
18.1: 免疫系统的架构
- Page ID
- 200174

学习目标
- 定义记忆、主要反应、次要反应和特异性
- 区分体液免疫和细胞免疫
- 区分抗原、抗原表位和半抗原
- 描述抗体的结构和功能,区分不同类别的抗体
临床重点:第 1 部分
奥利维亚是一名一岁的婴儿,被父母带到急诊室,父母报告了她的症状:哭得过多、烦躁、对光线敏感、异常嗜睡和呕吐。 医生感觉到奥利维亚的喉咙和腋窝淋巴结肿大。 此外,腹部在脾脏上方的区域肿胀而变软。
练习\(\PageIndex{1}\)
- 这些症状意味着什么?
- 可能要下令进行哪些测试来尝试诊断问题?
适应性免疫由两个重要特征定义:特异性和记忆力。 特异性是指适应性免疫系统靶向特定病原体的能力,记忆力是指其对先前接触过的病原体做出快速反应的能力。 例如,当一个人从水痘中恢复过来时,人体会对感染产生记忆,这可以特别保护其免受致病因子,即水痘带状疱疹病毒的侵害,如果它以后再次接触该病毒。
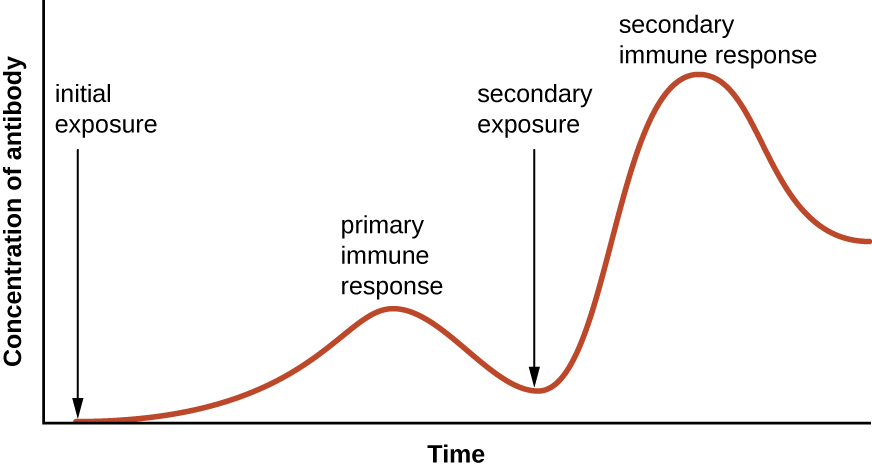
特异性和记忆力是通过对参与免疫反应的某些细胞进行编程来实现的,以便对随后的病原体暴露做出快速反应。 这种编程是由于首次接触病原体或疫苗而发生的,这会触发初级反应。 由于人体对第一次曝光的记忆,随后的曝光会产生更快、更强的二次反应(图\(\PageIndex{1}\))。 但是,这种次要反应是有关病原体所特有的。 例如,接触一种病毒(例如水痘带状疱疹病毒)无法抵御其他病毒性疾病(例如麻疹、腮腺炎或脊髓灰质炎)。
适应性特异性免疫涉及两种不同细胞类型的作用:B 淋巴细胞(B 细胞)和 T 淋巴细胞(T 细胞)。 尽管 B 细胞和 T 细胞来自常见的造血干细胞分化途径,但它们的成熟位点及其在适应性免疫中的作用却大不相同。
B 细胞在骨髓中成熟,负责产生称为抗体或免疫球蛋白的糖蛋白。 抗体参与人体对细胞外环境中病原体和毒素的防御。 涉及 B 细胞和抗体产生的适应性特异性免疫机制被称为体液免疫。 T 细胞的成熟发生在胸腺中。 T 细胞是先天免疫反应和适应性免疫反应的中心协调者。 它们还负责破坏感染细胞内病原体的细胞。 T 细胞靶向和破坏细胞内病原体称为细胞介导免疫或细胞免疫。
练习\(\PageIndex{2}\)
- 列出适应性免疫的两个决定性特征。
- 解释初级和次要免疫反应之间的区别。
- 体液免疫和细胞免疫有何不同?
