
24.2 : Métabolisme des glucides
- Page ID
- 195172

Objectifs d'apprentissage
- Expliquer les processus de glycolyse
- Décrire le parcours d'une molécule de pyruvate dans le cycle de Krebs
- Expliquer le transport des électrons à travers la chaîne de transport des électrons
- Décrire le processus de production d'ATP par phosphorylation oxydative
- Résumez le processus de la gluconéogenèse
Les glucides sont des molécules organiques composées d'atomes de carbone, d'hydrogène et d'oxygène. La famille des glucides comprend des sucres simples et complexes. Le glucose et le fructose sont des exemples de sucres simples, tandis que l'amidon, le glycogène et la cellulose sont tous des exemples de sucres complexes. Les sucres complexes sont également appelés polysaccharides et sont composés de plusieurs molécules de monosaccharides. Les polysaccharides servent de stockage d'énergie (par exemple, amidon et glycogène) et de composants structuraux (par exemple, la chitine chez les insectes et la cellulose chez les plantes).
Au cours de la digestion, les glucides sont décomposés en sucres simples et solubles qui peuvent être transportés à travers la paroi intestinale jusqu'au système circulatoire pour être transportés dans tout le corps. La digestion des glucides commence par la bouche sous l'action de l'amylase salivaire sur les amidons et se termine par l'absorption des monosaccharides à travers l'épithélium de l'intestin grêle. Une fois que les monosaccharides absorbés sont transportés vers les tissus, le processus de respiration cellulaire commence (Figure\(\PageIndex{1}\)). Cette section se concentrera d'abord sur la glycolyse, un processus par lequel le glucose monosaccharide est oxydé, libérant ainsi l'énergie stockée dans ses liaisons pour produire de l'ATP.

Glycolyse
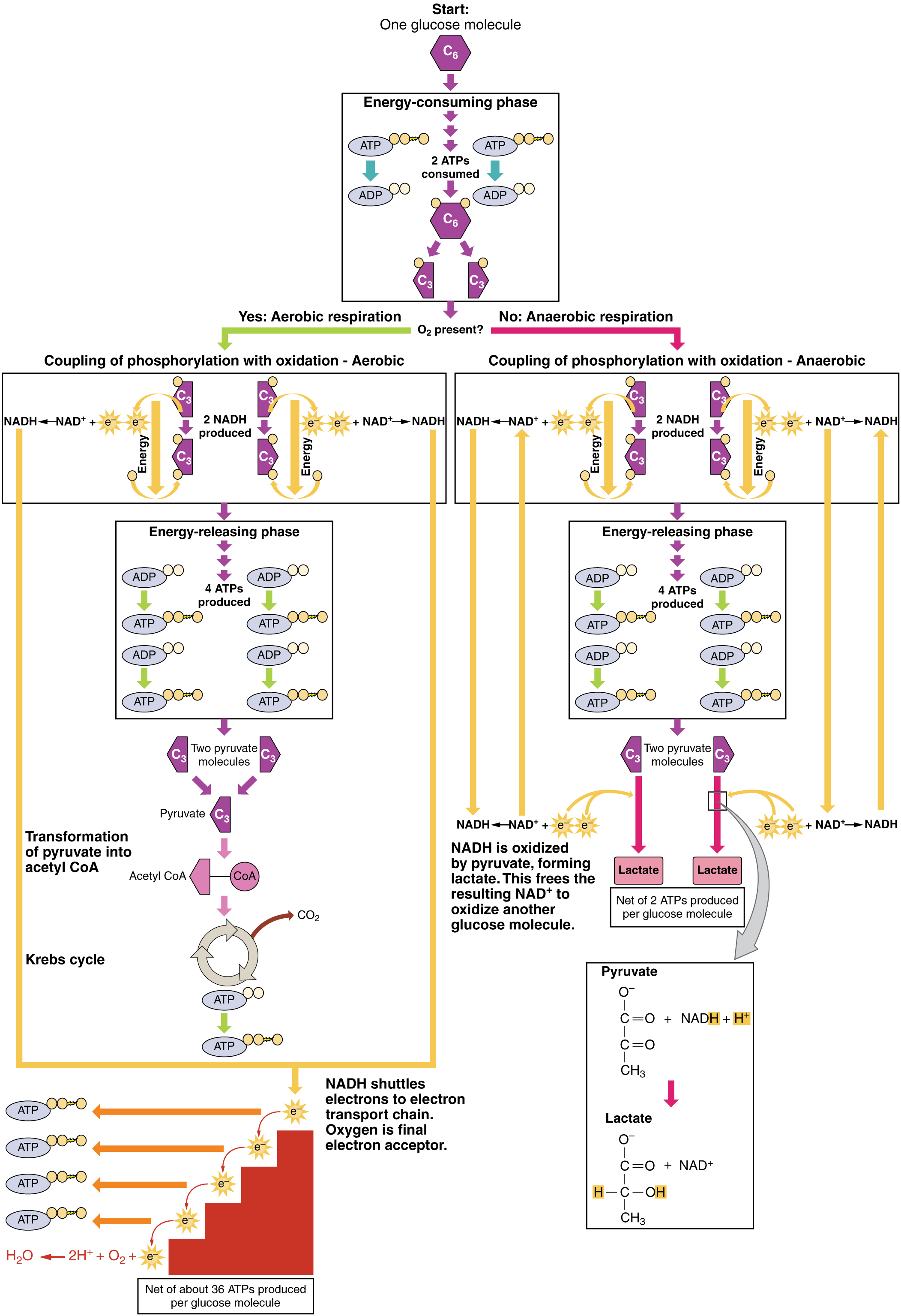
Le glucose est la source d'énergie la plus facilement disponible de l'organisme. Une fois que les processus digestifs ont décomposé les polysaccharides en monosaccharides, y compris le glucose, les monosaccharides sont transportés à travers la paroi de l'intestin grêle et dans le système circulatoire, qui les transporte vers le foie. Dans le foie, les hépatocytes transmettent le glucose