
11.1:发现人口如何变化
- Page ID
- 199108

自然选择进化理论描述了物种随时间推移而变化的机制。 这种物种变化早在达尔文之前就已经提出过建议和辩论。 认为物种是静止不变的观点以柏拉图的著作为基础,但也有古希腊人表达了进化思想。
在十八世纪,博物学家乔治·路易·勒克莱尔、伯爵德布冯甚至查尔斯·达尔文的祖父伊拉斯姆斯·达尔文都重新引入了关于动物进化的观念。 在这段时间里,人们也承认存在灭绝的物种。 同时,苏格兰博物学家詹姆斯·赫顿(James Hutton)提出,地质变化是通过过程中的微小变化(在很长一段时间内)的积累逐渐发生的,就像今天发生的那样。 这与占主导地位的观点形成鲜明对比,即地球的地质学是相对较短的过去发生的灾难性事件的结果。 赫顿的观点后来在十九世纪被地质学家查尔斯·莱尔所普及。 莱尔成为了达尔文的朋友,他的想法对达尔文的思想产生了很大的影响。 莱尔认为,地球年龄的延长为物种的逐渐变化提供了更多的时间,而这个过程为物种的逐渐变化提供了一个类比。
十九世纪初,让-巴蒂斯特·拉马克出版了一本书,详细介绍了进化变化的机制,这种机制现在被称为后天特征的继承。 在拉马克的理论中,由环境引起的个体改变,或者在其生命周期中使用或废弃结构,可以由其后代继承,从而带来物种的变化。 尽管拉马克所描述的这种进化变革机制被抹黑了,但拉马克的思想对进化思想产生了重要影响。 站在巴黎植物园门口的拉马克雕像上的铭文将他描述为 “进化论的创始人”。
查尔斯·达尔文与自然选择
进化的实际机制是由两位博物学家查尔斯·达尔文和阿尔弗雷德·罗素·华莱士在十九世纪中叶独立构思和描述的。 重要的是,每个人都花时间在热带探险中探索自然世界。 从 1831 年到 1836 年,达尔文乘坐 H.M.S. Beagle 环游世界,访问了南美、澳大利亚和非洲的南端。 华莱士于 1848 年至 1852 年前往巴西在亚马逊雨林采集昆虫,1854 年至 1862 年前往马来群岛。 达尔文的旅程,就像华莱士后来在马来群岛的旅程一样,包括在几个岛屿链上停留,最后一次是加拉帕戈斯群岛(厄瓜多尔以西)。 在这些岛屿上,达尔文观察到不同岛屿上存在明显相似但有明显差异的生物种类。 例如,居住在加拉帕戈斯群岛的地下雀由几种物种组成,每个物种都有独特的喙形状(图\(\PageIndex{1}\))。 他观察到这些雀科与南美大陆的另一种雀科物种非常相似,加拉帕戈斯群岛的物种群形成了一系列分级的喙大小和形状,最相似的鸟之间的差异很小。 达尔文认为,岛屿物种可能是从一个原始大陆物种改造而来的所有物种。 1860年,他写道:“看到一小群密切相关的鸟类结构的渐变和多样性,人们可能真的想到,从这个群岛最初缺少鸟类的情况来看,有一个物种被捕获和改造用于不同的目的。” 1
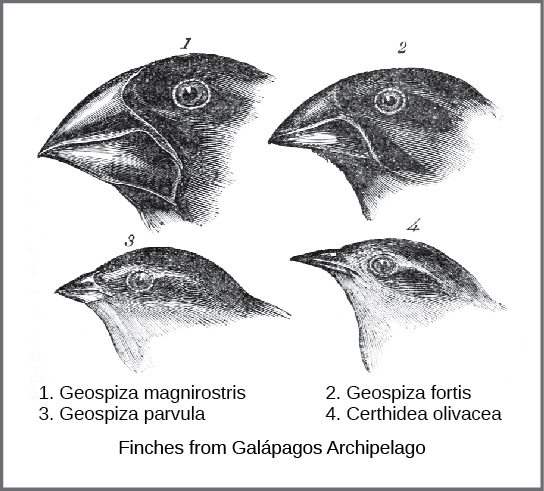
华莱士和达尔文都在其他生物中观察到类似的模式,并独立构思了一种机制来解释这种变化如何以及为什么会发生。 达尔文称这种机制为自然选择。 达尔文认为,自然选择是自然界中运作的三项原则的必然结果。 首先,生物的特征是遗传的,或者是从父母传给后代的。 其次,生出的后代多于存活的能力;换句话说,生存和繁殖的资源有限。 所有生物的繁殖能力都超过了支持其数量的可用资源。 因此,每一代人都在争夺这些资源。 达尔文和华莱士对这一原则的理解都来自阅读经济学家托马斯·马尔萨斯的一篇文章,他讨论了这一原则与人口的关系。 第三,后代的特征各不相同,这些变异是遗传的。 在这三个原则中,达尔文和华莱士认为,具有继承特征、使他们能够最好地竞争有限资源的后代将比那些具有竞争能力较差的变体的人存活下来并拥有更多的后代。 由于特征是遗传的,因此这些特征将在下一代中得到更好的体现。 这将导致人口在几代人之间发生变化,达尔文称之为 “变性后裔”。