抗原
适应性免疫防御的激活是由称为抗原的病原体特异性分子结构触发的。 抗原与《病原体识别和吞噬作用》中讨论的病原体相关分子模式 (PAMP) 相似;但是,PAMP 是在许多病原体上发现的分子结构,而抗原是特定病原体所独有的。 例如,刺激水痘适应性免疫的抗原是水痘带状疱疹病毒所独有的,但与其他病毒病原体相关的抗原有显著不同。
抗原一词最初用于描述刺激抗体产生的分子;实际上,该术语来自抗体和 gen erator这两个词的组合,刺激抗体产生的分子据说是抗原性的。 但是,抗原的作用不仅限于体液免疫和抗体的产生;抗原在刺激细胞免疫方面也起着至关重要的作用,因此,抗原有时被更准确地称为免疫原。 但是,在本文中,我们通常将它们称为抗原。
病原体具有多种可能含有抗原的结构。 例如,来自细菌细胞的抗原可能与它们的胶囊、细胞壁、纤维、鞭毛或 pili 有关。 细菌抗原也可能与它们分泌的细胞外毒素和酶有关。 病毒具有与其衣壳、包膜和用于附着在细胞上的尖刺结构相关的各种抗原。
抗原可能属于任意数量的分子类别,包括碳水化合物、脂质、核酸、蛋白质和这些分子的组合。 不同类别的抗原在刺激适应性免疫防御的能力以及它们刺激的反应类型(体液或细胞)方面各不相同。 抗原分子的结构复杂性是影响其抗原潜力的重要因素。 通常,更复杂的分子作为抗原更有效。 例如,蛋白质的三维复杂结构使其成为最有效和最有效的抗原,能够刺激体液和细胞免疫。 相比之下,碳水化合物的结构不那么复杂,因此作为抗原的效果较差;它们只能刺激体液免疫防御。 脂质和核酸是抗原性最低的分子,在某些情况下,只有在与蛋白质或碳水化合物结合形成糖脂、脂蛋白或核蛋白时才会产生抗原性。