par le système circulatoire ou stockent l'excès de glucose sous forme de glycogène. Les cellules du corps absorbent le glucose circulant en réponse à l'insuline et, par le biais d'une série de réactions appelées glycolyse, transfèrent une partie de l'énergie du glucose à l'ADP pour former de l'ATP (Figure\(\PageIndex{2}\)). La dernière étape de la glycolyse produit le pyruvate.
La glycolyse commence par la phosphorylation du glucose par l'hexokinase pour former du glucose-6-phosphate. Cette étape utilise un ATP, qui est le donneur du groupe phosphate. Sous l'action de la phosphofructokinase, le glucose-6-phosphate est transformé en fructose-6-phosphate. À ce stade, un deuxième ATP donne son groupe phosphate, formant du fructose-1,6-bisphosphate. Ce sucre à six carbones est divisé pour former deux molécules à trois carbones phosphorylées, le glycérodéhyde-3-phosphate et le phosphate de dihydroxyacétone, qui sont toutes deux converties en glycérodéhyde-3-phosphate. Le glycérodéhyde-3-phosphate est ensuite phosphorylé avec des groupes donnés par le dihydrogénophosphate présent dans la cellule pour former la molécule à trois carbones 1,3-bisphosphoglycérate. L'énergie de cette réaction provient de l'oxydation du glycérodéhyde-3-phosphate (élimination des électrons du) glycéraldéhyde-3-phosphate. Dans une série de réactions conduisant au pyruvate, les deux groupes phosphates sont ensuite transférés vers deux ADP pour former deux ATP. Ainsi, la glycolyse utilise deux ATP mais génère quatre ATP, ce qui donne un gain net de deux ATP et de deux molécules de pyruvate. En présence d'oxygène, le pyruvate poursuit son cours jusqu'au cycle de Krebs (également appelé cycle de l'acide citrique ou cycle de l'acide tricarboxylique (TCA), où de l'énergie supplémentaire est extraite et transmise.


La glycolyse peut être divisée en deux phases : consommation d'énergie (également appelée amorçage chimique) et production d'énergie. La première phase est la phase consommatrice d'énergie, elle nécessite donc deux molécules d'ATP pour démarrer la réaction pour chaque molécule de glucose. Cependant, la fin de la réaction produit quatre ATP, ce qui se traduit par un gain net de deux molécules d'énergie ATP.
