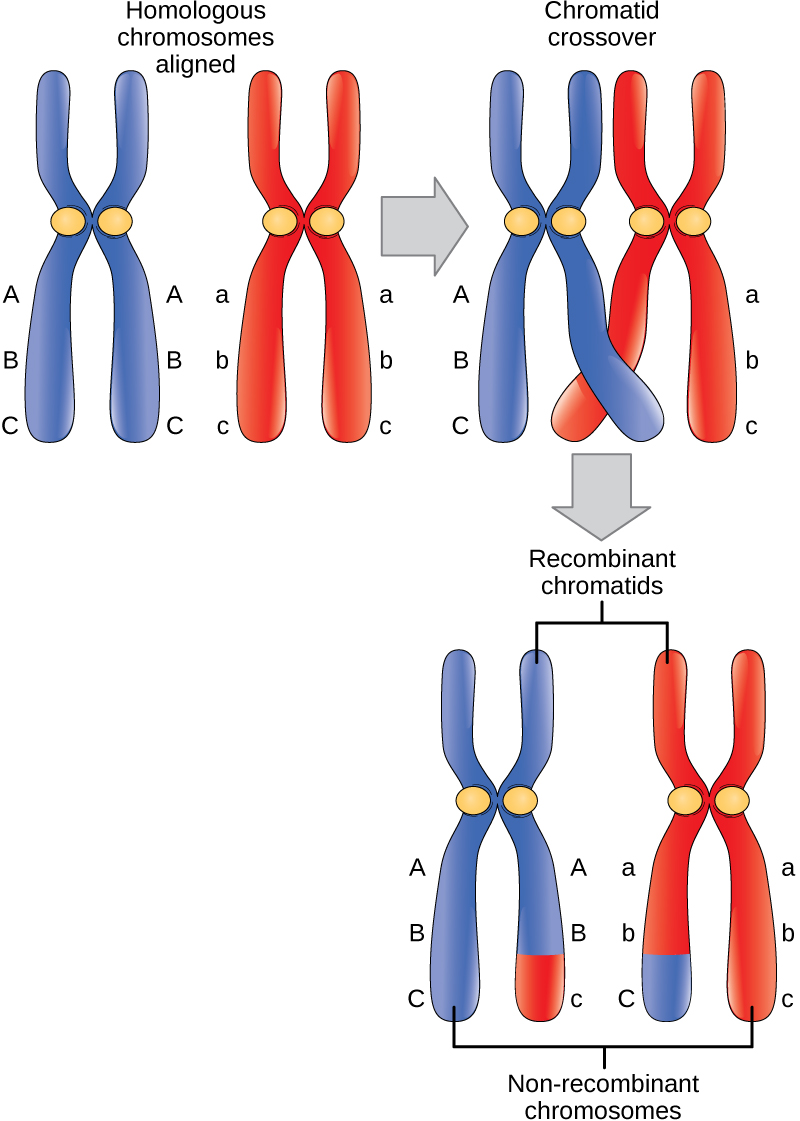
Prométaphase I
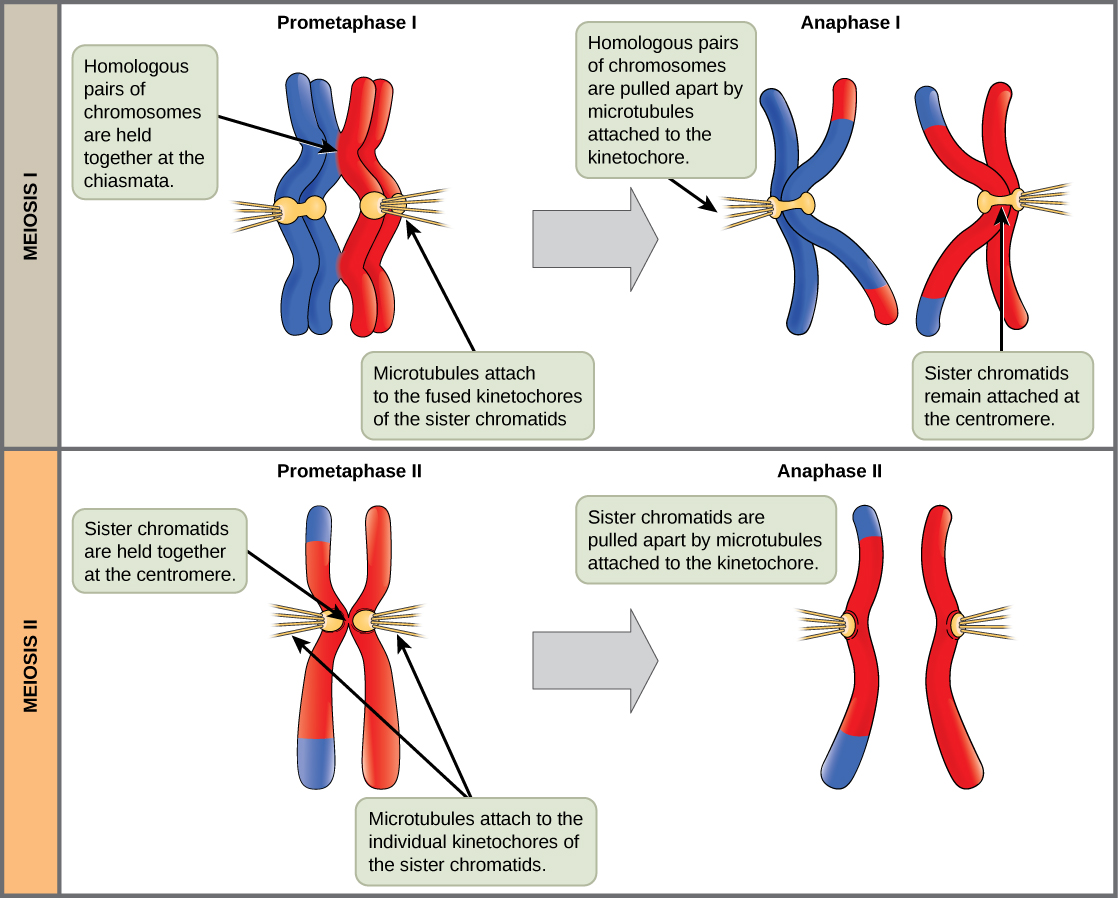
L'événement clé de la prométaphase I est la fixation des microtubules des fibres fusiformes aux protéines kinétochores au niveau des centromères. Les protéines kinétochores sont des complexes multiprotéiques qui lient les centromères d'un chromosome aux microtubules du fuseau mitotique. Les microtubules se développent à partir de centrosomes placés aux pôles opposés de la cellule. Les microtubules se déplacent vers le milieu de la cellule et se fixent à l'un des deux chromosomes homologues fusionnés. Les microtubules se fixent au niveau des kinétochores de chaque chromosome. Chaque membre de la paire homologue étant attaché aux pôles opposés de la cellule, lors de la phase suivante, les microtubules peuvent séparer la paire homologue. Une fibre fusiforme qui s'est attachée à un kinétochore est appelée microtubule kinétochore. À la fin de la prométaphase I, chaque tétrade est attachée à des microtubules à partir des deux pôles, un chromosome homologue faisant face à chaque pôle. Les chromosomes homologues sont toujours maintenus ensemble au niveau des chiasmas. De plus, la membrane nucléaire s'est complètement décomposée.
Métaphase I
Au cours de la métaphase I, les chromosomes homologues sont disposés au centre de la cellule, les kinétochores faisant face à des pôles opposés. Les paires homologues s'orientent aléatoirement à l'équateur. Par exemple, si les deux membres homologues du chromosome 1 sont marqués a et b, alors les chromosomes peuvent s'aligner sur a-b ou b-a. Ceci est important pour déterminer les gènes portés par un gamète, car chacun ne recevra qu'un seul des deux chromosomes homologues. Rappelons que les chromosomes homologues ne sont pas identiques. Ils présentent de légères différences dans leurs informations génétiques, ce qui confère à chaque gamète une constitution génétique unique.