1858 年,在伦敦林纳协会之前,一起阅读了达尔文和华莱士(图\(\PageIndex{2}\))介绍自然选择概念的论文。 次年,达尔文出版了《论物种起源》一书,该书相当详细地概述了他关于通过自然选择进化的论点。
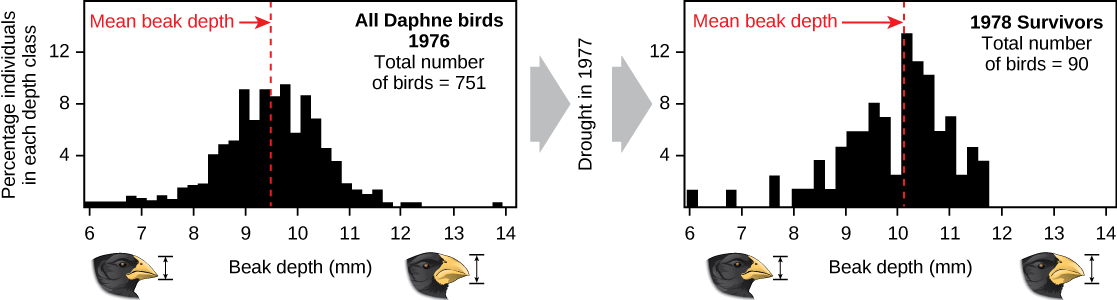
通过自然选择演示进化可能很耗时。 最好的示范之一是那些有助于激发理论灵感的鸟,即加拉帕戈斯雀科。 自1976年以来,彼得和罗斯玛丽·格兰特及其同事每年都对加拉帕戈斯雀科种群进行研究,并为自然选择的运作提供了重要的示范。 Grants 发现,加拉帕戈斯岛达芙妮·梅杰岛上中等地面雀的喙形状代代相传。 中等地面雀以种子为食。 这些鸟继承了账单形状的变化,有些人的账单很大,而另一些人的账单更薄。 大嘴鸟以大型硬种子为食,效率更高,而小嘴鸟以小而软的种子为食,效率更高。 1977 年,干旱时期改变了岛上的植被。 在此之后,种子数量急剧下降:小而软的种子的减少幅度大于大型硬种子的减少。 次年,大嘴鸟比小嘴鸟存活得更好。 在干旱后的第二年,当补助金在大幅减少的人口中测量了喙的大小时,他们发现平均账单规模更大(图\(\PageIndex{3}\))。 这清楚地证明了种子供应导致账单规模的自然选择(存活率差异)。 补助金研究了账单规模的继承,并且知道幸存的大嘴鸟往往会产生账单更大的后代,因此这种选择将导致账单规模的演变。 Grants随后的研究表明,该物种的账单规模是随着岛上条件的变化而选择和演变的。 这种演变既发生在较大的账单上(如本例中),也发生在大种子变得稀有时变为较小的账单。
变异和适应
只有在人群中的个体之间存在变异或差异时,才能进行自然选择。 重要的是,这些差异必须有一定的遗传基础;否则,选择不会导致下一代的改变。 这很关键,因为个体之间的变异可能是由非遗传原因引起的,例如个体因为营养好而不是基因不同而变高。
人群的遗传多样性来自两个主要来源:突变和有性生殖。 突变是DNA的改变,是任何人群中新等位基因或新遗传变异的最终来源。 具有突变基因的个体可能与人群中的其他个体具有不同的特征。 但是,情况并非总是如此。 突变可能对生物体外观(或表型)产生以下三种结果之一:
- 突变可能会影响生物体的表型,使其健康状况降低,存活的可能性降低,从而减少后代。
- 突变可能会产生对健康有益影响的表型。
- 许多被称为中性突变的突变对健康没有影响。
突变还可能对以其表型表达它们的生物的适应性产生各种效应大小,从小效应到大效应不等。 减数分裂中的有性繁殖和交叉也会导致遗传多样性:当两个父母繁殖时,独特的等位基因组合会聚在一起,产生独特的基因型,从而在每个后代中产生表型。
一种有助于生物在当前环境中存活和繁殖的可遗传特征称为适应。 适应是生物与环境的 “匹配”。 当遗传变异范围随着时间的推移发生变化,从而增加或维持种群与环境的匹配时,就会适应环境。 雀嘴的变化代代相传,从而适应了食物供应。
一个特质是否有利取决于当时的环境。 相同的特征并不总是具有相同的相对优势或劣势,因为环境条件可能会发生变化。 例如,持有大额钞票的雀科在一种环境中受益,而小额钞票则处于劣势;在另一种环境下,这种关系发生了逆转。