La glycolyse peut être exprimée par l'équation suivante :
Glucose + 2ATP + 2NAD + + 4ADP + P2 i → 2 Pyruvate + 4ATP + 2NADH + 2H+
Cette équation indique que le glucose, en combinaison avec l'ATP (la source d'énergie), le NAD+ (une coenzyme qui sert d'accepteur d'électrons) et le phosphate inorganique, se décompose en deux molécules de pyruvate, générant quatre molécules d'ATP, pour un rendement net de deux ATP, et deux coenzymes NADH contenant de l'énergie. Le NADH produit au cours de ce processus sera utilisé ultérieurement pour produire de l'ATP dans les mitochondries. Il est important de noter qu'à la fin de ce processus, une molécule de glucose génère deux molécules de pyruvate, deux molécules d'ATP à haute énergie et deux molécules de NADH porteuses d'électrons.
Les discussions suivantes sur la glycolyse incluent les enzymes responsables des réactions. Lorsque le glucose entre dans une cellule, l'enzyme hexokinase (ou glucokinase, présente dans le foie) ajoute rapidement un phosphate pour le convertir en glucose-6-phosphate. Une kinase est un type d'enzyme qui ajoute une molécule de phosphate à un substrat (dans ce cas, du glucose, mais cela peut également être vrai pour d'autres molécules). Cette étape de conversion nécessite un ATP et piège essentiellement le glucose dans la cellule, l'empêchant ainsi de revenir à travers la membrane plasmique, permettant ainsi à la glycolyse de se poursuivre. Il permet également de maintenir un gradient de concentration avec des taux de glucose plus élevés dans le sang que dans les tissus. En établissant ce gradient de concentration, le glucose dans le sang pourra s'écouler d'une zone de forte concentration (le sang) vers une zone de faible concentration (les tissus) pour être utilisé ou stocké. L'hexokinase est présente dans presque tous les tissus du corps. La glucokinase, quant à elle, est exprimée dans les tissus actifs lorsque la glycémie est élevée, tels que le foie. L'hexokinase a une affinité plus élevée pour le glucose que la glucokinase et est donc capable de convertir le glucose plus rapidement que la glucokinase. Cela est important lorsque le taux de glucose est très bas dans l'organisme, car cela permet au glucose de se déplacer de préférence vers les tissus qui en ont le plus besoin.
À l'étape suivante de la première phase de glycolyse, l'enzyme glucose-6-phosphate isomérase convertit le glucose-6-phosphate en fructose-6-phosphate. Comme le glucose, le fructose est également un sucre contenant six carbones. L'enzyme phosphofructokinase-1 ajoute ensuite un phosphate supplémentaire pour convertir le fructose-6-phosphate en fructose-1-6-bisphosphate, un autre sucre à six carbones, en utilisant une autre molécule d'ATP. L'aldolase décompose ensuite ce fructose-1-6-bisphosphate en deux molécules à trois carbones, le glycérodéhyde-3-phosphate et le phosphate de dihydroxyacétone. L'enzyme triosephosphate isomérase convertit ensuite le phosphate de dihydroxyacétone en une seconde molécule de glycéraldéhyde-3-phosphate. Par conséquent, à la fin de cette phase d'amorçage chimique ou de consommation d'énergie, une molécule de glucose est décomposée en deux molécules de glycéraldéhyde-3-phosphate.
La deuxième phase de la glycolyse, la phase productrice d'énergie, crée l'énergie qui est le produit de la glycolyse. La glycéraldéhyde-3-phosphate déshydrogénase convertit chaque glycérodéhyde-3-phosphate à trois carbones produit pendant la phase consommatrice d'énergie en 1,3-bisphosphoglycérate. Cette réaction libère un électron qui est ensuite capté par le NAD+ pour créer une molécule de NADH. Le NADH est une molécule à haute énergie, comme l'ATP, mais contrairement à l'ATP, il n'est pas utilisé comme monnaie énergétique par la cellule. Comme il existe deux molécules de glycéraldéhyde-3-phosphate, deux molécules de NADH sont synthétisées au cours de cette étape. Chaque 1,3-bisphosphoglycérate est ensuite déphosphorylé (c'est-à-dire qu'un phosphate est retiré) par la phosphoglycérate kinase en 3-phosphoglycérate. Chaque phosphate libéré lors de cette réaction peut convertir une molécule d'ADP en une molécule d'ATP à haute énergie, ce qui se traduit par un gain de deux molécules d'ATP.
L'enzyme phosphoglycérate mutase convertit ensuite les molécules de 3-phosphoglycérate en 2-phosphoglycérate. L'enzyme énolase agit ensuite sur les molécules de 2-phosphoglycérate pour les convertir en molécules de phosphoénolpyruvate. La dernière étape de la glycolyse implique la déphosphorylation des deux molécules de phosphoénolpyruvate par la pyruvate kinase pour créer deux molécules de pyruvate et deux molécules d'ATP.
