
22.5 : Transport de gaz
- Page ID
- 195283

Objectifs d'apprentissage
- Décrire les principes du transport de l'oxygène
- Décrire la structure de l'hémoglobine
- Comparer et contraster l'hémoglobine foetale et adulte
- Décrire les principes du transport du dioxyde de carbone
L'autre activité majeure des poumons est le processus de respiration, le processus d'échange de gaz. La fonction de la respiration est de fournir de l'oxygène aux cellules du corps pendant la respiration cellulaire et d'éliminer du corps le dioxyde de carbone, un déchet de la respiration cellulaire. Pour que l'échange d'oxygène et de dioxyde de carbone se produise, les deux gaz doivent être transportés entre les sites de respiration externe et interne. Bien que le dioxyde de carbone soit plus soluble que l'oxygène dans le sang, les deux gaz nécessitent un système de transport spécialisé pour que la majorité des molécules de gaz soient déplacées entre les poumons et les autres tissus.
Transport de l'oxygène dans le sang
Même si l'oxygène est transporté par le sang, vous vous souvenez peut-être que l'oxygène n'est pas très soluble dans les liquides. Une petite quantité d'oxygène se dissout dans le sang et est transportée dans le sang, mais elle ne représente qu'environ 1,5 % de la quantité totale. La majorité des molécules d'oxygène sont transportées des poumons vers les tissus de l'organisme par un système de transport spécialisé qui repose sur les érythrocytes, les globules rouges. Les érythrocytes contiennent une métalloprotéine, l'hémoglobine, qui sert à lier les molécules d'oxygène aux érythrocytes (Figure\(\PageIndex{1}\)). L'hème est la partie de l'hémoglobine qui contient du fer et c'est l'hème qui lie l'oxygène. Un érythrocyte contient quatre ions fer et, de ce fait, chaque érythrocyte est capable de transporter jusqu'à quatre molécules d'oxygène. Lorsque l'oxygène se diffuse à travers la membrane respiratoire, de l'alvéole au capillaire, il se diffuse également dans les globules rouges et est lié par l'hémoglobine. La réaction chimique réversible suivante décrit la production du produit final, l'oxyhémoglobine (Hb—O 2), qui se forme lorsque l'oxygène se lie à l'hémoglobine. L'oxyhémoglobine est une molécule de couleur rouge vif qui contribue à la couleur rouge vif du sang oxygéné.
\[Hb + O_{2(g)} \rightleftharpoons Hb-O_2\]
Dans cette formule, Hb représente l'hémoglobine réduite, c'est-à-dire l'hémoglobine à laquelle l'oxygène n'est pas lié. Plusieurs facteurs entrent en jeu dans la rapidité avec laquelle l'hème se lie à l'oxygène et s'en dissocie, qui seront abordés dans les sections suivantes.

Fonction de l'hémoglobine
L'hémoglobine est composée de sous-unités, une structure protéique appelée structure quaternaire. Chacune des quatre sous-unités qui composent l'hémoglobine est organisée en anneau, avec un atome de fer lié de manière covalente à l'hème au centre de chaque sous-unité. La liaison de la première molécule d'oxygène provoque une modification conformationnelle de l'hémoglobine qui permet à la seconde molécule d'oxygène de se lier plus facilement. Lorsque chaque molécule d'oxygène est liée, cela facilite davantage la liaison de la molécule suivante, jusqu'à ce que les quatre sites hémiques soient occupés par de l'oxygène. L'inverse se produit également : une fois que la première molécule d'oxygène s'est dissociée et est « déposée » dans les tissus, la molécule d'oxygène suivante se dissocie plus facilement. Lorsque les quatre sites hémiques sont occupés, on dit que l'hémoglobine est saturée. Lorsqu'un à trois sites hémiques sont occupés, on dit que l'hémoglobine est partiellement saturée. Par conséquent, si l'on considère le sang dans son ensemble, le pourcentage d'unités d'hème disponibles qui sont liées à l'oxygène à un moment donné est appelé saturation en hémoglobine. Une saturation en hémoglobine de 100 pour cent signifie que chaque unité d'hème de tous les érythrocytes du corps est liée à l'oxygène. Chez une personne en bonne santé dont le taux d'hémoglobine est normal, la saturation en hémoglobine varie généralement de 95 % à 99 %.
