
10.4: 细胞基因组的结构和功能
- Page ID
- 200095

学习目标
- 定义基因和基因型并将基因型与表型区分开来
- 描述染色体结构和包装
- 比较原核和真核染色体
- 解释为什么染色体外的 DNA 在细胞中很重要
到目前为止,我们已经讨论了单个DNA和RNA片段的结构和功能。 在本节中,我们将讨论生物体的所有遗传物质(统称为其基因组)是如何在细胞内部组织的。 由于生物体的遗传学在很大程度上决定了其特征,因此生物的DNA和RNA排列的不同也就不足为奇了。
基因型与表型
所有细胞活动都编码在细胞的 DNA 中。 DNA 分子中的碱基序列代表细胞的遗传信息。 DNA 分子片段被称为基因,单个基因包含合成各种蛋白质、酶或稳定 RNA 分子所需的教学代码。
细胞在其基因组中包含的全部基因集合称为其基因型。 但是,细胞不能同时表达其所有基因。 相反,它会在必要时开启(表达)或关闭某些基因。 在任何给定时间点表达的基因组决定了细胞的活性及其可观察的特征,称为其表型。 经常表达的基因被称为组成基因;一些组成基因被称为内务基因,因为它们是细胞基本功能所必需的。
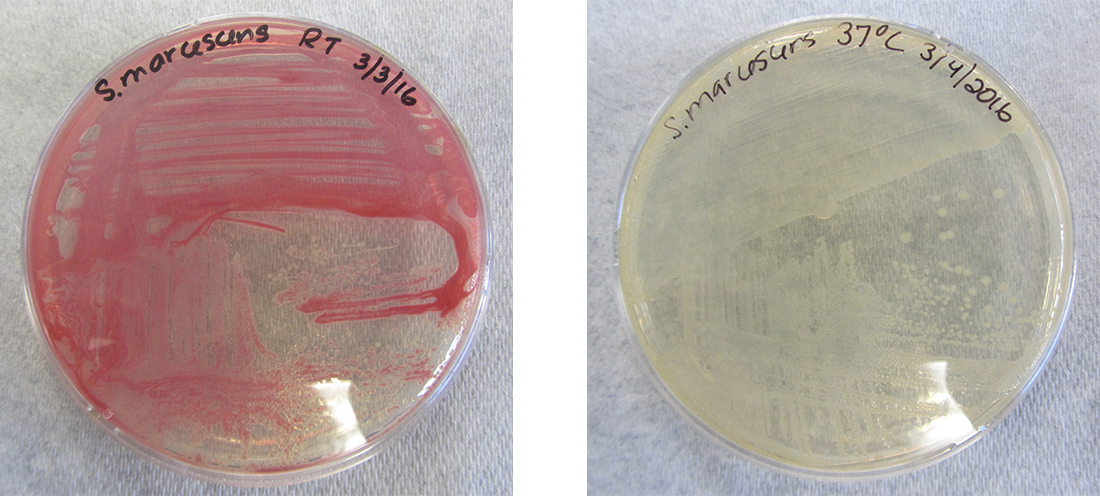
虽然细胞的基因型保持不变,但表型可能会随着环境信号(例如温度或营养可用性的变化)而发生变化,这些信号会影响哪些非组成基因的表达。 例如,口腔细菌 Streptoccus mutans 会产生粘稠的粘液层,使其能够粘附在牙齿上,形成牙菌斑;但是,控制粘液层产生的基因只有在蔗糖(食用糖)存在的情况下才能表达。 因此,尽管变异杆菌的基因型是恒定的,但其表型会根据环境中糖的存在与否而变化。 温度还可以调节基因表达。 例如,革兰氏阴性细菌 Serratia marcescens 是一种经常与医院获得性感染相关的病原体,在 28 °C 时会产生红色色素,但在 37 °C(人体正常内部温度)时不会产生红色色素(图\(\PageIndex{1}\))。
遗传物质的组织
生物体的绝大多数基因组都组织成细胞的染色体,染色体是细胞内控制细胞活动的离散DNA结构。 回想一下,虽然真核生物染色体存放在膜结合的细胞核中,但大多数原核生物都含有单个圆形染色体,该染色体存在于细胞质的一个称为核样体的区域中(参见原核细胞的独特特征)。 一条染色体可能包含数千个基因。
真核染色体的组织
真核细胞和原核细胞的染色体结构略有不同。 真核生物染色体通常是线性的,真核细胞包含多个不同的染色体。 许多真核细胞包含每条染色体的两个副本,因此是二倍体。