Ce caractère aléatoire est la base physique de la création de la deuxième forme de variation génétique chez la progéniture. Considérez que les chromosomes homologues d'un organisme se reproduisant sexuellement sont hérités à l'origine sous la forme de deux ensembles distincts, un de chaque parent. Si l'on prend l'exemple des humains, un ensemble de 23 chromosomes est présent dans l'ovule donné par la mère. Le père fournit l'autre ensemble de 23 chromosomes dans le sperme qui féconde l'ovule. Chaque cellule de la progéniture multicellulaire possède des copies des deux ensembles originaux de chromosomes homologues. Dans la prophase I de la méiose, les chromosomes homologues forment les tétrades. Dans la métaphase I, ces paires s'alignent à mi-chemin entre les deux pôles de la cellule pour former la plaque de métaphase. Comme il existe une chance égale qu'une fibre microtubulaire rencontre un chromosome héréditaire maternel ou paternel, la disposition des tétrades sur la plaque de métaphase est aléatoire. Tout chromosome héréditaire maternel peut faire face à l'un ou l'autre pôle. Tout chromosome héréditaire peut également faire face à l'un ou l'autre des pôles. L'orientation de chaque tétrade est indépendante de l'orientation des 22 autres tétrades.
Cet événement, l'assortiment aléatoire (ou indépendant) de chromosomes homologues sur la plaque de métaphase, est le deuxième mécanisme qui introduit une variation dans les gamètes ou les spores. Dans chaque cellule soumise à une méiose, la disposition des tétrades est différente. Le nombre de variations dépend du nombre de chromosomes composant un ensemble. Il existe deux possibilités d'orientation au niveau de la plaque métaphasique ; le nombre possible d'alignements est donc égal à 2 n, où n est le nombre de chromosomes par ensemble. Les humains possèdent 23 paires de chromosomes, ce qui donne plus de huit millions (2 23) de gamètes génétiquement distincts. Ce nombre n'inclut pas la variabilité qui a été créée précédemment dans les chromatides sœurs par croisement. Compte tenu de ces deux mécanismes, il est très peu probable que deux cellules haploïdes issues de la méiose aient la même composition génétique (Figure\(\PageIndex{3}\)).