Dissociation de l'oxygène de l'
La pression partielle est un aspect important de la liaison de l'oxygène à l'hème et de sa dissociation de celui-ci. Une courbe de dissociation oxygène—hémoglobine est un graphique qui décrit la relation entre la pression partielle et la liaison de l'oxygène à l'hème et sa dissociation ultérieure de l'hème (Figure\(\PageIndex{2}\)). N'oubliez pas que les gaz se déplacent d'une zone de pression partielle plus élevée vers une zone de pression partielle plus faible. De plus, l'affinité d'une molécule d'oxygène pour l'hème augmente à mesure que de plus en plus de molécules d'oxygène sont liées. Par conséquent, dans la courbe de saturation oxygène—hémoglobine, lorsque la pression partielle d'oxygène augmente, un nombre proportionnellement plus élevé de molécules d'oxygène sont liées par l'hème. Il n'est pas surprenant que la courbe de saturation/dissociation oxygène—hémoglobine montre également que plus la pression partielle d'oxygène est faible, moins de molécules d'oxygène sont liées à l'hème. Par conséquent, la pression partielle d'oxygène joue un rôle majeur dans la détermination du degré de liaison de l'oxygène à l'hème au niveau de la membrane respiratoire, ainsi que du degré de dissociation de l'oxygène de l'hème au niveau des tissus corporels.
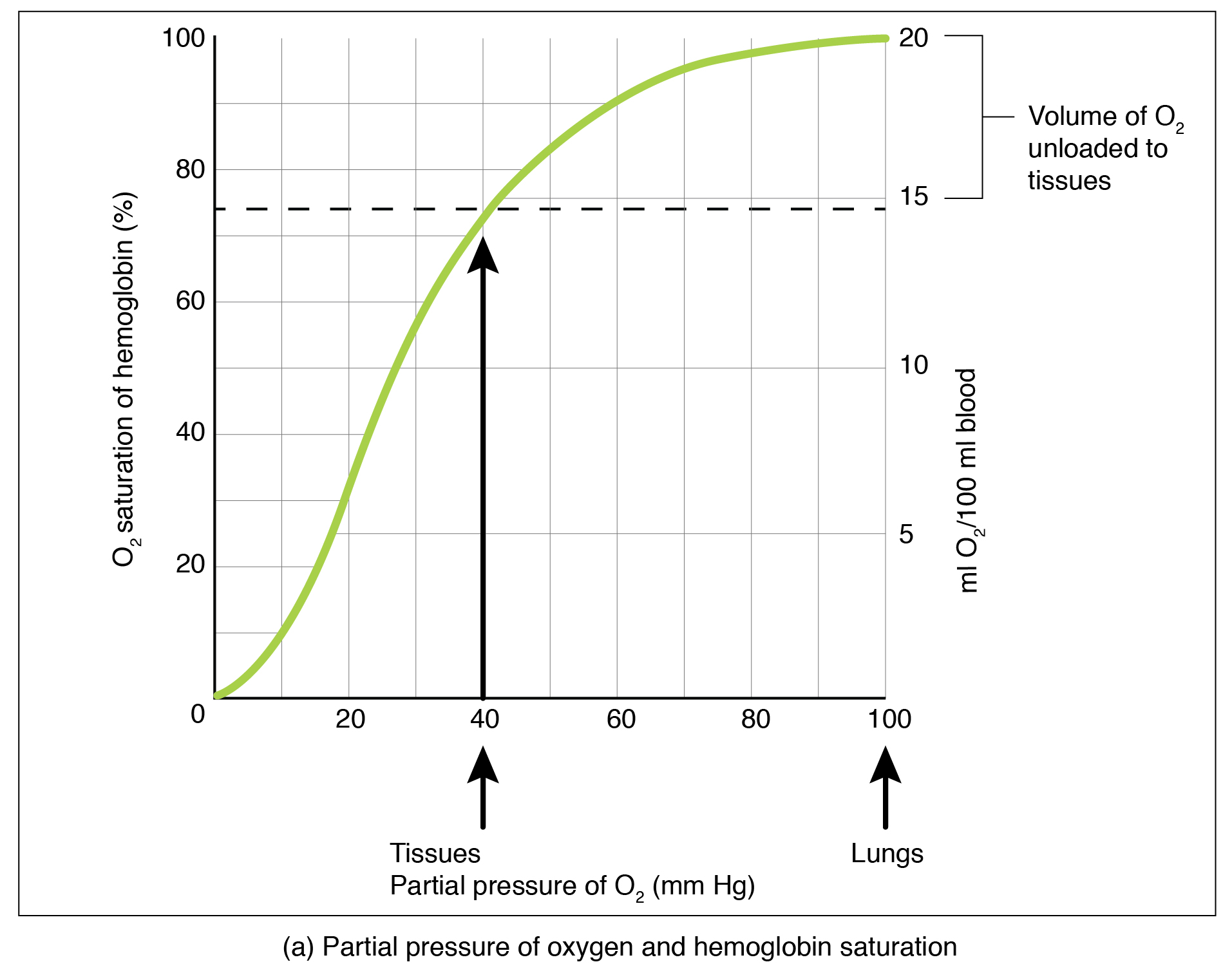
Les mécanismes à l'origine de la courbe de saturation/dissociation oxygène-hémoglobine servent également de mécanismes de contrôle automatique qui régulent la quantité d'oxygène délivrée aux différents tissus du corps. Ceci est important parce que certains tissus ont un taux métabolique plus élevé que d'autres. Les tissus très actifs, tels que les muscles, utilisent rapidement l'oxygène pour produire de l'ATP, abaissant ainsi la pression partielle d'oxygène dans les tissus à environ 20 mm Hg. La pression partielle de l'oxygène à l'intérieur des capillaires est d'environ 100 mm Hg, de sorte que la différence entre les deux devient assez élevée, environ 80 mm Hg. En conséquence, un plus grand nombre de molécules d'oxygène se dissocient de l'hémoglobine et pénètrent dans les