染色体的长度大大超过了细胞的长度,因此需要将染色体封装到一个非常小的空间中才能容纳在细胞内。 例如,如果完全伸展,人类基因组中所有30亿个碱基对 1 的总长度将达到大约2米,而一些真核生物基因组比人类基因组大许多倍。 DNA supercoiling 是指 DNA 被扭曲以适应细胞内的过程。 Supercoiling 可能导致 DNA 从其正常的松弛状态下弯曲(每 10 个碱基对中螺旋线少于一圈),要么超出(每 10 个碱基对超过一圈)。 已知参与超线圈的蛋白质包括拓扑异构酶;这些酶有助于维持超线圈染色体的结构,防止DNA在某些细胞过程(例如DNA复制)中过度卷曲。
在 DNA 包装过程中,称为组蛋白的 DNA 结合蛋白进行不同程度的 DNA 包裹和附着在支架蛋白上。 DNA与这些附着蛋白的组合被称为染色质。 在真核生物中,组蛋白对DNA的包装可能受到环境因素的影响,这些环境因素会影响DNA的某些胞嘧啶核苷酸上存在甲基。 环境因素对DNA封装的影响称为表观遗传学。 表观遗传学是在不改变核苷酸序列的情况下调节基因表达的另一种机制。 表观遗传学变化可以通过多轮细胞分裂来维持,因此可以遗传。
观看来自 DNA 学习中心的动画,了解有关真核生物中 DNA 包装的更多信息。
原核染色体的组织
细菌和古细菌中的染色体通常是圆形的,而原核细胞在核样体中通常只包含一条染色体。 由于染色体仅包含每个基因的一个拷贝,因此原核生物是单倍体。 与真核细胞一样,DNA超级线圈是基因组融入原核细胞的必要条件。 细菌染色体中的DNA排列在几个超线圈结构域中。 与真核生物一样,拓扑异构酶参与超线圈 DNA。 DNA gyrase 是一种拓扑异构酶,存在于细菌和某些古细菌中,有助于防止 DNA 的过载。 (有些抗生素通过靶向 DNA 回旋酶来杀死细菌。) 此外,组蛋白样蛋白结合 DNA 并帮助 DNA 包装。 其他蛋白质与复制起源(DNA 复制在染色体中的起始位置)结合。 由于 DNA 的不同区域包装不同,因此酶更容易获得染色体 DNA 的某些区域,因此可能更容易用作基因表达的模板。 有趣的是,包括幽门螺杆菌和弗莱克斯内里志贺氏杆菌在内的几种细菌已被证明会在感染时诱发宿主的表观遗传学变化,从而导致染色质重塑,可能对宿主免疫造成长期影响。 2
练习\(\PageIndex{1}\)
- 细胞的基因型和表型有什么区别?
- DNA 如何融入细胞?