Pour résumer les conséquences génétiques de la méiose I, les gènes maternel et paternel sont recombinés par des événements croisés qui se produisent entre chaque paire homologue au cours de la prophase I. De plus, l'assortiment aléatoire de tétrades sur la plaque de métaphase produit une combinaison unique de chromosomes maternels et paternels. qui se retrouveront dans les gamètes.
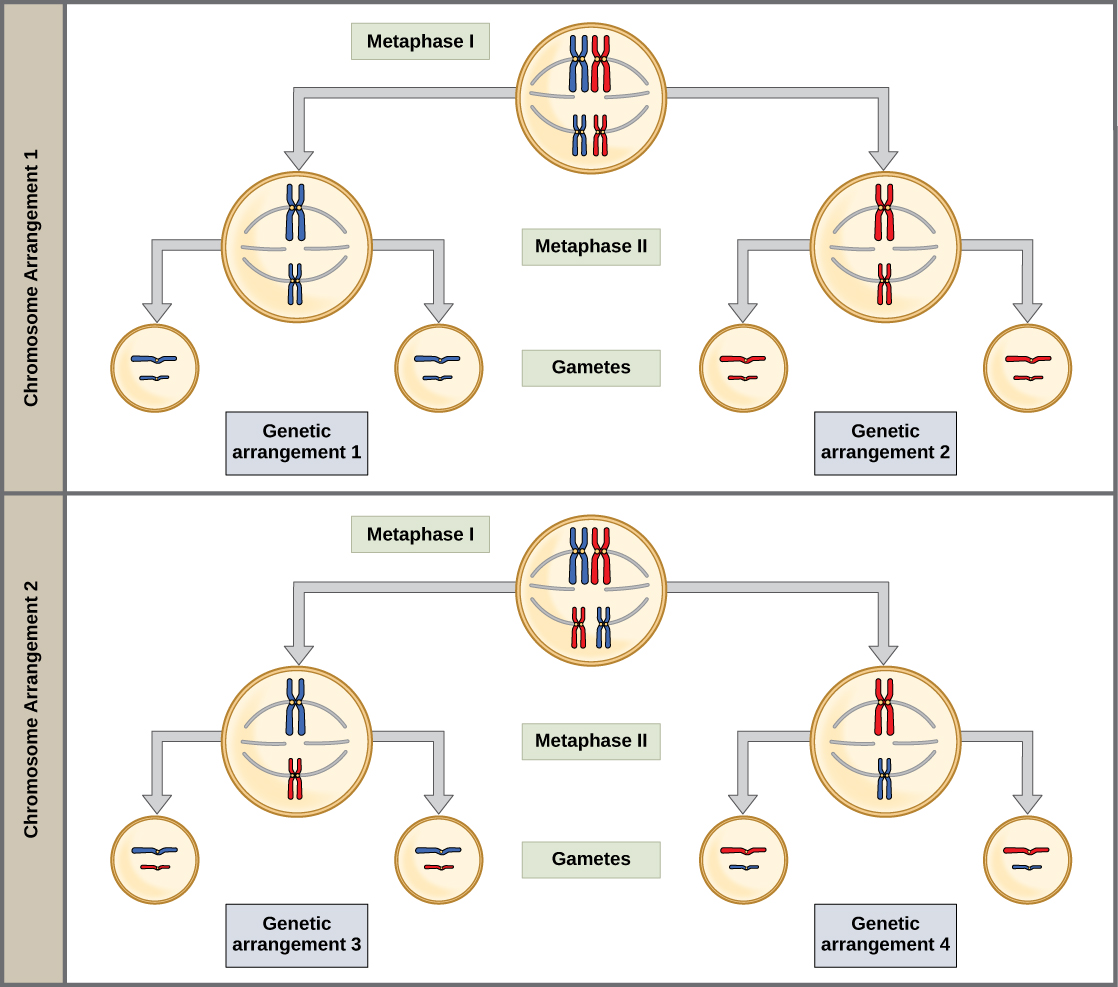
Anaphase I
Dans l'anaphase I, les microtubules séparent les chromosomes liés. Les chromatides sœurs restent étroitement liées entre elles au centromère. Les chiasmas sont cassés en anaphase I lorsque les microtubules attachés aux kinétochores fusionnés séparent les chromosomes homologues (Figure\(\PageIndex{4}\)).
Télophase I et cytokinèse
En télophase, les chromosomes séparés arrivent à des pôles opposés. Le reste des épisodes de télophase typiques peuvent se produire ou ne pas se produire, selon l'espèce. Chez certains organismes, les chromosomes se décondensent et des enveloppes nucléaires se forment autour des chromatides dans la télophase I. Dans d'autres organismes, la cytokinèse, c'est-à-dire la séparation physique des composants cytoplasmiques en deux cellules filles, se produit sans reformation des noyaux. Chez presque toutes les espèces animales et chez certains champignons, la cytokinèse sépare le contenu cellulaire par un sillon de clivage (constriction de l'anneau d'actine qui entraîne la division cytoplasmique). Chez les plantes, une plaque cellulaire est formée au cours de la cytokinèse cellulaire par la fusion de vésicules de Golgi au niveau de la plaque métaphasique. Cette plaque cellulaire conduira finalement à la formation de parois cellulaires qui séparent les deux cellules filles.
Deux cellules haploïdes sont le résultat final de la première division méiotique. Les cellules sont haploïdes car à chaque pôle, il n'y a qu'un seul chromosome homologue de chaque paire. Par conséquent, un seul ensemble complet de chromosomes est présent. C'est pourquoi les cellules sont considérées comme haploïdes : il n'existe qu'un seul ensemble de chromosomes, même si chaque homologue est toujours composé de deux chromatides sœurs. Rappelons que les chromatides sœurs ne sont que des doublons de l'un des deux chromosomes homologues (à l'exception des modifications survenues lors du croisement). Dans la méiose II, ces deux chromatides sœurs se sépareront, créant ainsi quatre cellules filles haploïdes.