tissus. L'inverse est vrai pour les tissus, tels que le tissu adipeux (graisse corporelle), dont le taux métabolique est plus faible. Comme ces cellules utilisent moins d'oxygène, la pression partielle d'oxygène dans ces tissus reste relativement élevée, ce qui se traduit par une diminution du nombre de molécules d'oxygène qui se dissocient de l'hémoglobine et pénètrent dans le liquide interstitiel tissulaire. Bien que l'on dise que le sang veineux est désoxygéné, une partie de l'oxygène est toujours liée à l'hémoglobine dans ses globules rouges. Cela fournit une réserve d'oxygène qui peut être utilisée lorsque les tissus demandent soudainement plus d'oxygène.
Des facteurs autres que la pression partielle influent également sur la courbe de saturation/dissociation oxygène-hémoglobine. Par exemple, une température plus élevée favorise la dissociation plus rapide de l'hémoglobine et de l'oxygène, alors qu'une température plus basse inhibe la dissociation (voir Figure\(\PageIndex{2}\) au milieu). Cependant, le corps humain régule étroitement la température, de sorte que ce facteur peut ne pas affecter les échanges gazeux dans tout le corps. Les tissus très actifs font exception à cette règle, car ils peuvent libérer une plus grande quantité d'énergie que celle dégagée sous forme de chaleur. En conséquence, l'oxygène se dissocie facilement de l'hémoglobine, un mécanisme qui contribue à fournir plus d'oxygène aux tissus actifs.
Certaines hormones, telles que les androgènes, l'épinéphrine, les hormones thyroïdiennes et l'hormone de croissance, peuvent affecter la courbe de saturation/dissociation oxygène-hémoglobine en stimulant la production d'un composé appelé 2,3-bisphosphoglycérate (BPG) par les érythrocytes. Le BPG est un sous-produit de la glycolyse. Comme les érythrocytes ne contiennent pas de mitochondries, la glycolyse est la seule méthode par laquelle ces cellules produisent de l'ATP. Le BPG favorise la dissociation de l'oxygène de l'hémoglobine. Par conséquent, plus la concentration de BPG est élevée, plus l'oxygène se dissocie facilement de l'hémoglobine, malgré sa pression partielle.