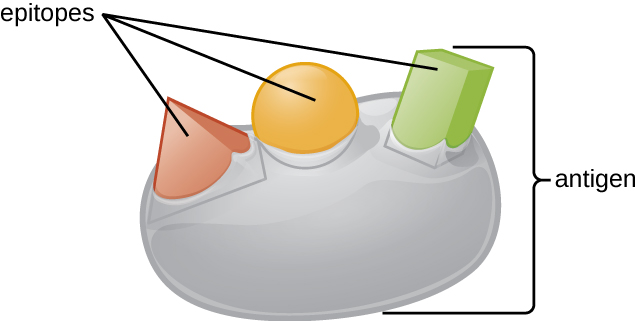
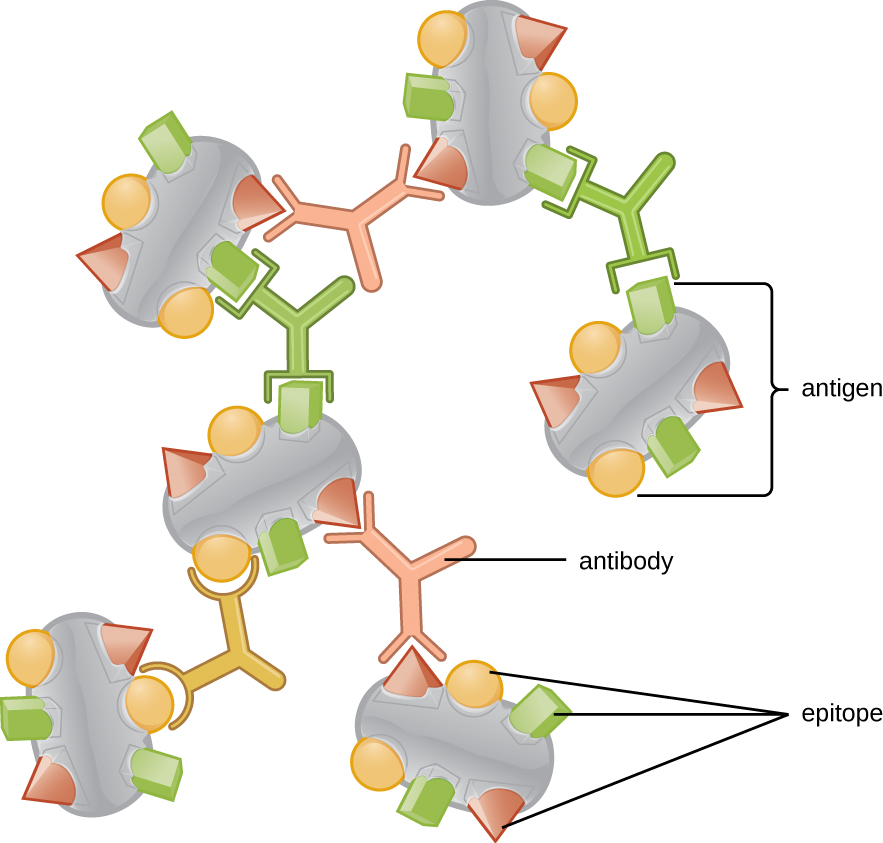
抗原的三维复杂性如此重要的原因之一是,抗体和T细胞无法识别整个抗原并与之相互作用,而是在抗原表面具有较小的暴露区域,称为抗原表位。 单个抗原可能具有几种不同的抗原表位(图\(\PageIndex{2}\)),不同的抗体可能与同一抗原上的不同表位结合(图\(\PageIndex{3}\))。 例如,细菌鞭毛是一种庞大而复杂的蛋白质结构,可以拥有数百甚至数千个具有独特三维结构的表位。 此外,来自不同细菌物种(甚至是同一物种的菌株)的鞭毛含有独特的抗原表位,只能通过特异性抗体结合。
抗原的大小是其抗原潜力的另一个重要因素。 尽管鞭毛等大型抗原结构具有多个抗原表位,但有些分子太小,本身无法产生抗原。 这种被称为半抗原的分子本质上是游离的表位,不是较大抗原复杂的三维结构的一部分。 为了使半抗原成为抗原,它必须首先附着在较大的载体分子(通常是蛋白质)上才能产生共轭抗原。 然后,响应偶联抗原而产生的半抗原特异性抗体能够与未共轭的游离半抗原分子相互作用。 目前尚不清楚半抗原与任何特定的病原体有关,但它们是造成某些过敏反应的原因。 例如,hapten urushiol(一种存在于植物油中的分子,会引起毒藤),它会引起免疫反应,从而导致严重的皮疹(称为接触性皮炎)。 同样,半抗原青霉素可引起对青霉素类药物的过敏反应。
练习\(\PageIndex{3}\)
- 抗原和表位有什么区别?
- 哪些因素会影响抗原的抗原潜力?
- 为什么半抗原通常不是抗原,它们是如何变成抗原的?