Lien vers l'apprentissage

Passez en revue le processus de la méiose, en observant comment les chromosomes s'alignent et migrent, sur Meiosis : An Interactive Animation.
Méiose II
Chez certaines espèces, les cellules entrent dans une brève interphase, ou interkinésie, avant d'entrer dans la méiose II. L'interkinésie n'a pas de phase S, de sorte que les chromosomes ne sont pas dupliqués. Les deux cellules produites lors de la méiose I traversent les événements de la méiose II de manière synchronisée. Au cours de la méiose II, les chromatides sœurs des deux cellules filles se séparent, formant quatre nouveaux gamètes haploïdes. La mécanique de la méiose II est similaire à celle de la mitose, sauf que chaque cellule en division possède un seul ensemble de chromosomes homologues. Chaque cellule possède donc la moitié du nombre de chromatides sœurs à séparer pour former une cellule diploïde en mitose.
Prophase II
Si les chromosomes se décondensent en télophase I, ils se condensent à nouveau. Si des enveloppes nucléaires se forment, elles se fragmentent en vésicules. Les centrosomes qui ont été dupliqués pendant l'interkinésie s'éloignent l'un de l'autre vers des pôles opposés, et de nouveaux fuseaux se forment.
Prométaphase II
Les enveloppes nucléaires sont complètement cassées et la broche est complètement formée. Chaque chromatide sœur forme un kinétochore individuel qui se fixe aux microtubules à partir de pôles opposés.
Métaphase II
Les chromatides sœurs sont condensées au maximum et alignées à l'équateur de la cellule.
Anaphase II
Les chromatides sœurs sont séparées par les microtubules du kinétochore et se déplacent vers des pôles opposés. Les microtubules non kinétochores allongent la cellule.
Télophase II et cytokinèse
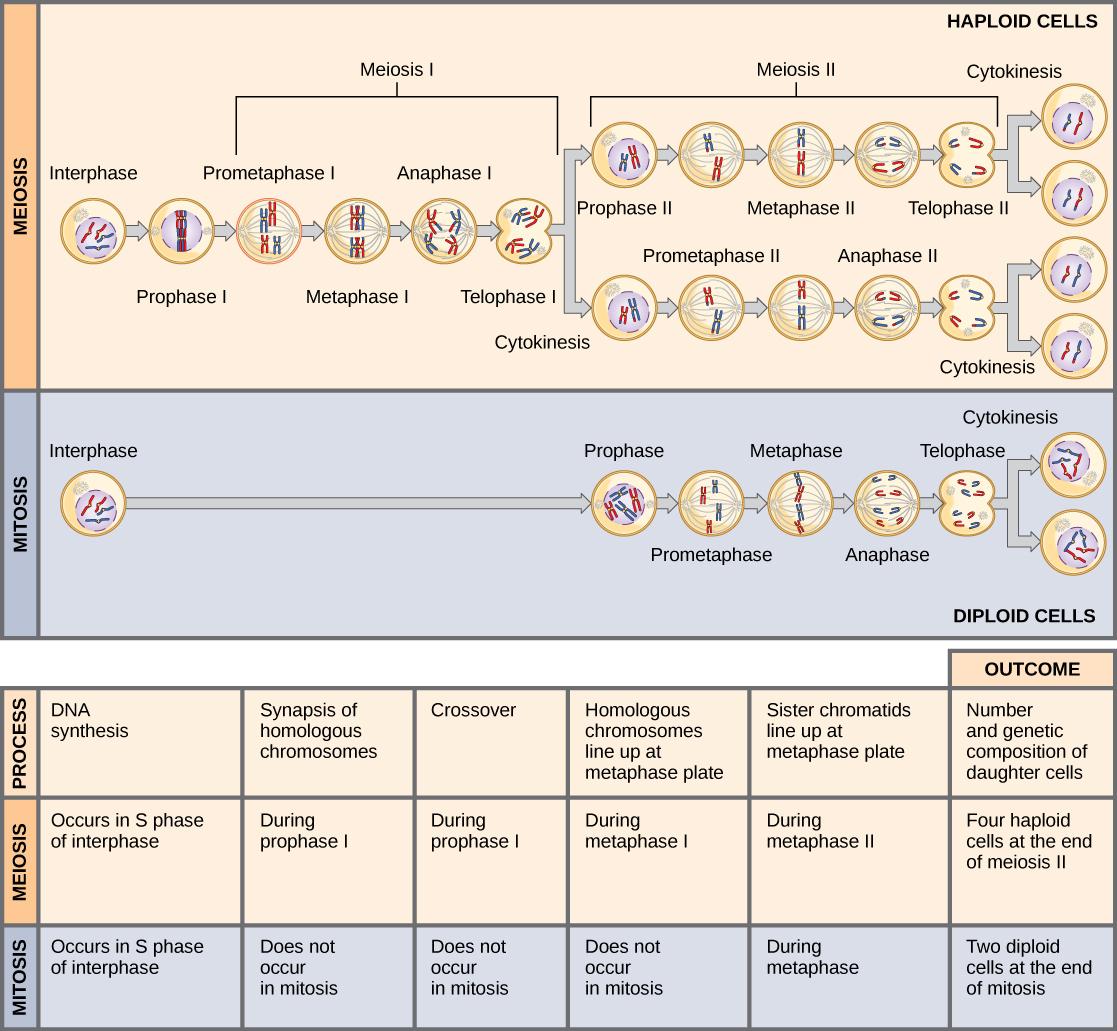
Les chromosomes arrivent aux pôles opposés et commencent à se décondenser. Des enveloppes nucléaires se forment autour des chromosomes. La cytokinèse sépare les deux cellules en quatre cellules haploïdes uniques. À ce stade, les noyaux nouvellement formés sont tous deux haploïdes. Les cellules produites sont génétiquement uniques en raison de l'assortiment aléatoire d'homologues paternels et maternels et de la recombinaison des segments maternels et paternels des chromosomes (avec leurs ensembles de gènes) qui se produit lors du croisement. L'ensemble du processus de méiose est décrit dans la figure\(\PageIndex{5}\).
