
9.5: 如何调节基因
- Page ID
- 199284

为了使细胞正常运作,必须在适当的时间合成必需的蛋白质。 所有生物和细胞都控制或调节其DNA转录和转化为蛋白质。 开启基因产生 RNA 和蛋白质的过程称为基因表达。 无论是在简单的单细胞生物体中还是在复杂的多细胞生物体中,每个细胞都控制着其基因的表达时间和方式。 要做到这一点,必须有一种机制来控制何时表达基因以制造RNA和蛋白质,产生了多少蛋白质,以及何时该停止制造该蛋白质,因为不再需要这种蛋白质。
多细胞生物体中的细胞是特殊的;不同组织中的细胞看起来截然不同,功能也不同。 例如,肌肉细胞与肝细胞有很大不同,后者与皮肤细胞有很大不同。 这些差异是这些细胞中不同基因组表达的结果。 所有细胞都有必须自己发挥的某些基本功能,例如将糖分子中的能量转化为ATP中的能量。 每个细胞还有许多未表达的基因,并且表达了许多其他细胞未表达的基因,因此它可以发挥其特殊功能。 此外,细胞将在不同的时间开启或关闭某些基因,以应对环境的变化,或者在生物体发育过程中的不同时期。 单细胞生物,包括真核生物和原核生物,也会根据环境的需求开启和关闭基因,以便它们能够对特殊条件做出反应。
基因表达的控制极其复杂。 这个过程中的故障对细胞有害,并可能导致许多疾病的发展,包括癌症。
原核生物与真核生物基因表达
要了解基因表达是如何调节的,我们必须首先了解基因是如何成为细胞中的功能性蛋白质的。 该过程发生在原核细胞和真核细胞中,只是以略有不同的方式发生。
由于原核生物缺乏细胞核,因此转录和翻译过程几乎同时发生。 当不再需要蛋白质时,转录就会停止。 因此,控制原核细胞中表达哪种类型和多少蛋白质的主要方法是通过调节 DNA 转录为 RNA。 所有后续步骤都会自动执行。 当需要更多蛋白质时,会发生更多的转录。 因此,在原核细胞中,基因表达的控制几乎完全处于转录水平。
这种控制的第一个例子是使用 E 发现的。 1950年代和1960年代由法国研究人员创作@@ 的大肠杆菌,被称为 lac 操作元。 lac operon 是一段具有三个相邻基因的 DNA 片段,它们编码参与乳糖吸收和代谢的蛋白质,乳糖是 E 的食物来源。 大肠杆菌。 当细菌环境中不存在乳糖时,lac 基因会被少量转录。 当存在乳糖时,基因会被转录,细菌能够使用乳糖作为食物来源。 操纵子还包含启动子序列,RNA聚合酶与该序列结合以开始转录;启动子和三个基因之间有一个称为操作符的区域。 当不存在乳糖时,一种被称为抑制剂的蛋白质会与操作员结合,阻止 RNA 聚合酶与启动子结合,除非在极少数情况下。 因此,这三个基因的蛋白质产物很少产生。 当存在乳糖时,乳糖代谢的最终产物会与抑制蛋白结合,阻止其与操作员结合。 这允许RNA聚合酶与启动子结合并自由转录这三个基因,从而使生物体能够代谢乳糖。
相比之下,真核细胞具有细胞内细胞器,而且要复杂得多。 回想一下,在真核细胞中,DNA包含在细胞核内,并在那里被转录为mRNA。 然后,新合成的 mRNA 从细胞核输送到细胞质中,核糖体在细胞质中将 mRNA 转化为蛋白质。 转录和翻译过程在物理上被核膜分开;转录仅发生在细胞核内,翻译仅发生在细胞质的核外。 基因表达的调节可能发生在过程的各个阶段(图\(\PageIndex{1}\))。 当 DNA 解开并从核小体中松开以结合转录因子(表观遗传学水平)、RNA 被转录(转录水平)、转录后 RNA 经过处理并输出到细胞质时(转录后水平)、RNA 被翻译时,可能会发生调节转化为蛋白质(翻译水平),或蛋白质生成后(翻译后水平)。