抗体
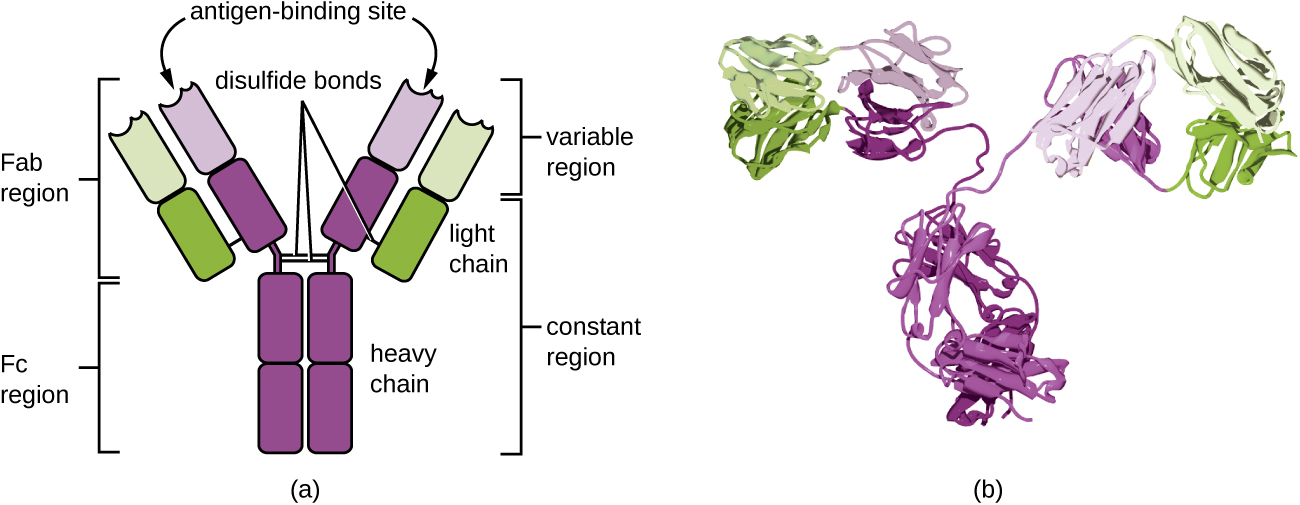
抗体(也称为免疫球蛋白)是存在于血液和组织液中的糖蛋白。 抗体单体的基本结构由四条由二硫键连接在一起的蛋白质链组成(图\(\PageIndex{4}\))。 二硫键是在两个半胱氨酸氨基酸上发现的磺酰基团之间的共价键。 两条最大的链条彼此相同,被称为重链。 两条较小的链条也彼此相同,被称为轻链。 重链和轻链结合在一起,形成基本的Y形结构。
Y 形抗体分子的两个 “臂” 被称为 Fab 区域,用于 “抗原结合片段”。 Fab 区域的远端是可变区域,它是抗原结合的位点。 可变区域中的氨基酸序列决定了三维结构,从而决定了Fab区域能够与之结合的特定三维表位。 尽管对于单个抗体分子的每个分支,Fab区域的抗原表位特异性是相同的,但该区域在具有不同表位特异性的抗体之间显示出很大的差异。 与 Fab 区域结合是中和病原体、病原体的凝集或聚集以及抗体依赖性细胞介导的细胞毒性所必需的。
抗体分子的恒定区域包括 Y 的树干和 Y 的每只手臂的下半部分。Y 的树干也被称为 Fc 区域,用于 “结晶片段”,是抗体介导的 opsonization 期间补体因子结合和结合吞噬细胞的位点。
练习\(\PageIndex{4}\)
描述 Fab 区域和 Fc 区域的不同功能。
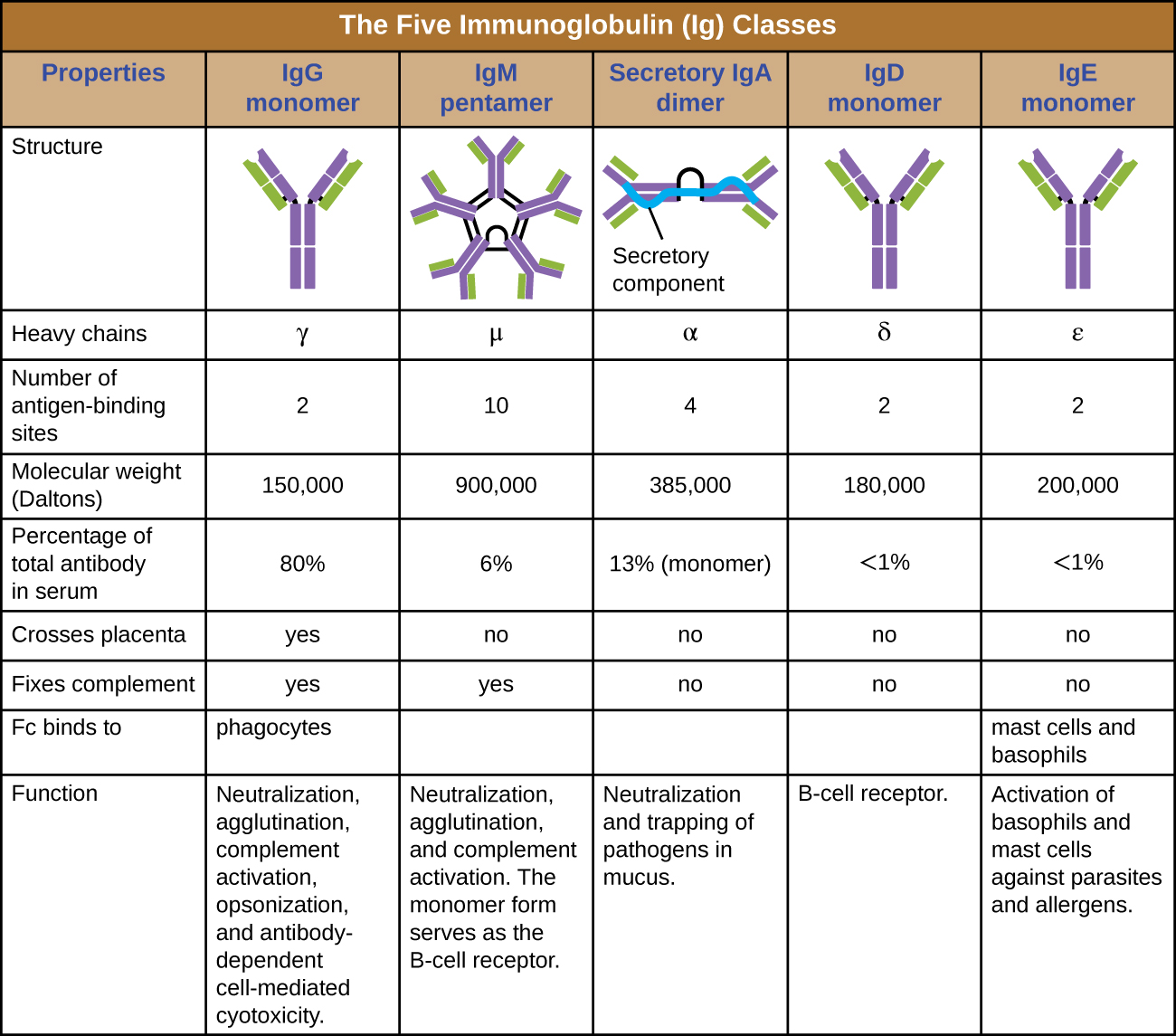
抗体类别
抗体分子的恒定区域决定其类别或同型型。 五类抗体是 IgG、IgM、IgA、IgD 和 IgE。 每个类别都拥有独特的重链,分别由希腊字母 γ、μ、α、β 和 ε表示。 抗体类别在血清丰度、排列、身体作用部位、功能作用和大小方面也表现出重要差异(图\(\PageIndex{5}\))。