Le pH du sang est un autre facteur qui influence la courbe de saturation/dissociation oxygène-hémoglobine (voir Figure\(\PageIndex{2}\)). L'effet Bohr est un phénomène qui résulte de la relation entre le pH et l'affinité de l'oxygène pour l'hémoglobine : un pH plus bas et plus acide favorise la dissociation de l'oxygène de l'hémoglobine. En revanche, un pH plus élevé ou plus basique inhibe la dissociation de l'oxygène de l'hémoglobine. Plus la quantité de dioxyde de carbone dans le sang est importante, plus il faut convertir de molécules, ce qui génère des ions hydrogène et abaisse ainsi le pH sanguin. De plus, le pH sanguin peut devenir plus acide lorsque certains sous-produits du métabolisme cellulaire, tels que l'acide lactique, l'acide carbonique et le dioxyde de carbone, sont libérés dans la circulation sanguine.
Hémoglobine du foetus
Le foetus a sa propre circulation avec ses propres érythrocytes ; cependant, il dépend de la mère pour l'oxygène. Le sang est fourni au foetus par le cordon ombilical, qui est relié au placenta et séparé du sang maternel par le chorion. Le mécanisme d'échange gazeux au niveau du chorion est similaire à celui de l'échange gazeux au niveau de la membrane respiratoire. Cependant, la pression partielle d'oxygène est plus faible dans le sang maternel dans le placenta, à environ 35 à 50 mm Hg, que dans le sang artériel maternel. La différence de pression partielle entre le sang maternel et le sang foetal n'est pas grande, car la pression partielle d'oxygène dans le sang foetal au niveau du placenta est d'environ 20 mm Hg. Par conséquent, il n'y a pas autant de diffusion d'oxygène dans l'approvisionnement en sang du fœtus. L'hémoglobine du fœtus surmonte ce problème en ayant une plus grande affinité pour l'oxygène que l'hémoglobine maternelle (Figure\(\PageIndex{3}\)). L'hémoglobine foetale et l'hémoglobine adulte ont quatre sous-unités, mais deux des sous-unités de l'hémoglobine foetale ont une structure différente qui fait que l'hémoglobine foetale a une plus grande affinité pour l'oxygène que l'hémoglobine adulte.

Transport du dioxyde de carbone dans le sang
Le dioxyde de carbone est transporté par trois mécanismes principaux. Le premier mécanisme de transport du dioxyde de carbone est le plasma sanguin, car certaines molécules de dioxyde de carbone se dissolvent dans le sang. Le deuxième mécanisme est le transport sous forme de bicarbonate (HCO 3 —), qui se dissout également dans le plasma. Le troisième mécanisme de transport du dioxyde de carbone est similaire au transport de l'oxygène par les érythrocytes (Figure\(\PageIndex{4}\)).
Dioxyde de carbone dissous
Bien que le dioxyde de carbone ne soit pas considéré comme très soluble dans le sang, une petite fraction (environ 7 à 10 %) du dioxyde de carbone diffusé dans le sang à partir des tissus se dissout dans le plasma. Le dioxyde de carbone dissous se déplace ensuite dans la circulation sanguine et, lorsque le sang atteint les capillaires pulmonaires, le dioxyde de carbone dissous se diffuse à travers la membrane respiratoire jusqu'aux alvéoles, où il est ensuite expiré lors de la ventilation pulmonaire.