进化模式
物种的进化导致了形态和功能的巨大差异。 当两个物种从一个共同点朝不同的方向进化时,它被称为发散进化。 这种不同的进化可以从开花植物的生殖器官的形式中看出,它们具有相同的基本解剖结构;但是,由于在不同的物理环境中进行选择以及对不同种类的传粉媒介的适应,它们看起来可能会大不相同(图\(\PageIndex{4}\))。

在其他情况下,相似的表型在远距离相关的物种中独立进化。 例如,蝙蝠和昆虫都有飞行进化,它们都有我们称之为翅膀的结构,可以适应飞行。 但是,蝙蝠和昆虫的翅膀是由截然不同的原始结构进化而来的。 当不同物种通过独立进化产生相似结构时,它被称为会聚进化。 蝙蝠和昆虫的翅膀被称为类似结构;它们的功能和外观相似,但并不具有共同祖先的起源。 相反,它们在两个谱系中独立进化。 蜂鸟和鸵鸟的翅膀是同源结构,这意味着它们有相似之处(尽管它们的差异是由进化分歧造成的)。 蜂鸟和鸵鸟的翅膀不是在蜂鸟谱系和鸵鸟谱系中独立进化的,它们来自一个有翅膀的共同祖先。
《现代合成》
在达尔文和华莱士发展自然选择理念时,遗传机制即遗传学还不为人所知。 这种缺乏理解是理解进化许多方面的绊脚石。 事实上,混合遗传是当时占主导地位(也是错误的)遗传理论,这使得人们很难理解自然选择会如何运作。 达尔文和华莱士没有意识到奥地利僧侣格雷戈尔·孟德尔的遗传学著作,该著作于 1866 年出版,也就是《论物种起源》出版后不久。 孟德尔的作品在二十世纪初被重新发现,当时遗传学家迅速了解了继承的基础知识。 最初,新发现的基因颗粒性质使生物学家难以理解逐渐进化是如何发生的。 但是在接下来的几十年中,遗传学和进化被纳入了所谓的现代合成中,即对自然选择与遗传学之间关系的连贯理解,这种理解在20世纪40年代形成,如今已被普遍接受。 总而言之,现代综合描述了自然选择等进化压力如何影响种群的基因构成,进而描述了这如何导致种群和物种的逐渐进化。 该理论还将种群随着时间的推移逐渐变化(称为微进化)与产生具有巨大差异特征的新物种和更高分类群的过程(称为宏观进化)联系起来。
种群遗传学
回想一下,特定角色的基因可能有多个变体或等位基因,它们编码与该角色相关的不同特征。 例如,在人类的ABO血型系统中,三个等位基因决定红细胞表面的特定血型蛋白。 二倍体生物群体中的每个人只能携带特定基因的两个等位基因,但构成该群体的个体中可能存在两个以上的等位基因。 孟德尔跟随等位基因,因为它们是从父母遗传给后代的。 二十世纪初,生物学家开始在一个被称为群体遗传学的研究领域研究人群中所有等位基因会发生什么。
到目前为止,我们将进化定义为生物群体特征的变化,但表型变化的背后是遗传变化。 在种群遗传学术语中,进化被定义为群体中等位基因频率的变化。 以 ABO 系统为例,其中一个等位基因 I A 的频率是该等位基因的拷贝数除以群体中 ABO 基因的所有拷贝数。 例如,约旦的一项研究发现 I A 的频率为 26.1%。 2 I B、I 0 等位基因分别占等位基因的 13.4% 和 60.5%,所有频率加起来为 100%。 随着时间的推移,这种频率的变化将构成人口的演变。
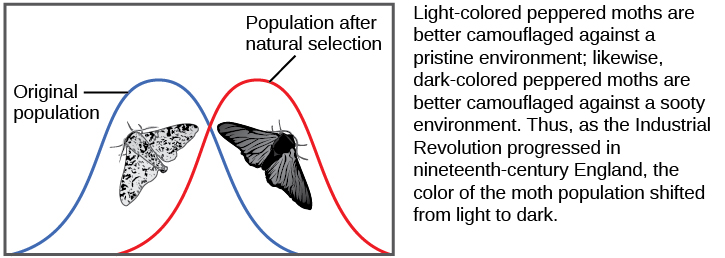