Comparaison de la méiose et de la mitose
La mitose et la méiose sont deux formes de division du noyau dans les cellules eucaryotes. Ils partagent certaines similitudes, mais présentent également des différences distinctes qui se traduisent par des résultats très différents (Figure\(\PageIndex{6}\)). La mitose est une division nucléaire unique qui donne naissance à deux noyaux qui sont généralement divisés en deux nouvelles cellules. Les noyaux résultant d'une division mitotique sont génétiquement identiques au noyau d'origine. Elles possèdent le même nombre d'ensembles de chromosomes, un ensemble dans le cas des cellules haploïdes et deux ensembles dans le cas des cellules diploïdes. Dans la plupart des plantes et toutes les espèces animales, ce sont généralement les cellules diploïdes qui subissent une mitose pour former de nouvelles cellules diploïdes. En revanche, la méiose se compose de deux divisions nucléaires résultant en quatre noyaux qui sont généralement divisés en quatre nouvelles cellules. Les noyaux issus de la méiose ne sont pas génétiquement identiques et ne contiennent qu'un seul ensemble de chromosomes. Cela représente la moitié du nombre d'ensembles chromosomiques de la cellule d'origine, qui est diploïde.
Les principales différences entre la mitose et la méiose se situent dans la méiose I, qui est une division nucléaire très différente de la mitose. Dans la méiose I, les paires de chromosomes homologues s'associent les unes aux autres, se lient au complexe synaptonémal, développent des chiasmas et subissent un croisement entre des chromatides sœurs, et s'alignent le long de la plaque métaphasique en tétrades avec des fibres kinétochores provenant de pôles fusiformes opposés attachés à chaque kinétochore d'un homologue dans une tétrade. Tous ces événements ne se produisent que dans la méiose I.