非编码 DNA
除基因外,基因组还包含许多非编码 DNA 区域,这些区域不编码蛋白质或稳定 RNA 产物。 非编码 DNA 通常存在于基因序列编码开始之前的区域以及基因间区域(即位于基因之间的 DNA 序列)(图\(\PageIndex{2}\))。
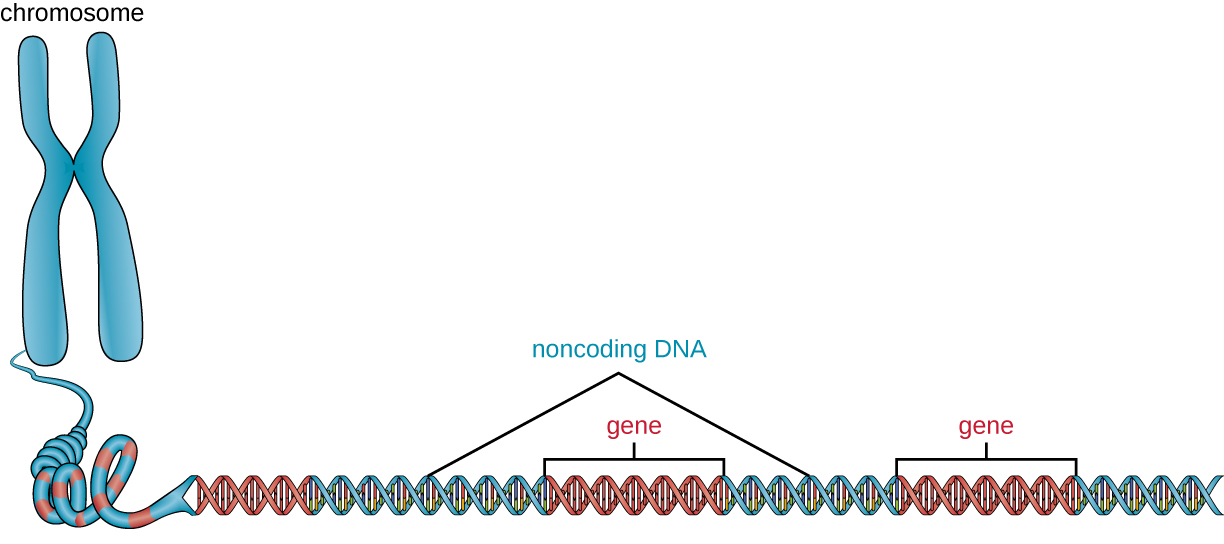
原核生物似乎非常有效地使用其基因组,平均只有12%的基因组被非编码序列占用。 相比之下,非编码DNA可以占真核生物基因组的98%左右,如人类所见,但非编码DNA的百分比因物种而异。 3 这些非编码 DNA 区域曾经被称为 “垃圾 DNA”;但是,这个术语已不再被广泛接受,因为科学家此后发现了其中一些区域的作用,其中许多区域通过产生小的 DNA 有助于调节转录或翻译非编码 RNA 分子、DNA 封装和染色体稳定性。 尽管科学家可能不完全了解 DNA 所有非编码区域的作用,但人们普遍认为它们在细胞内确实有用途。
练习\(\PageIndex{2}\)
非编码 DNA 的作用是什么?