tampon au bicarbonate
Une grande partie (environ 70 %) des molécules de dioxyde de carbone qui se diffusent dans le sang est transportée vers les poumons sous forme de bicarbonate. La majeure partie du bicarbonate est produite dans les érythrocytes après la diffusion du dioxyde de carbone dans les capillaires, puis dans les globules rouges. L'anhydrase carbonique (CA) amène le dioxyde de carbone et l'eau à former de l'acide carbonique (H 2 CO 3), qui se dissocie en deux ions : le bicarbonate (HCO 3 —) et l'hydrogène (H +). La formule suivante décrit cette réaction :
Le bicarbonate a tendance à s'accumuler dans les érythrocytes, de sorte qu'il y a une plus grande concentration de bicarbonate dans les érythrocytes que dans le plasma sanguin environnant. Par conséquent, une partie du bicarbonate quittera les érythrocytes et se déplacera le long de son gradient de concentration vers le plasma en échange d'ions chlorure (Cl —). Ce phénomène, appelé déplacement du chlorure, se produit parce qu'en échangeant un ion négatif contre un autre ion négatif, ni la charge électrique des érythrocytes ni celle du sang ne sont modifiées.
Au niveau des capillaires pulmonaires, la réaction chimique qui a produit le bicarbonate (voir ci-dessus) est inversée, et le dioxyde de carbone et l'eau sont les produits. Une grande partie du bicarbonate présent dans le plasma rentre dans les érythrocytes en échange d'ions chlorure. Les ions hydrogène et les ions bicarbonate se rejoignent pour former de l'acide carbonique, qui est converti en dioxyde de carbone et en eau par l'anhydrase carbonique. Le dioxyde de carbone sort des érythrocytes et pénètre dans le plasma, où il peut ensuite se diffuser à travers la membrane respiratoire et atteindre les alvéoles pour être expiré lors de la ventilation pulmonaire.
Carbaminohémoglobine
Environ 20 pour cent du dioxyde de carbone est lié à l'hémoglobine et est transporté vers les poumons. Le dioxyde de carbone ne se lie pas au fer comme le fait l'oxygène ; il lie plutôt des groupes d'acides aminés sur les parties globines de l'hémoglobine pour former de la carbaminohémoglobine, qui se forme lorsque l'hémoglobine et le dioxyde de carbone se lient. Lorsque l'hémoglobine ne transporte pas l'oxygène, elle a tendance à prendre une teinte bleu-violet, ce qui donne la couleur marron foncé typique du sang désoxygéné. La formule suivante décrit cette réaction réversible :
\[CO_{2(g)}+Hb \rightleftharpoons HbCO_2\]
Tout comme le transport de l'oxygène par l'hème, la liaison et la dissociation du dioxyde de carbone vers et depuis l'hémoglobine dépendent de la pression partielle du dioxyde de carbone. Comme le dioxyde de carbone est libéré par les poumons, le sang qui quitte les poumons et atteint les tissus corporels a une pression partielle de dioxyde de carbone inférieure à celle que l'on trouve dans les tissus. En conséquence, le dioxyde de carbone quitte les tissus en raison de sa pression partielle plus élevée, pénètre dans le sang, puis se déplace dans les globules rouges et se lie à l'hémoglobine. En revanche, dans les capillaires pulmonaires, la pression partielle de dioxyde de carbone est élevée par rapport à l'intérieur des alvéoles. Par conséquent, le dioxyde de carbone se dissocie facilement de l'hémoglobine et se diffuse dans l'air à travers la membrane respiratoire.
Outre la pression partielle du dioxyde de carbone, la saturation en oxygène de l'hémoglobine et la pression partielle d'oxygène dans le sang influencent également l'affinité de l'hémoglobine pour le dioxyde de carbone. L'effet Haldane est un phénomène qui résulte de la relation entre la pression partielle d'oxygène et l'affinité de l'hémoglobine pour le dioxyde de carbone. L'hémoglobine saturée en oxygène ne lie pas facilement le dioxyde de carbone. Cependant, lorsque l'oxygène n'est pas lié à l'hème et que la pression partielle de l'oxygène est faible, l'hémoglobine se lie facilement au dioxyde de carbone.

Regardez cette vidéo pour voir le transport de l'oxygène des poumons vers les tissus. Pourquoi le sang oxygéné est-il rouge vif alors que le sang désoxygéné a tendance à être de couleur violette ?