En résumé, une molécule de glucose se décompose en deux molécules de pyruvate et crée deux molécules nettes d'ATP et deux molécules de NADH par glycolyse. Par conséquent, la glycolyse génère de l'énergie pour la cellule et crée des molécules de pyruvate qui peuvent être traitées ultérieurement par le biais du cycle aérobie de Krebs (également appelé cycle de l'acide citrique ou cycle de l'acide tricarboxylique) ; converties en acide lactique ou en alcool (dans la levure) par fermentation ; ou utilisées ultérieurement pour la synthèse de glucose par gluconéogenèse.
Respiration anaérobie
Lorsque l'oxygène est limité ou absent, le pyruvate entre dans une voie anaérobie. Dans ces réactions, le pyruvate peut être transformé en acide lactique. En plus de générer un ATP supplémentaire, cette voie permet de maintenir la concentration de pyruvate à un faible niveau afin que la glycolyse se poursuive, et elle oxyde le NADH en NAD+ nécessaire à la glycolyse. Dans cette réaction, l'acide lactique remplace l'oxygène en tant que dernier accepteur d'électrons. La respiration anaérobie se produit dans la plupart des cellules du corps lorsque l'oxygène est limité ou que les mitochondries sont absentes ou non fonctionnelles. Par exemple, comme les érythrocytes (globules rouges) sont dépourvus de mitochondries, ils doivent produire leur ATP par la respiration anaérobie. Il s'agit d'une voie efficace de production d'ATP pendant de courtes périodes, allant de quelques secondes à quelques minutes. L'acide lactique produit diffuse dans le plasma et est transporté vers le foie, où il est reconverti en pyruvate ou en glucose via le cycle de Cori. De même, lorsqu'une personne fait de l'exercice, les muscles utilisent l'ATP plus rapidement que l'oxygène ne peut leur être fourni. Ils dépendent de la glycolyse et de la production d'acide lactique pour une production rapide d'ATP.
Respiration aérobie
En présence d'oxygène, le pyruvate peut entrer dans le cycle de Krebs où de l'énergie supplémentaire est extraite lorsque des électrons sont transférés du pyruvate vers les récepteurs NAD +, GDP et FAD, le dioxyde de carbone étant un « déchet » (Figure\(\PageIndex{3}\)). Le NADH et le FADH 2 transmettent des électrons à la chaîne de transport d'électrons, qui utilise l'énergie transférée pour produire de l'ATP. En tant qu'étape terminale de la chaîne de transport des électrons, l'oxygène est l'accepteur d'électrons terminal et crée de l'eau à l'intérieur des mitochondries.
Cycle de Krebs/Cycle de l'acide citrique/Cycle de l'acide tricarboxylique
Les molécules de pyruvate générées lors de la glycolyse sont transportées à travers la membrane mitochondriale jusqu'à la matrice mitochondriale interne, où elles sont métabolisées par des enzymes selon une voie appelée cycle de Krebs (Figure\(\PageIndex{4}\)). Le cycle de Krebs est également communément appelé cycle de l'acide citrique ou cycle de l'acide tricarboxylique (TCA). Au cours du cycle de Krebs, des molécules à haute énergie, notamment l'ATP, le NADH et le FADH 2, sont créées. Le NADH et le FADH 2 font ensuite passer des électrons à travers la chaîne de transport d'électrons dans les mitochondries pour générer davantage de molécules d'ATP.


La molécule de pyruvate à trois carbones générée lors de la glycolyse passe du cytoplasme à la matrice mitochondriale, où elle est convertie par l'enzyme pyruvate déshydrogénase en une molécule d'acétylcoenzyme A à deux carbones (acétyl CoA). Cette réaction est une réaction de décarboxylation oxydative. Il convertit le pyruvate à trois carbones en une molécule d'acétyl CoA à deux carbones, libérant du dioxyde de carbone et transférant deux électrons qui se combinent au NAD+ pour former du NADH. L'acétyl CoA entre dans le cycle de Krebs en se combinant à une molécule à quatre carbones, l'oxaloacétate, pour former le citrate, ou acide citrique, une molécule à six carbones, tout en libérant la molécule de coenzyme A.