IgG 是一种单体,是迄今为止人体血液中最丰富的抗体,约占血清抗体总量的 80%。 IgG 可有效渗透到组织空间,是唯一能够穿过胎盘屏障的抗体类别,在怀孕期间为发育中的胎儿提供被动免疫力。 就其在人体抵御病原体中的作用而言,IgG也是用途最广泛的抗体类别。
IgM 最初以单体膜结合形式产生,作为 B 细胞上的抗原结合受体。 分泌形式的 IgM 组装成五聚体,五个 IgM 单体通过一种叫做 J 链的蛋白质结构结合在一起。 尽管J链相对于五种单体的Fc区域的位置使IgM无法发挥IgG的某些功能,但与五聚体IgM相关的十个可用Fab位点使其成为人体防御库中的重要抗体。 IgM 是 B 细胞在原发和继发免疫反应期间产生和分泌的第一种抗体,使病原体特异性 IgM 成为活跃或近期感染期间有价值的诊断标志。
IgA约占血清抗体总量的13%,分泌型IgA是粘液分泌物中发现的最常见和最丰富的抗体类别,可保护粘膜。 IgA 也可以在其他分泌物中找到,例如母乳、眼泪和唾液。 Secretory IgA 组装成二聚体形式,两个单体由一种称为分泌成分的蛋白质结构连接在一起。 分泌性IgA的重要功能之一是将病原体捕获在粘液中,以便以后可以将其从体内消除。
与 IgM 类似,IgD 是一种存在于 B 细胞表面的膜结合单体,用作抗原结合受体。 但是,IgD 不是由 B 细胞分泌的,血清中仅检测到微量。 这些微量很可能来自于旧 B 细胞的降解和 IgD 分子从其细胞质膜中释放。