Lorsque les chiasmas disparaissent et que la tétrade est brisée et que les homologues se déplacent vers un pôle ou un autre, le niveau de ploïdie, c'est-à-dire le nombre d'ensembles de chromosomes dans chaque noyau futur, est réduit de deux à un. Pour cette raison, la méiose I est appelée division de réduction. Il n'y a pas de telle réduction du niveau de ploïdie pendant la mitose.
La méiose II ressemble beaucoup plus à une division mitotique. Dans ce cas, les chromosomes dupliqués (un seul ensemble d'entre eux) s'alignent sur la plaque de métaphase avec des kinétochores divisés attachés aux fibres kinétochores à partir de pôles opposés. Au cours de l'anaphase II, comme dans l'anaphase mitotique, les kinétochores se divisent et une chromatide sœur, maintenant appelée chromosome, est tirée vers un pôle tandis que l'autre chromatide sœur est tirée vers l'autre pôle. S'il n'y avait pas eu de croisement, les deux produits de chaque division individuelle de la méiose II seraient identiques (comme dans le cas de la mitose). Ils sont plutôt différents parce qu'il y a toujours eu au moins un croisement par chromosome. La méiose II n'est pas une division de réduction car, bien qu'il y ait moins de copies du génome dans les cellules résultantes, il existe toujours un ensemble de chromosomes, comme c'était le cas à la fin de la méiose I.
Evolution Connection : Le mystère de l'évolution de la méiose
Certaines caractéristiques des organismes sont si répandues et fondamentales qu'il est parfois difficile de se rappeler qu'elles ont évolué comme d'autres traits plus simples. La méiose est une série d'événements cellulaires d'une telle complexité que les biologistes ont eu du mal à émettre des hypothèses et à tester son évolution. Bien que la méiose soit inextricablement liée à la reproduction sexuée et à ses avantages et inconvénients, il est important de séparer les questions de l'évolution de la méiose et de l'évolution du sexe, car une méiose précoce peut avoir été avantageuse pour des raisons différentes de celles qu'elle ne l'est actuellement. Sortir des sentiers battus et imaginer quels auraient pu être les premiers avantages de la méiose est une approche pour découvrir comment elle a pu évoluer.
La méiose et la mitose partagent des processus cellulaires évidents et il est logique que la méiose ait évolué à partir de la mitose. La difficulté réside dans les différences claires entre la méiose I et la mitose. Adam Wilkins et Robin Holliday 1 ont résumé les événements uniques qui devaient se produire pour l'évolution de la méiose à la suite d'une mitose. Ces étapes sont l'appariement des chromosomes homologues, les échanges croisés, le maintien des chromatides sœurs pendant l'anaphase et la suppression de la réplication de l'ADN en interphase. Ils soutiennent que la première étape est la plus difficile et la plus importante, et que comprendre comment elle a évolué permettrait de clarifier le processus évolutif. Ils proposent des expériences génétiques qui pourraient permettre de mieux comprendre l'évolution des synapses.
Il existe d'autres approches en cours pour comprendre l'évolution de la méiose. Différentes formes de méiose existent chez les protistes unicellulaires. Certaines semblent être des formes de méiose plus simples ou plus « primitives ». La comparaison des divisions méiotiques de différents protistes peut permettre de mieux comprendre l'évolution de la méiose. Marilee Ramesh et ses collègues 2 ont comparé les gènes impliqués dans la méiose chez les protistes afin de comprendre quand et où la méiose a pu évoluer. Bien que les recherches soient toujours en cours, des études récentes sur la méiose chez les protistes suggèrent que certains aspects de la méiose ont peut-être évolué plus tard que d'autres. Ce type de comparaison génétique peut nous indiquer quels aspects de la méiose sont les plus anciens et quels processus cellulaires ils ont pu emprunter dans des cellules antérieures.