La molécule de citrate à six carbones est systématiquement convertie en une molécule à cinq carbones puis en une molécule à quatre carbones, se terminant par l'oxaloacétate, début du cycle. En cours de route, chaque molécule de citrate produira un ATP, un FADH 2 et trois NADH. Le FADH 2 et le NADH entreront dans le système de phosphorylation oxydative situé dans la membrane mitochondriale interne. De plus, le cycle de Krebs fournit les matières premières nécessaires à la transformation et à la décomposition des protéines et des graisses.
Pour démarrer le cycle de Krebs, la citrate synthase combine l'acétyl CoA et l'oxaloacétate pour former une molécule de citrate à six carbones ; le CoA est ensuite libéré et peut se combiner avec une autre molécule de pyruvate pour recommencer le cycle. L'enzyme aconitase convertit le citrate en isocitrate. Au cours de deux étapes successives de décarboxylation oxydative, deux molécules de CO 2 et deux molécules de NADH sont produites lorsque l'isocitrate déshydrogénase convertit l'isocitrate en α-cétoglutarate à cinq carbones, qui est ensuite catalysé et converti en succinyl CoA à quatre carbones par l'α-cétoglutarate déshydrogénase. L'enzyme succinyl CoA déshydrogénase convertit ensuite le succinyl CoA en succinate et forme la molécule à haute énergie GTP, qui transfère son énergie à l'ADP pour produire de l'ATP. La succinate déshydrogénase convertit ensuite le succinate en fumarate, formant une molécule de FADH 2. La fumarase convertit ensuite le fumarate en malate, que la malate déshydrogénase reconvertit ensuite en oxaloacétate tout en réduisant le NAD+ en NADH. L'oxaloacétate est alors prêt à se combiner avec l'acétyl CoA suivant pour recommencer le cycle de Krebs (voir Figure\(\PageIndex{4}\)). Pour chaque tour du cycle, trois NADH, un ATP (via GTP) et un FADH 2 sont créés. Chaque carbone du pyruvate est converti en CO 2, qui est libéré en tant que sous-produit de la respiration oxydative (aérobie).
Phosphorylation oxydative et chaîne de transport d'électrons
La chaîne de transport d'électrons (ETC) utilise le NADH et le FADH 2 produits par le cycle de Krebs pour générer de l'ATP. Les électrons du NADH et du FADH 2 sont transférés par des complexes protéiques intégrés dans la membrane mitochondriale interne par une série de réactions enzymatiques. La chaîne de transport d'électrons se compose d'une série de quatre complexes enzymatiques (complexe I — complexe IV) et de deux coenzymes (ubiquinone et cytochrome c), qui agissent comme des porteurs d'électrons et des pompes à protons utilisées pour transférer les ions H+ dans l'espace situé entre les membranes mitochondriales interne et externe (Figure \(\PageIndex{5}\)). L'ETC couple le transfert d'électrons entre un donneur (comme le NADH) et un accepteur d'électrons (comme l'O 2) au transfert de protons (ions H +) à travers la membrane mitochondriale interne, permettant ainsi le processus de phosphorylation oxydative. En présence d'oxygène, l'énergie passe, par étapes, à travers les porteurs d'électrons pour collecter progressivement l'énergie nécessaire pour fixer un phosphate à l'ADP et produire de l'ATP. L'oxygène moléculaire, O 2, joue le rôle d'accepteur d'électrons terminal pour l'ETC. Cela signifie qu'une fois que les électrons ont traversé l'ensemble de l'ETC, ils doivent être transmis à une autre molécule distincte. Ces électrons, O 2 et les ions H + de la matrice, se combinent pour former de nouvelles molécules d'eau. C'est la base de votre besoin de respirer de l'oxygène. Sans oxygène, le flux d'électrons à travers l'ETC cesse.

Les électrons libérés par le NADH et le FADH 2 sont transmis le long de la chaîne par chacun des porteurs, qui sont réduits lorsqu'ils reçoivent l'électron et oxydés lorsqu'ils le transmettent au porteur suivant. Chacune de ces réactions libère une petite quantité d'énergie, qui est utilisée pour pomper les ions H + à travers la membrane interne. L'accumulation de ces protons dans l'espace entre les membranes crée un gradient de protons par rapport à la matrice mitochondriale.