种群的等位基因频率可以通过多种方式改变。 其中一种方法是自然选择。 如果给定的等位基因赋予的表型允许个体有更多的存活和繁殖的后代,那么该等位基因由于被这些后代遗传,在下一代中出现的频率将更高。 由于等位基因频率加起来总是达到100%,因此一个等位基因频率的增加总是意味着一个或多个其他等位基因的相应降低。 高度有益的等位基因可能会在几代人的时间里以这种方式变得 “固定”,这意味着种群中的每个人都将携带该等位基因。 同样,有害的等位基因可能会很快从基因库中消除,基因库是群体中所有等位基因的总和。 种群遗传学研究的一部分是追踪选择性力如何随着时间的推移改变人群中的等位基因频率,这可以为科学家提供有关可能对给定人群起作用的选择力的线索。 研究胡椒蛾的翅膀颜色因烟灰覆盖的树干而从斑驳的白色变为深色,然后在工厂停止生产如此多的烟灰时变为斑驳的白色,这是研究自然种群进化的典型例子(图\(\PageIndex{5}\))。
二十世纪初,英国数学家戈弗雷·哈迪和德国医生威廉·温伯格独立解释了一个有点违反直觉的概念。 哈迪最初的解释是对一种误解的回应,即为什么掩盖隐性等位基因的 “显性” 等位基因在消灭所有其他等位基因之前不应该增加人群中的频率。 这个问题源于人们对 “占主导地位” 含义的普遍困惑,但它迫使甚至不是生物学家的哈迪指出,如果没有影响等位基因频率的因素,这些频率将从一代到下一代保持不变。 这个原理现在被称为哈迪-温伯格均衡。 该理论指出,种群的等位基因和基因型频率本质上是稳定的,除非某种进化力量作用于种群,否则种群将一代又一代地以相同的比例携带相同的等位基因。 总体而言,个体看起来基本相同,这与等位基因是显性还是隐性无关。 将破坏平衡的四种最重要的进化力量是自然选择、突变、遗传漂移和迁入或迁出种群。 第五个因素是非随机交配,也将破坏哈迪-温伯格平衡,但只能通过改变基因型频率而不是等位基因频率来实现。 在非随机交配中,个体更有可能与相似的个体(或不同个体)交配,而不是随机交配。 由于非随机交配不会改变等位基因频率,因此不会直接导致进化。 已经描述了自然选择。 突变从另一个等位基因中产生一个等位基因,并且每代都会使一个等位基因的频率变化很小但连续的量。 每个等位基因都是由低而恒定的突变率产生的,如果没有其他力量作用于等位基因,则该变异率将慢慢增加该等位基因在群体中的频率。 如果自然选择对等位基因起作用,它将以较低的速率从群体中移除,从而产生选择和突变之间的平衡产生的频率。 这是遗传疾病在人类中以非常低的频率存在的原因之一。 如果等位基因受到选择的青睐,则其频率会增加。 当种群较少时,遗传漂移会导致等位基因频率的随机变化。 如下一节所述,遗传漂移在进化中通常很重要。 最后,如果一个物种的两个种群具有不同的等位基因频率,则个体在它们之间的迁移将导致两个种群的频率变化。 碰巧的是,没有人群中的一个或多个过程不起作用,因此种群总是在演变,哈迪-温伯格均衡永远无法准确观察。 但是,Hardy-Weinberg原理为科学家提供了对非进化人群中等位基因频率的基准预期,他们可以将不断演变的种群与之进行比较,从而推断出哪些进化力量可能在起作用。 如果等位基因或基因型的频率偏离哈迪-温伯格原理的预期值,则种群正在演变。