染色体外的 DNA
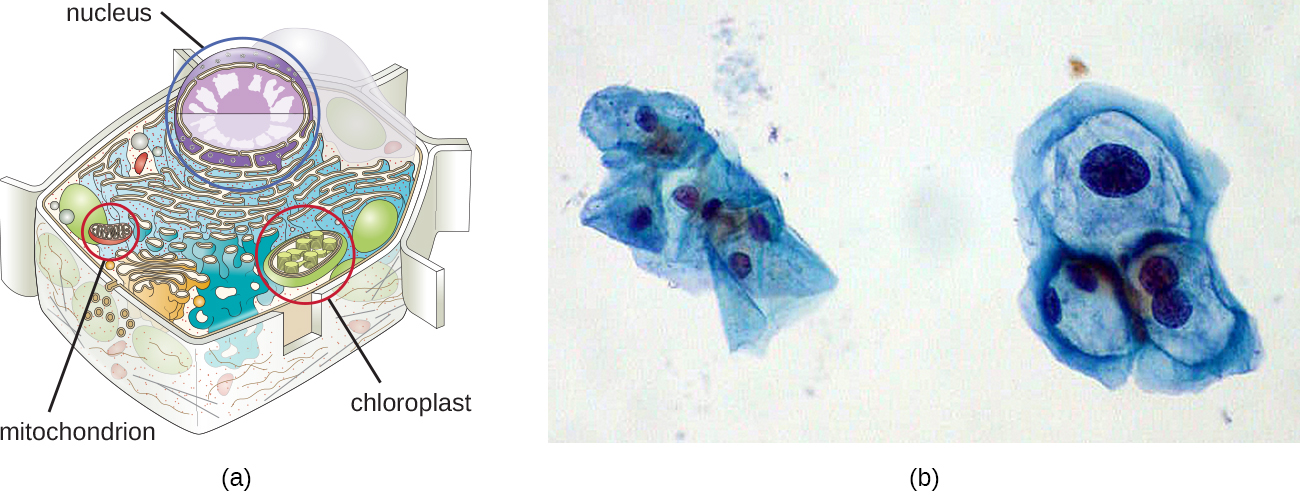
尽管大多数DNA都包含在细胞的染色体中,但许多细胞在染色体之外还有额外的DNA分子,称为染色体外DNA,它们也是其基因组的一部分。 真核细胞的基因组还将包括来自这些细胞维持的任何细胞器的染色体,例如线粒体和/或叶绿体(图\(\PageIndex{3}\))。 维持这些细胞器中的圆形染色体是其原核生物起源的痕迹,并支持内共生理论(参见《现代细胞理论基础》)。 在某些情况下,在潜伏病毒感染期间,某些DNA病毒的基因组也可以在宿主细胞中独立维持。 在这些情况下,这些病毒是另一种形式的染色体外DNA。 例如,人乳头瘤病毒(HPV)可以通过这种方式维持在受感染的细胞中。
除了染色体外,一些原核生物还有较小的DNA环,称为质粒,其中可能包含一个或几个对正常生长不重要的基因(图3.3.1)。 细菌可以在称为水平基因转移(HGT)的过程中与其他细菌交换这些质粒。 质粒上遗传物质的交换有时会为微生物提供有利于特殊条件下生长和存活的新基因。 在某些情况下,从质粒中获得的基因可能具有临床意义,它们编码的毒性因子使微生物能够引起疾病或使微生物对某些抗生素产生耐药性。 质粒还大量用于基因工程和生物技术,作为将基因从一个细胞转移到另一个细胞的一种方式。 质粒在水平基因转移和生物技术中的作用将在《微生物遗传学机制和微生物遗传学的现代应用》中进一步讨论。
练习\(\PageIndex{3}\)
质粒如何参与抗生素耐药性?
致命质粒
玛丽亚是一名来自德克萨斯州的20岁人类学专业学生,最近在非洲国家博茨瓦纳生病,她正在那里作为海外留学计划的一部分进行研究。 玛丽亚的研究重点是传统的非洲制革皮革生产方法。 在三周的时间里,她每天到一家制革厂参观几个小时,观察和参与制革过程。 有一天,玛丽亚从制革厂回来后,出现了发烧、发冷和头痛,还有胸痛、肌肉酸痛、恶心和其他类似流感的症状。 起初,她并不担心,但是当她的发烧激增并开始咳血时,她的非洲寄宿家庭惊慌失措,将她送往医院,她的病情继续恶化。