Un incroyable complexe protéique poreux appelé ATP synthase est également intégré à la membrane mitochondriale interne. En fait, il s'agit d'une turbine qui est alimentée par le flux d'ions H + à travers la membrane interne, le long d'un gradient et jusque dans la matrice mitochondriale. Lorsque les ions H + traversent le complexe, l'arbre du complexe tourne. Cette rotation permet à d'autres portions de l'ATP synthase d'encourager l'ADP et le P i à créer de l'ATP. Pour comptabiliser le nombre total d'ATP produit par molécule de glucose par la respiration aérobie, il est important de se rappeler les points suivants :
- Un filet de deux ATP est produit par glycolyse (quatre produits et deux consommés pendant la phase consommatrice d'énergie). Cependant, ces deux ATP sont utilisés pour transporter le NADH produit lors de la glycolyse du cytoplasme vers les mitochondries. Par conséquent, la production nette d'ATP lors de la glycolyse est nulle.
- Dans toutes les phases suivant la glycolyse, le nombre d'ATP, de NADH et de FADH 2 produits doit être multiplié par deux pour refléter la façon dont chaque molécule de glucose produit deux molécules de pyruvate.
- Dans l'ETC, environ trois ATP sont produits pour chaque NADH oxydé. Cependant, seuls deux ATP environ sont produits pour chaque FADH 2 oxydé. Les électrons du FADH 2 produisent moins d'ATP, car ils commencent à un point plus bas de l'ETC (Complexe II) que les électrons du NADH (Complexe I) (voir Figure\(\PageIndex{5}\)).
Par conséquent, pour chaque molécule de glucose qui entre dans la respiration aérobie, un total net de 36 ATP sont produits (Figure\(\PageIndex{6}\)).

Gluconéogenèse
La gluconéogenèse est la synthèse de nouvelles molécules de glucose à partir du pyruvate, du lactate, du glycérol ou des acides aminés alanine ou glutamine. Ce processus se produit principalement dans le foie pendant les périodes de faible taux de glucose, c'est-à-dire dans des conditions de jeûne, de famine et de régimes pauvres en glucides. On peut donc se demander pourquoi le corps créerait quelque chose qu'il vient de dépenser beaucoup d'efforts pour décomposer. Certains organes clés, dont le cerveau, peuvent utiliser uniquement le glucose comme source d'énergie ; il est donc essentiel que le corps maintienne une glycémie minimale. Lorsque la glycémie tombe en dessous de ce point, du nouveau glucose est synthétisé par le foie pour ramener la concentration sanguine à la normale.
La gluconéogenèse n'est pas simplement l'inverse de la glycolyse. Il existe des différences importantes (Figure\(\PageIndex{7}\)). Le pyruvate est une matière première courante pour la gluconéogenèse. Tout d'abord, le pyruvate est converti en oxaloacétate. L'oxaloacétate sert ensuite de substrat à l'enzyme phosphoénolpyruvate carboxykinase (PEPCK), qui transforme l'oxaloacétate en phosphoénolpyruvate (PEP). À partir de cette étape, la gluconéogenèse est presque l'inverse de la glycolyse. La PEP est reconvertie en 2-phosphoglycérate, qui est converti en 3-phosphoglycérate. Ensuite, le 3-phosphoglycérate est converti en 1,3 bisphosphoglycérate puis en glycérodéhyde-3-phosphate. Deux molécules de glycéraldéhyde-3-phosphate se combinent ensuite pour former du fructose-1-6-bisphosphate, qui est converti en fructose 6-phosphate puis en glucose-6-phosphate. Enfin, une série de réactions génère le glucose lui-même. Dans la gluconéogenèse (par rapport à la glycolyse), l'enzyme hexokinase est remplacée par la glucose-6-phosphatase et l'enzyme phosphofructokinase-1 est remplacée par la fructose-1,6-bisphosphatase. Cela aide la cellule à réguler la glycolyse et la gluconéogenèse indépendamment l'une de l'autre.
Comme nous le verrons dans le cadre de la lipolyse, les graisses peuvent être décomposées en glycérol, qui peut être phosphorylé pour former du phosphate de dihydroxyacétone ou du DHAP. Le DHAP peut soit entrer dans la voie glycolytique, soit être utilisé par le foie comme substrat pour la gluconéogenèse.

LE VIEILLISSEMENT ET...