达尔文发现了一个自然选择的特例,他称之为性选择。 性选择会影响个体交配从而产生后代的能力,它会导致戏剧性特征的演变,这些特征在生存方面往往显得适应不良,但由于它们能给主人带来更大的生殖成功而持续存在。 性选择有两种方式:通过男性和男性争夺伴侣,以及通过女性选择伴侣。 男性与男性之间的竞争采取男性之间的冲突的形式,这种冲突通常是仪式化的,但也可能对男性的生存构成重大威胁。 有时候,竞争是争夺领地,雌性更有可能与地域质量更高的雄性交配。 当雌性根据特定的特征(例如羽毛颜色、交配舞的表演或精致的建筑物)选择雄性时,就会出现雌性的选择。 在某些情况下,在交配过程中,雄性竞争和雌性的选择结合在一起。 在每种情况下,选择的特征,例如战斗能力或羽毛颜色和长度,都会在雄性中得到增强。 总的来说,人们认为性选择可以发展到自然选择而不是角色的进一步增强阻止其进一步进化的程度,因为它会对雄性的生存能力产生负面影响。 例如,五颜六色的羽毛或精心制作的展示会使雄性对捕食者更加明显。
摘要
自然选择的进化源于三个条件:物种中的个体各不相同,其中一些变异是可遗传的,生物的后代比资源所能支持的要多。 结果是,与具有不同特征的人相比,具有相对优势变异的人更有可能存活并获得更高的生殖率。 优势特征将更大比例地传给后代。 因此,该特征在下一代和后代中将具有更高的代表性,从而导致人群的遗传变化。
进化论的现代综合源于达尔文、华莱士和孟德尔关于进化和遗传的思想的调和。 种群遗传学是一个理论框架,用于通过等位基因频率的变化描述种群的进化变化。 种群遗传学将进化定义为代际等位基因频率的变化。 在没有进化力的情况下,种群中的等位基因频率不会改变;这就是所谓的哈迪-温伯格平衡原理。 但是,在所有种群中,突变、自然选择、遗传漂移和迁移都会改变等位基因频率。
脚注
- 1 查尔斯·达尔文,《H.M.S. Beagle 环游世界期间访问的国家的自然历史和地质研究杂志》,由罗得角菲茨·罗伊上尉指挥,第二版 (伦敦:约翰·默里,1860 年),http://www.archive.org/details/journalofresea00darw。
- 2 Sahar S. Hanania、Dhia S. Hassawi 和 Nidal M. Irshaid,“约旦人群中 ABO 血型系统的等位基因频率和分子基因型”,《医学科学杂志》 7(2007):51-58,doi: 10.3923/jms.2007.51.58
词汇表
- 适应
- 生物体中可遗传的特征或行为,有助于其在当前环境中生存
- 类似的结构
- 这种结构之所以相似,是因为在相似的选择压力下进化导致趋同进化,而不相似是因为来自共同祖先的血统
- 趋同进化
- 这种进化导致不同物种的形式相似
- 发散进化
- 这种进化导致两个具有共同祖先的物种以不同的形式出现
- 基因库
- 人群中所有个体携带的所有等位基因
- 遗传漂移
- 偶然对人群基因库的影响
- 同源结构
- 这种结构之所以相似,是因为来自共同祖先
- 继承获得的特征
- 这句话描述了拉马克提出的进化机制,在这种机制中,个人通过使用或废弃获得的特征可以传给他们的后代,从而导致种群的进化变化
- 宏观进化
- 古生物学时代出现了更大规模的进化变化
- 微进化
- 人群遗传结构的变化(即等位基因频率)
- 迁徙
- 人口中的个体向新地点的移动;在种群遗传学中,它是指个体及其等位基因从一个种群向另一个种群的移动,这可能会改变新旧种群中的等位基因频率
- 现代合成
- 在 20 世纪 40 年代形成、今天被普遍接受的总体进化范式
- 自然选择
- 在具有良好遗传特征的人群中,个体的相对存活率和繁殖率更高,从而导致进化变化
- 种群遗传学
- 研究选择性力如何随着时间的推移改变人群中的等位基因频率
- 变体
- 人群中等位基因的种类