在得知她最近在制革厂工作后,医生怀疑玛丽亚曾接触过炭疽病。 他下令做胸部 X 光检查、血液样本和脊柱抽头,然后立即开始静脉注射青霉素疗程。 不幸的是,实验室检查证实了医生的推定诊断。 玛丽亚的胸部 X 光检查显示胸腔积液、胸膜之间空间积液以及血液中的革兰染色显示短链中存在革兰氏阳性的棒状细菌,这与炭疽芽孢杆菌一致。 血液和细菌也被证明存在于她的脑脊液中,这表明感染已发展为脑膜炎。 尽管进行了支持性治疗和积极的抗生素治疗,但玛丽亚还是进入了无反应状态,并在三天后死亡。
炭疽是一种由革兰氏阳性细菌炭疽杆菌的内孢子引入体内引起的疾病。 一旦感染,患者通常会患上脑膜炎,通常会导致致命的后果。 就玛丽亚而言,她在处理被感染动物的皮革时吸入了内孢子。
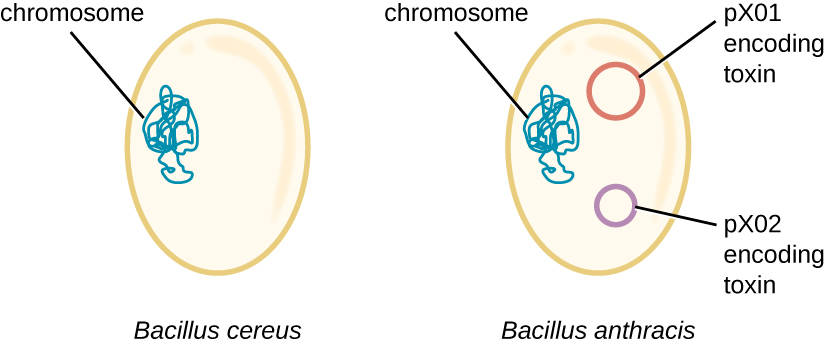
炭疽杆菌的基因组说明了微小的结构差异会导致毒力的重大差异。 2003年,对炭疽杆菌和蜡样芽孢杆菌(一种相同属的相似但致病性较小的细菌)的基因组进行了测序和比较。 4 研究人员发现,这些细菌的16S rRNA基因序列的相同性超过99%,这意味着尽管它们传统上被归类为单独的物种,但它们实际上是同一个物种的成员。 尽管它们的染色体序列也显示出很大的相似性,但发现炭疽杆菌的几种毒力因子编码在蜡样芽孢杆菌中找不到的两个大质粒上。 质粒 px01 编码一种抑制宿主免疫系统的三部分毒素,而质粒 px02 编码一种囊多糖,进一步保护细菌免受宿主免疫系统的侵害(图\(\PageIndex{4}\))。 由于蜡样芽孢杆菌缺乏这些质粒,因此它不会产生这些毒力因子,尽管它仍然具有致病性,但它通常与人体可以迅速从中恢复的轻度腹泻病例有关。 对玛丽亚来说,不幸的是,炭疽杆菌中存在这些编码毒素的质粒使其具有致命的毒性。
练习\(\PageIndex{4}\)
如果炭疽杆菌丢失了一个或两个质粒,你认为它的致病性会怎样?