IgE 是血清中含量最少的抗体类别。 与IgG一样,它以单体形式分泌,但其在适应性免疫中的作用仅限于抗寄生虫防御。 IgE 的 Fc 区域与嗜碱性粒细胞和肥大细胞结合。 然后,结合的 IgE 的 Fab 区域与特定的抗原表位相互作用,导致细胞释放出强效的促炎介质。 肥大细胞和嗜碱性粒细胞的激活所产生的炎症反应有助于抵御寄生虫,但这种反应也是过敏反应的核心(参见免疫系统疾病)。
练习\(\PageIndex{5}\)
- 抗体分子的哪一部分决定其类别?
- 哪类抗体参与寄生虫防护?
- 描述 IgM 和 IgG 在结构上的差异。
抗原-抗体相互作用
不同类别的抗体在人体对病原体的防御中起着重要作用。 这些功能包括中和病原体、吞噬作用的 opsonic 作用、凝集、补体激活和抗体依赖性细胞介导的细胞毒性。 对于其中的大多数功能,抗体还提供了适应性特异性免疫和先天非特异性免疫之间的重要联系。
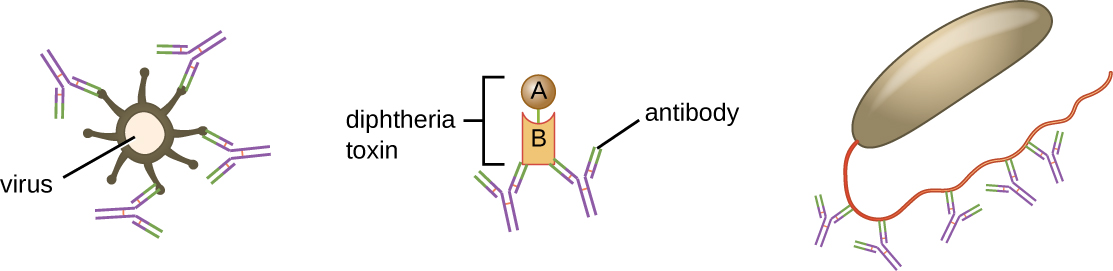
中和涉及将某些抗体(IgG、IgM 或 IgA)与病原体或毒素表面的抗原表位结合,防止它们附着在细胞上。 例如,Secretory IgA 可以与特定的病原体结合并阻断与肠道粘膜细胞的初始附着。 同样,特异性抗体可以与某些毒素结合,阻止它们附着在靶细胞上,从而中和它们的毒性作用。 病毒可以通过相同的机制被中和并防止其感染细胞(图\(\PageIndex{6}\))。
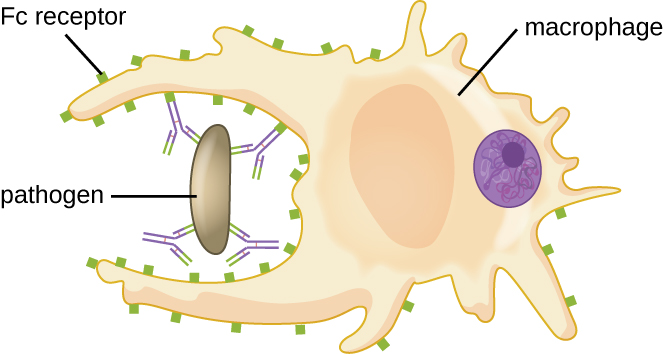