Révision du chapitre
L'oxygène est principalement transporté dans le sang par les érythrocytes. Ces cellules contiennent une métalloprotéine appelée hémoglobine, qui est composée de quatre sous-unités ayant une structure en anneau. Chaque sous-unité contient un atome de fer lié à une molécule d'hème. L'hème lie l'oxygène de sorte que chaque molécule d'hémoglobine peut lier jusqu'à quatre molécules d'oxygène. Lorsque toutes les unités hémiques du sang sont liées à l'oxygène, l'hémoglobine est considérée comme saturée. L'hémoglobine est partiellement saturée lorsque seules certaines unités hémiques sont liées à l'oxygène. Une courbe de saturation/dissociation oxygène-hémoglobine est une méthode courante pour décrire la relation entre la facilité avec laquelle l'oxygène se lie à l'hémoglobine ou s'en dissocie en fonction de la pression partielle de l'oxygène. Plus la pression partielle d'oxygène augmente, plus l'hémoglobine se lie facilement à l'oxygène. En même temps, une fois qu'une molécule d'oxygène est liée à l'hémoglobine, d'autres molécules d'oxygène se lient plus facilement à l'hémoglobine. D'autres facteurs tels que la température, le pH, la pression partielle du dioxyde de carbone et la concentration de 2,3-bisphosphoglycérate peuvent également améliorer ou inhiber la liaison de l'hémoglobine et de l'oxygène. L'hémoglobine foetale a une structure différente de l'hémoglobine adulte, ce qui fait que l'hémoglobine foetale a une plus grande affinité pour l'oxygène que l'hémoglobine adulte.
Le dioxyde de carbone est transporté dans le sang par trois mécanismes différents : sous forme de dioxyde de carbone dissous, de bicarbonate ou de carbaminohémoglobine. Il reste une petite portion de dioxyde de carbone. La plus grande quantité de dioxyde de carbone transporté se trouve sous forme de bicarbonate, formé dans les érythrocytes. Pour cette conversion, le dioxyde de carbone est combiné à de l'eau à l'aide d'une enzyme appelée anhydrase carbonique. Cette combinaison forme de l'acide carbonique qui se dissocie spontanément en ions bicarbonate et hydrogène. Au fur et à mesure que le bicarbonate s'accumule dans les érythrocytes, il traverse la membrane et pénètre dans le plasma en échange d'ions chlorure par un mécanisme appelé déplacement du chlorure. Au niveau des capillaires pulmonaires, le bicarbonate rentre dans les érythrocytes en échange d'ions chlorure, et la réaction avec l'anhydrase carbonique est inversée, recréant du dioxyde de carbone et de l'eau. Le dioxyde de carbone se diffuse ensuite hors des érythrocytes et traverse la membrane respiratoire dans l'air. Une quantité intermédiaire de dioxyde de carbone se lie directement à l'hémoglobine pour former de la carbaminohémoglobine. Les pressions partielles du dioxyde de carbone et de l'oxygène, ainsi que la saturation en oxygène de l'hémoglobine, influencent la facilité avec laquelle l'hémoglobine lie le dioxyde de carbone. Moins l'hémoglobine est saturée et plus la pression partielle d'oxygène dans le sang est faible, plus l'hémoglobine se lie facilement au dioxyde de carbone. C'est un exemple de l'effet Haldane.
Questions sur les liens interactifs
Regardez cette vidéo pour voir le transport de l'oxygène des poumons vers les tissus. Pourquoi le sang oxygéné est-il rouge vif alors que le sang désoxygéné a tendance à être de couleur violette ?
Réponse : Lorsque l'oxygène se lie à la molécule d'hémoglobine, de l'oxyhémoglobine est créée, qui a une couleur rouge. L'hémoglobine qui n'est pas liée à l'oxygène a tendance à être de couleur bleu-violet. Le sang oxygéné circulant dans les artères systémiques contient de grandes quantités d'oxyhémoglobine. Lorsque le sang traverse les tissus, une grande partie de l'oxygène est libérée dans les capillaires systémiques. Le sang désoxygéné qui revient par les veines systémiques contient donc des quantités beaucoup plus faibles d'oxyhémoglobine. Plus il y a d'oxyhémoglobine dans le sang, plus le liquide sera rouge. En conséquence, le sang oxygéné sera beaucoup plus rouge que le sang désoxygéné.