临床重点:分辨率
在24小时内,对Alex粪便样本的诊断测试分析结果显示,该样本的耐热肠毒素(LT)、热稳定肠毒素(ST)和定植因子(CF)呈阳性,证实了医院医生对ETEC的怀疑。 在对亚历克斯的家庭医生进行随访期间,这位医生指出,亚历克斯的症状没有很快消失,他感到不适,使他无法重返课堂。 家庭医生给亚历克斯开了一个疗程的环丙沙星来缓解他的症状。 幸运的是,环丙沙星在几天之内就解决了亚历克斯的症状。
亚历克斯很可能是由于摄入受污染的食物或水而感染的。 像墨西哥这样的新兴工业化国家仍在制定卫生措施,防止水被粪便污染。 这些国家的旅行者应避免摄入未煮熟的食物,尤其是肉类、海鲜、蔬菜和未经巴氏消毒的乳制品。 他们还应避免使用未经处理的水;这包括饮用水、冰块,甚至是用于刷牙的水。 将瓶装水用于这些目的是一个不错的选择。 良好的卫生习惯(洗手)也有助于预防ETEC感染。 亚历克斯对自己的食物或水消耗不够谨慎,这导致了他生病。
亚历克斯的症状与霍乱的症状非常相似,霍乱是由革兰氏阴性细菌霍乱弧菌引起的,霍乱还会产生类似于ST和LT的毒素。 在 ETEC 进化史的某个时刻,一种与肠道中常见的大肠杆菌相似的非致病性大肠杆菌菌株可能已经从霍乱杆菌中获得了编码 ST 和 LT 毒素的基因。 编码这些毒素的基因是在 ETEC 中的染色体外质粒上编码的,这一事实支持了这样的观点,即这些基因是由大肠杆菌获得的,很可能通过水平基因转移在细菌群体中保存。
病毒基因组
病毒基因组在结构上表现出明显的多样性。 有些病毒的基因组由DNA作为其遗传物质组成。 这种DNA可能是单链的,以人类细小病毒为例,也可以是双链的,如疱疹病毒和痘病毒所示。 此外,尽管所有细胞生命都使用DNA作为其遗传物质,但正如我们已经讨论过的那样,一些病毒基因组是由单链或双链RNA分子组成的。 病毒基因组通常比大多数细菌基因组小,仅编码少数基因,因为它们依赖宿主来执行复制所需的许多功能。 《病毒生命周期》更详细地讨论了病毒基因组结构的多样性及其对病毒复制生命周期的影响。
练习\(\PageIndex{5}\)
为什么病毒基因组在病毒之间差异很大?
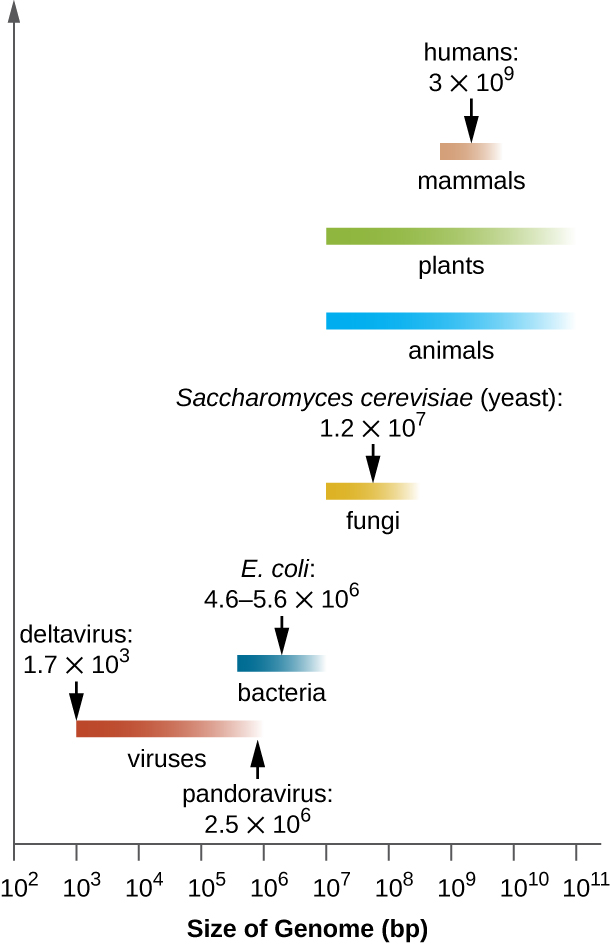
基因组大小很重要
不同生物之间的基因组大小差异很大。 大多数真核生物维持多条染色体;例如,人类有23对染色体,产生46条染色体。 尽管人类基因组有30亿个碱基对,但远非最大的基因组。 植物通常维持着非常大的基因组,多达1500亿个碱基对,通常是多倍体,每条染色体都有多个拷贝。
细菌基因组的大小也差异很大,尽管它们往往比真核生物基因组小(图\(\PageIndex{5}\))。 一些细菌基因组可能只有11.2万个碱基对。 通常,细菌基因组的大小与细菌的生存依赖程度直接相关。 当细菌依靠宿主细胞执行某些功能时,它就会失去编码自己执行这些功能的能力的基因。 这些类型的细菌 endosymbionts 让人想起线粒体和叶绿体的原核起源。