Taux métabolique du corps
Le taux métabolique du corps humain diminue de près de 2 % par décennie après l'âge de 30 ans. Les modifications de la composition corporelle, y compris la réduction de la masse musculaire maigre, sont principalement responsables de cette diminution. La perte de masse musculaire la plus spectaculaire et la baisse du taux métabolique qui en résulte se produisent entre 50 et 70 ans. La perte de masse musculaire équivaut à une diminution de la force, ce qui tend à empêcher les personnes âgées de pratiquer suffisamment d'activité physique. Il en résulte un système de rétroaction positive dans lequel la réduction de l'activité physique entraîne une perte musculaire encore plus importante, réduisant ainsi davantage le métabolisme.
Plusieurs mesures peuvent être prises pour aider à prévenir les déclins généraux du métabolisme et à lutter contre la nature cyclique de ces déclins. Il s'agit notamment de prendre le petit déjeuner, de prendre fréquemment de petits repas, de consommer beaucoup de protéines maigres, de boire de l'eau pour rester hydraté, de faire de l'exercice (y compris la musculation) et de dormir suffisamment Ces mesures peuvent aider à empêcher les niveaux d'énergie de baisser et à réduire l'envie d'augmenter la consommation de calories due à une consommation excessive de collations. Bien que ces stratégies ne garantissent pas le maintien du métabolisme, elles aident à prévenir la perte musculaire et peuvent augmenter les niveaux d'énergie. Certains experts suggèrent également d'éviter le sucre, qui peut entraîner un stockage excessif des graisses. Les aliments épicés et le thé vert peuvent également être bénéfiques. Comme le stress active la libération de cortisol et que le cortisol ralentit le métabolisme, éviter le stress, ou du moins pratiquer des techniques de relaxation, peut également aider.
Révision du chapitre
Les enzymes métaboliques catalysent les réactions cataboliques qui dégradent les glucides contenus dans les aliments. L'énergie libérée est utilisée pour alimenter les cellules et les systèmes qui composent votre corps. L'énergie excédentaire ou non utilisée est stockée sous forme de graisse ou de glycogène pour une utilisation ultérieure. Le métabolisme des glucides commence dans la bouche, où l'enzyme amylase salivaire commence à décomposer les sucres complexes en monosaccharides. Ceux-ci peuvent ensuite être transportés à travers la membrane intestinale dans la circulation sanguine, puis vers les tissus corporels. Dans les cellules, le glucose, un sucre à six carbones, est transformé par une séquence de réactions en sucres plus petits, et l'énergie stockée à l'intérieur de la molécule est libérée. La première étape du catabolisme des glucides est la glycolyse, qui produit du pyruvate, du NADH et de l'ATP. Dans des conditions anaérobies, le pyruvate peut être converti en lactate pour que la glycolyse continue de fonctionner. Dans des conditions aérobies, le pyruvate entre dans le cycle de Krebs, également appelé cycle de l'acide citrique ou cycle de l'acide tricarboxylique. Outre l'ATP, le cycle de Krebs produit des molécules de FADH 2 et de NADH à haute énergie, qui fournissent des électrons au processus de phosphorylation oxydative qui génère davantage de molécules d'ATP à haute énergie. Pour chaque molécule de glucose traitée par glycolyse, un réseau de 36 ATP peut être créé par la respiration aérobie.
Dans des conditions anaérobies, la production d'ATP est limitée à celle générée par la glycolyse. Alors qu'un total de quatre ATP sont produits par glycolyse, deux sont nécessaires pour commencer la glycolyse, ce qui donne un rendement net de deux molécules d'ATP.
Dans des conditions de faible teneur en glucose, telles que le jeûne, la famine ou les régimes pauvres en glucides, le glucose peut être synthétisé à partir de lactate, de pyruvate, de glycérol, d'alanine ou de glutamate. Ce processus, appelé gluconéogenèse, est presque l'inverse de la glycolyse et sert à créer des molécules de glucose pour les organes dépendant du glucose, tels que le cerveau, lorsque le taux de glucose tombe en dessous de la normale.
Questions de révision
Q. La glycolyse entraîne la production de deux molécules ________ à partir d'une seule molécule de glucose. En l'absence de ________, le produit final de la glycolyse est ________.
A. acétyl CoA, pyruvate, lactate
B. ATP, carbone, pyruvate
C. pyruvate, oxygène, lactate
D. pyruvate, carbone, acétyl CoA
Réponse : C
Q. Le cycle de Krebs convertit ________ en un cycle de réactions. Au cours du processus, de l'ATP, du ________ et du ________ sont produits.