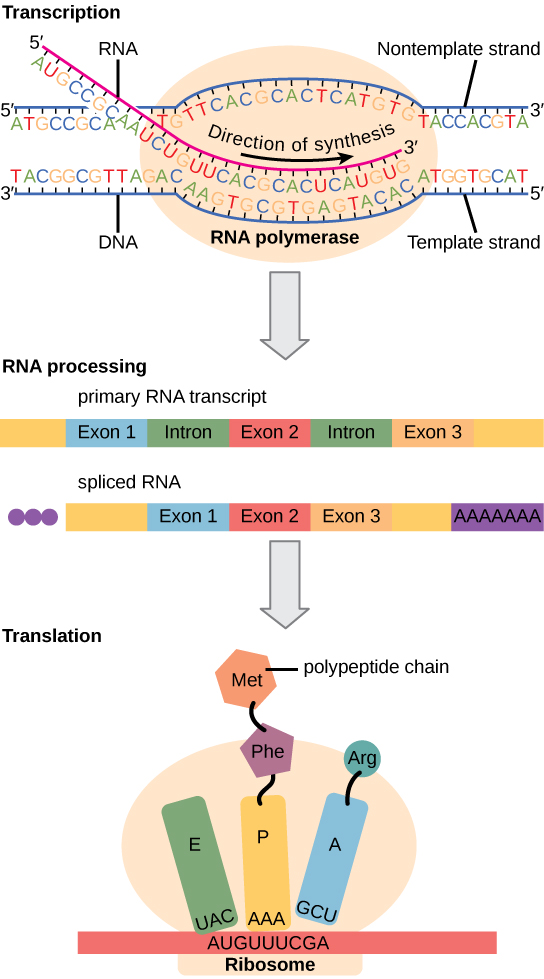
表中总结了原核生物和真核生物之间在基因表达调控方面的差异\(\PageIndex{1}\)。
| 原核生物 | 真核生物 |
|---|---|
| 缺少核 | 包含核 |
| RNA 转录和蛋白质翻译几乎同时发生 |
|
| 基因表达主要在转录水平上受到调节 | 基因表达在多个层面上受到调节(表观遗传学、转录、转录后、翻译和翻译后) |
行动中的进化:替代性 RNA 拼接
在20世纪70年代,首次观察到表现出替代性RNA拼接的基因。 替代性RNA拼接是一种机制,当从转录本中移除不同的内含子(有时是外显子)组合时,允许从一个基因中产生不同的蛋白质产物(图\(\PageIndex{2}\))。 这种替代拼接可能是随意的,但更常见的是它受到控制并起到基因调控机制的作用,不同的拼接替代方案的频率由细胞控制,以此来控制不同细胞中或不同阶段不同蛋白质产物的产生开发。 现在,人们认为替代拼接是真核生物中常见的基因调控机制;根据一项估计,人类中有70%的基因是通过替代拼接以多种蛋白质表达的。
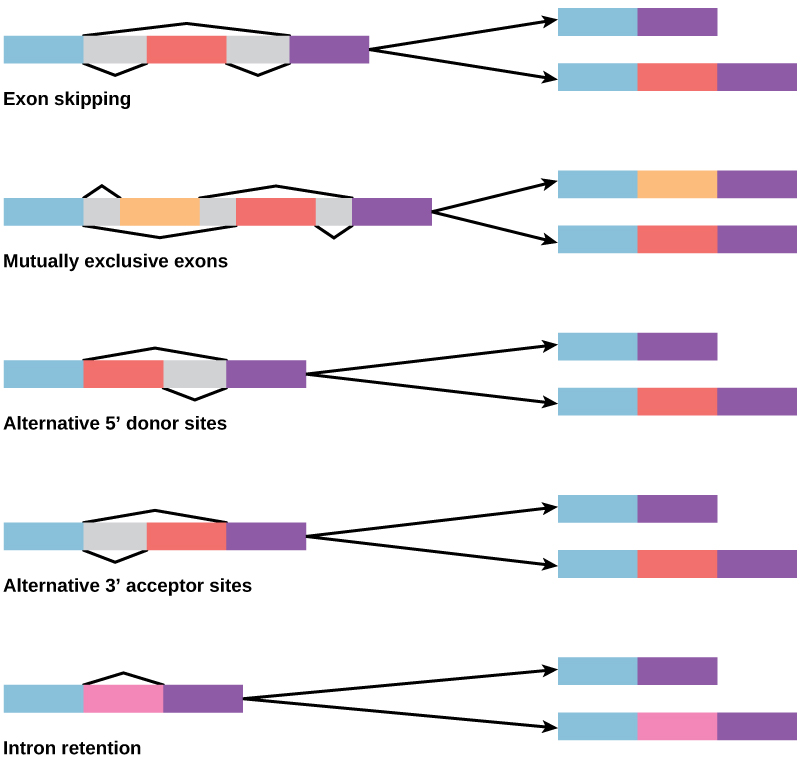
替代拼接会如何演变? 内含子有开头和结尾识别序列,很容易想象拼接机制无法识别内含子的末端并找到下一个内含子的结尾,从而移除两个内含子和中间的外显子。 实际上,有防止这种外显子跳过的机制,但是突变很可能导致其失效。 这种 “错误” 很可能会产生不起作用的蛋白质。 事实上,许多遗传疾病的起因是替代性拼接而不是序列中的突变。 但是,替代拼接将在不损失原始蛋白质的情况下产生蛋白质变体,从而为新变体适应新功能开辟可能性。 基因复制以类似的方式在新功能的演变中发挥了重要作用——它提供了可以在不消灭原始功能蛋白的情况下进化的基因。
摘要
虽然生物体内的所有体细胞都含有相同的 DNA,但并非该生物体内的所有细胞都表达相同的蛋白质。 原核生物在每个细胞中表达它们编码的全部 DNA,但不一定同时表达所有的 DNA。 蛋白质只有在需要时才表达。 真核生物表达编码在任何给定细胞中的 DNA 子集。 在每种细胞类型中,蛋白质的类型和数量都通过控制基因表达来调节。 为了表达蛋白质,首先将DNA转录为RNA,然后将其转化为蛋白质。 在原核细胞中,这些过程几乎同时发生。 在真核细胞中,转录发生在细胞核中,与细胞质中发生的翻译是分开的。 原核生物中的基因表达仅在转录水平上受到调节,而在真核细胞中,基因表达在表观遗传学、转录后、翻译和翻译后水平受到调节。
词汇表
- 替代 RNA 拼接
- 真核生物中的一种转录后基因调控机制,其中单个基因通过 RNA 转录本的替代拼接组合产生多种蛋白质产物
- 表观遗传学
- 描述非遗传调节因素,例如控制染色体中基因可及性的组蛋白和DNA的修饰变化
- 基因表达
- 控制基因是否表达的过程
- 转录后
- 在 RNA 分子产生之后但在转化为蛋白质之前控制基因表达
- 翻译后
- 蛋白质产生后控制基因表达