从临床角度来看,专性细胞内病原体也往往具有较小的基因组(大约100万个碱基对)。 由于宿主细胞提供大部分营养,因此编码代谢功能的基因数量往往会减少。 由于体积小,生殖器支原体(58万个碱基对)、沙眼衣原体(100万个)、普罗瓦泽基立克氏菌(110万个)和梅毒螺旋体(110万个)等生物的基因组是早期的细菌基因组中的一部分按顺序排列。 这些病原体分别引起尿道炎和盆腔炎症、衣原体、斑疹伤寒和梅毒。
尽管专性细胞内病原体的基因组异常小,但其他具有多种代谢和酶促能力的细菌基因组却异常大。 例如,铜绿假单胞菌是一种常见于环境中的细菌,能够在各种基质上生长。 它的基因组包含630万个碱基对,使其具有很高的代谢能力和产生导致多种机会性感染的毒性因子的能力。
有趣的是,病毒的基因组大小也存在显著差异,从3500个碱基对到250万个碱基对不等,大大超过了许多细菌基因组的大小。 观察到的病毒基因组大小的巨大差异进一步促成了已经讨论过的病毒基因组特征的巨大多样性。
访问国家生物技术信息中心 (NCBI) 的基因组数据库,查看已测序的基因组及其大小。
关键概念和摘要
- 细胞的全部遗传成分是其基因组。
- 基因编码蛋白质或稳定的RNA分子,每种分子在细胞中都具有特定的功能。
- 尽管细胞拥有的基因型保持不变,但基因的表达取决于环境条件。
- 表型是细胞(或生物体)在给定时间点的可观察特征,由目前使用的基因补体产生。
- 大多数遗传物质被组织成包含控制细胞活动的 DNA 的染色体。
- 原核生物通常是单倍体,通常在核样体中发现单个圆形染色体。 真核生物是二倍体;DNA 被组织成在细胞核中发现的多个线性染色体。
- 使用DNA结合蛋白进行超级线圈和DNA封装可以使长分子装入细胞内。 真核生物和古细菌使用组蛋白蛋白,细菌使用具有相似功能的不同蛋白质。
- 原核生物和真核生物基因组都含有非编码 DNA,其功能尚不清楚。 一些非编码 DNA 似乎参与了影响基因表达的小型非编码 RNA 分子的形成;有些似乎在维持染色体结构和 DNA 包装方面发挥了作用。
- 真核生物中的 Extrachromosomal DNA 包括在由内共生体进化而来的原核细胞器(线粒体和叶绿体)中发现的染色体。 有些病毒也可能在染色体外维持自身。
- 原核生物中的染色体外DNA通常以质粒的形式保存,它们编码一些在特定条件下可能有用的非必需基因。 质粒可以通过水平基因转移在细菌群落中传播。
- 病毒基因组显示出广泛的变异,可能由RNA或DNA组成,可以是双链或单链。
脚注
- 1 国家人类基因组研究所。 “人类基因组计划完成:常见问题。” https://www.genome.gov/11006943。 已于 2016 年 6 月 10 日访问
- 2 H. Bierne 等人。 “表观遗传学和细菌感染。” 《冷泉港医学展望》第 2 期第 12 期 (2012): a010272。
- 3 R.J. Taft 等人 “非蛋白质编码DNA与真核生物复杂性之间的关系。” Bioessays 29 第 3 期 (2007): 288—299。
- 4 N. Ivanova 等人 “蜡样芽孢杆菌的基因组序列和与炭疽芽孢杆菌的比较分析。” 《自然》423 编号 6935 (2003): 87—91。