正如《化学防御》中所述,opsonization 是用互补因子、C 反应蛋白和血清淀粉样蛋白 A 等分子包裹病原体,以帮助吞噬细胞结合,促进吞噬作用。 IgG 抗体还可以作为优秀的 opsonins,将其晶圆厂位点与病原体表面的特定抗原表位结合。 吞噬细胞,例如巨噬细胞、树突状细胞和中性粒细胞,其表面具有识别和结合IgG分子Fc部分的受体;因此,IgG帮助这些吞噬细胞附着并吞没它们所结合的病原体(图\(\PageIndex{7}\))。
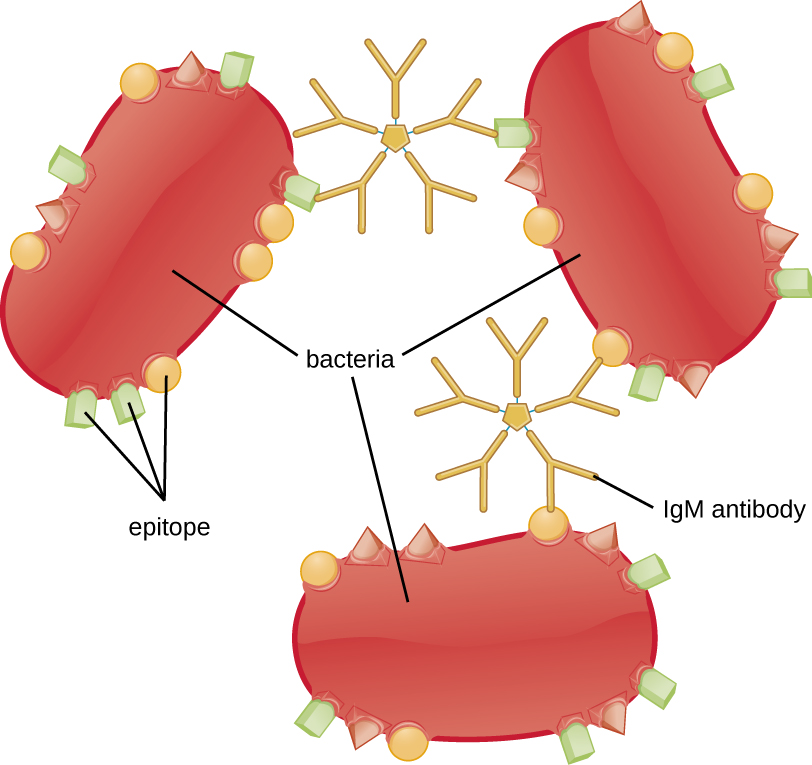
凝集或聚集涉及通过抗体将病原体交联以产生大型聚集体(图\(\PageIndex{8}\))。 IgG 有两个 Fab 抗原结合位点,它们可以与两个独立的病原体细胞结合,将它们聚集在一起。 当涉及多个 IgG 抗体时,可能会形成大型聚集体;这些聚集体更容易让肾脏和脾脏从血液中过滤出来,吞噬细胞更容易摄入以进行破坏。 IgM 的五聚体结构为每个分子提供十个 Fab 结合位点,使其成为最有效的凝集抗体。
抗体的另一个重要功能是激活补体级联。 如前一章所述,补体系统是先天防御的重要组成部分,它促进炎症反应,将吞噬细胞招募到感染部位,通过 opsonization 增强吞噬作用,以及使用膜攻击复合物 (MAC) 杀死革兰氏阴性细菌病原体)。 补体激活可以通过三种不同的途径发生(见图17.2.2),但最有效的是经典途径,它要求IgG或IgM抗体与病原体细胞表面进行初始结合,从而允许C1复合物的招募和激活。
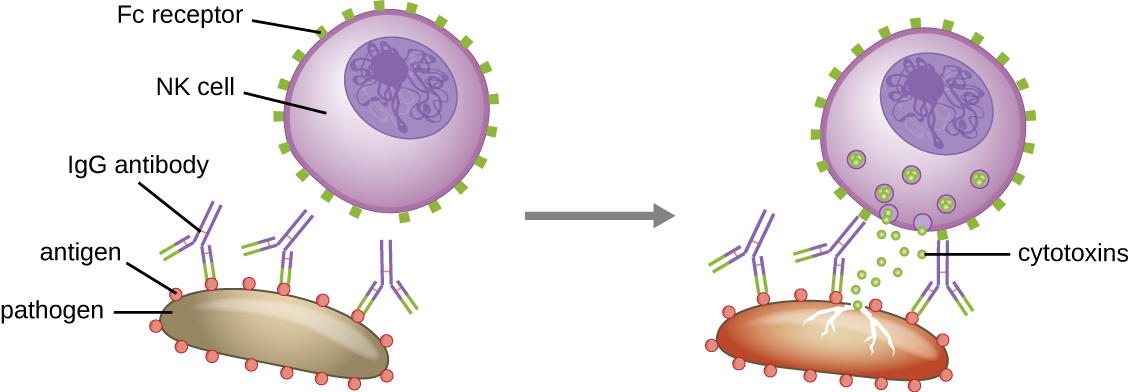