Questions de révision
Q. L'oxyhémoglobine se forme par réaction chimique entre laquelle des substances suivantes ?
A. hémoglobine et dioxyde de carbone
B. anhydrase carbonique et dioxyde de carbone
C. hémoglobine et oxygène
D. anhydrase carbonique et oxygène
Réponse : C
Q. Lequel des facteurs suivants joue un rôle dans la courbe de saturation/dissociation oxygène-hémoglobine ?
A. température
B. pH
C. BPG
D. tout ce qui précède
Réponse : D
Q. Lequel des événements suivants se produit lors du changement de chlorure ?
R. Le chlorure est retiré de l'érythrocyte.
B. Le chlorure est remplacé par du bicarbonate.
C. Le bicarbonate est retiré de l'érythrocyte.
D. Le bicarbonate est éliminé du sang.
Réponse : B
Q. Une faible pression partielle d'oxygène favorise la liaison de l'hémoglobine au dioxyde de carbone. Voici un exemple de ________.
A. Effet Haldane
B. Effet Bohr
C. La loi de Dalton
D. La loi de Henry
Réponse : A
Questions sur la pensée critique
Q. Comparez et comparez l'hémoglobine adulte et l'hémoglobine foetale.
R. L'hémoglobine adulte et foetale transporte l'oxygène via des molécules de fer. Cependant, l'hémoglobine foetale a une affinité pour l'oxygène environ 20 fois plus grande que l'hémoglobine adulte. Cela est dû à une différence de structure ; l'hémoglobine foetale comporte deux sous-unités qui ont une structure légèrement différente de celle des sous-unités de l'hémoglobine adulte.
Q. Décrivez la relation entre la pression partielle de l'oxygène et la liaison de l'oxygène à l'hémoglobine.
A. La relation entre la pression partielle d'oxygène et la liaison de l'hémoglobine à l'oxygène est décrite par la courbe de saturation/dissociation oxygène—hémoglobine. À mesure que la pression partielle d'oxygène augmente, le nombre de molécules d'oxygène liées à l'hémoglobine augmente, augmentant ainsi la saturation de l'hémoglobine.
Q. Décrivez trois façons dont le dioxyde de carbone peut être transporté.
R. Le dioxyde de carbone peut être transporté par trois mécanismes : dissous dans le plasma, sous forme de bicarbonate ou de carbaminohémoglobine. Dissoutes dans le plasma, les molécules de dioxyde de carbone se diffusent simplement dans le sang à partir des tissus. Le bicarbonate est créé par une réaction chimique qui se produit principalement dans les érythrocytes, réunissant le dioxyde de carbone et l'eau par l'anhydrase carbonique, produisant de l'acide carbonique, qui se décompose en ions bicarbonate et hydrogène. La carbaminohémoglobine est la forme liée de l'hémoglobine et du dioxyde de carbone.
Lexique
- Effet Bohr
- relation entre le pH sanguin et la dissociation de l'oxygène de l'hémoglobine
- carbaminohémoglobine
- forme liée de l'hémoglobine et du dioxyde de carbone
- anhydrase carbonique (CA)
- enzyme qui catalyse la réaction qui provoque la formation d'acide carbonique par le dioxyde de carbone et l'eau
- déplacement des chlorures
- diffusion facilitée qui échange du bicarbonate (HCO 3 —) avec des ions chlorure (Cl —)
- Effet Haldane
- relation entre la pression partielle d'oxygène et l'affinité de l'hémoglobine pour le dioxyde de carbone
- oxyhémoglobine
- Forme liée à l'hémoglobine et à l'oxygène (Hb—O 2)
- courbe de dissociation oxygène-hémoglobine
- graphique décrivant la relation entre la pression partielle et la liaison et la dissociation de l'oxygène vers et depuis l'hème