A. acétyl CoA ; FAD, NAD
B. acétyl CoA ; FADH 2 ; NADH
C. pyruvate ; NAD ; ADH 2
D. pyruvate ; oxygène ; oxaloacétate
Réponse : B
Q. Quelle voie produit le plus de molécules d'ATP ?
A. Fermentation de l'acide lactique
B. le cycle de Krebs
C. la chaîne de transport des électrons
D. glycolyse
Réponse : C
Q. La respiration cellulaire aérobie entraîne la production de ces deux produits.
A. NADH et FADH 2
B. ATP et pyruvate
C. ATP et glucose
D. ATP et H 2 O
Réponse : D
Q. Lorsque le NAD+ devient du NADH, la coenzyme est ________.
A. réduit
B. oxydé
C. métabolisé
D. hydrolysé
Réponse : A
Questions sur la pensée critique
Q. Expliquez comment le glucose est métabolisé pour produire de l'ATP.
R. Le glucose est oxydé pendant la glycolyse, créant du pyruvate, qui est traité selon le cycle de Krebs pour produire du NADH, du FADH 2, de l'ATP et du CO 2. Le FADH 2 et le NADH produisent de l'ATP.
Q. L'insuline est libérée lors de l'ingestion d'aliments et stimule l'absorption du glucose dans la cellule. Discutez du mécanisme utilisé par les cellules pour créer un gradient de concentration afin d'assurer l'absorption continue du glucose par le sang.
R. À l'entrée dans la cellule, l'hexokinase ou la glucokinase phosphoryle le glucose, le convertissant en glucose-6-phosphate. Sous cette forme, le glucose-6-phosphate est piégé dans la cellule. Comme tout le glucose a été phosphorylé, de nouvelles molécules de glucose peuvent être transportées dans la cellule en fonction de son gradient de concentration.
Lexique
- acétylcoenzyme A (acétyl CoA)
- molécule de départ du cycle de Krebs
- ATP synthase
- complexe de pores protéiques qui crée de l'ATP
- respiration cellulaire
- production d'ATP par oxydation du glucose par glycolyse, cycle de Krebs et phosphorylation oxydative
- cycle d'acide citrique
- également appelé cycle de Krebs ou cycle de l'acide tricarboxylique ; convertit le pyruvate en CO 2 et en molécules de FADH 2, de NADH et d'ATP à haute énergie
- chaîne de transport d'électrons (ETC)
- Voie de production d'ATP dans laquelle les électrons sont soumis à une série de réactions d'oxydoréduction qui forment de l'eau et produisent un gradient de protons
- phase consommatrice d'énergie
- première phase de glycolyse, dans laquelle deux molécules d'ATP sont nécessaires pour démarrer la réaction
- phase de production d'énergie
- deuxième phase de glycolyse, au cours de laquelle de l'énergie est produite
- glucokinase
- enzyme cellulaire, présente dans le foie, qui convertit le glucose en glucose-6-phosphate lors de son absorption dans la cellule
- gluconéogenèse
- processus de synthèse du glucose à partir de pyruvate ou d'autres molécules
- glucose-6-phosphate
- glucose phosphorylé produit lors de la première étape de la glycolyse
- glycolyse
- série de réactions métaboliques qui décomposent le glucose en pyruvate et produisent de l'ATP
- hexokinase
- enzyme cellulaire, présente dans la plupart des tissus, qui convertit le glucose en glucose-6-phosphate lors de son absorption dans la cellule
- Cycle de Krebs
- également appelé cycle de l'acide citrique ou cycle de l'acide tricarboxylique, convertit le pyruvate en CO 2 et en molécules de FADH 2, de NADH et d'ATP à haute énergie
- monosaccharide
- la plus petite molécule de sucre monomère
- phosphorylation oxydative
- procédé qui convertit le NADH et le FADH 2 à haute énergie en ATP
- polysaccharides
- glucides complexes composés de nombreux monosaccharides
- pyruvate
- produit final à trois carbones de la glycolyse et matière de départ convertie en acétyl CoA entrant dans le cycle de Krebs
- amylase salivaire
- enzyme digestive qui se trouve dans la salive et qui commence la digestion des glucides dans la bouche
- accepteur d'électrons terminal
- l'oxygène, récepteur de l'hydrogène libre à la fin de la chaîne de transport des électrons
Contributeurs et attributions
- tricarboxylic acid cycle (TCA)
- also called the Krebs cycle or the citric acid cycle; converts pyruvate into CO2 and high-energy FADH2, NADH, and ATP molecules
- Template:ContribOpenStaxAP