抗体的另一个重要功能是抗体依赖性细胞介导的细胞毒性(ADCC),它可以增强对体积过大而无法吞噬的病原体的杀灭作用。 如图所示,这个过程最能表征自然杀伤细胞(NK 细胞)\(\PageIndex{9}\),但它也可能涉及巨噬细胞和嗜酸性粒细胞。 当 IgG 抗体的 Fab 区域与大型病原体结合时,就会发生 ADCC;效应细胞(例如 NK 细胞)上的 Fc 受体随后与抗体的 Fc 区域结合,使它们与靶病原体非常接近。 然后,效应细胞分泌强大的细胞毒素(例如穿孔素和颗粒酶),从而杀死病原体。
练习\(\PageIndex{6}\)
- IgA 通常在哪里找到?
- 哪类抗体穿过胎盘,为胎儿提供保护?
- 比较 opsonization 和抗体依赖性细胞介导的细胞毒性的机制。
关键概念和摘要
- 适应性免疫是对外来病原体的获得性防御,其特征是特异性和记忆力。 第一次接触抗原会刺激初级反应,随后的暴露会刺激更快、更强的二次反应。
- 适应性免疫是一种双重系统,涉及体液免疫(B 细胞产生的抗体)和细胞免疫(针对细胞内病原体的 T 细胞)。
- 抗原,也称为免疫原,是激活适应性免疫的分子。 单个抗原具有较小的表位,每种抗原都能诱导特定的适应性免疫反应。
- 抗原刺激免疫反应的能力取决于多种因素,包括其分子类别、分子复杂性和大小。
- 抗体(免疫球蛋白)是 Y 形糖蛋白,有两个用于结合抗原的 Fab 位点和一个 Fc 部分参与补体激活和超声化。
- 五类抗体是 IgM、I g G、IgA、IgE 和 IgD,每类抗体的大小、排列、在体内的位置和功能各不相同。 抗体的五个主要功能是中和、选音、凝集、补体激活和抗体依赖性细胞介导的细胞毒性 (ADCC)。