Résumé
La reproduction sexuée exige que les organismes diploïdes produisent des cellules haploïdes qui peuvent fusionner pendant la fécondation pour former une progéniture diploïde. Comme pour la mitose, la réplication de l'ADN se produit avant la méiose pendant la phase S du cycle cellulaire. La méiose est une série d'événements qui organisent et séparent les chromosomes et les chromatides en cellules filles. Au cours des interphases de la méiose, chaque chromosome est dupliqué. Dans la méiose, il y a deux cycles de division nucléaire qui donnent quatre noyaux et généralement quatre cellules filles, chacune ayant deux fois moins de chromosomes que la cellule mère. La première sépare les homologues et la seconde, comme une mitose, sépare les chromatides en chromosomes individuels. Au cours de la méiose, une variation des noyaux filles est introduite en raison du croisement de la prophase I et de l'alignement aléatoire des tétrades à la métaphase I. Les cellules produites par la méiose sont génétiquement uniques.
La méiose et la mitose présentent des similitudes, mais ont des résultats distincts. Les divisions mitotiques sont des divisions nucléaires uniques qui produisent des noyaux filles génétiquement identiques et qui possèdent le même nombre d'ensembles chromosomiques que la cellule d'origine. Les divisions méiotiques comprennent deux divisions nucléaires qui produisent quatre noyaux filles génétiquement différents et qui possèdent un ensemble de chromosomes au lieu des deux ensembles de chromosomes de la cellule mère. Les principales différences entre les processus se situent dans la première division de la méiose, dans laquelle des chromosomes homologues sont appariés et échangent des segments de chromatides non sœurs. Les chromosomes homologues se séparent en différents noyaux au cours de la méiose I, provoquant une réduction du niveau de ploïdie dans la première division. La deuxième division de la méiose ressemble davantage à la division mitotique, sauf que les cellules filles ne contiennent pas de génomes identiques en raison du croisement.
Notes
- 1 Adam S. Wilkins et Robin Holliday, « L'évolution de la méiose à partir de la mitose », Genetics 181 (2009) : 3—12.
- 2 Marilee A. Ramesh, Shehre-Banoo Malik et John M. Logsdon, Jr, « Un inventaire phylogénétique des gènes méiotiques : preuves du sexe chez Giardia et origine eucaryote précoce de la méiose », Current Biology 15 (2005) :185—91.
Lexique
- chiasmata
- (singulier, chiasma) structure qui se forme aux points de croisement après l'échange de matériel génétique
- cohésine
- protéines qui forment un complexe qui scelle les chromatides sœurs au niveau de leurs centromères jusqu'à l'anaphase II de la méiose
- croisement
- échange de matériel génétique entre des chromatides non sœurs, aboutissant à des chromosomes qui incorporent des gènes des deux parents de l'organisme
- fertilisation
- union de deux cellules haploïdes provenant de deux organismes individuels
- interkinésie
- (également, interphase II) brève période de repos entre la méiose I et la méiose II
- méiose
- un processus de division nucléaire qui donne naissance à quatre cellules haploïdes
- méiose I
- premier cycle de division cellulaire méiotique ; appelé division de réduction parce que le niveau de ploïdie est réduit de diploïde à haploïde
- méiose II
- deuxième cycle de division cellulaire méiotique après la méiose I ; les chromatides sœurs sont séparées en chromosomes individuels, ce qui donne quatre cellules haploïdes uniques
- nodules de recombinaison
- assemblages de protéines formés sur le complexe synaptonémal qui marquent les points des événements de croisement et assurent la médiation du processus en plusieurs étapes de recombinaison génétique entre des chromatides non sœurs
- division de réduction
- division nucléaire qui produit des noyaux filles contenant chacun deux fois moins d'ensembles chromosomiques que le noyau parental ; la méiose I est une division de réduction
- cellule somatique
- toutes les cellules d'un organisme multicellulaire à l'exception des gamètes ou des cellules reproductrices
- spore
- cellule haploïde capable de produire un organisme multicellulaire haploïde ou de fusionner avec une autre spore pour former une cellule diploïde
- synapsis
- formation d'une association étroite entre des chromosomes homologues au cours de la prophase I
- complexe synaptonémal
- réseau protéique qui se forme entre les chromosomes homologues au cours de la prophase I, favorisant le croisement
- tétrade
- deux chromosomes homologues dupliqués (quatre chromatides) liés entre eux par des chiasmas au cours de la prophase I