
38.4: 肌肉收缩和运动
- Page ID
- 202375

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 对不同类型的肌肉组织进行分类
- 解释肌肉在运动中的作用
肌肉细胞专门用于收缩。 肌肉允许诸如行走之类的运动,它们还可以促进呼吸和消化等身体过程。 人体包含三种类型的肌肉组织:骨骼肌、心肌和平滑肌(图\(\PageIndex{1}\))。
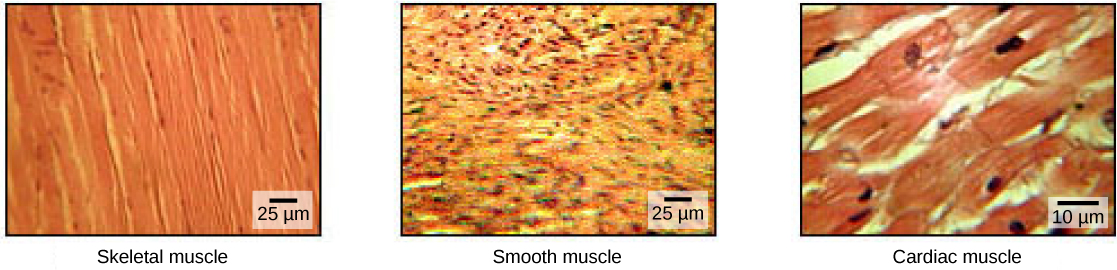
骨骼肌组织形成骨骼肌,骨骼肌附着在骨骼或皮肤上,控制运动和任何可以有意识控制的运动。 因为它可以通过思想来控制,所以骨骼肌也被称为自愿肌肉。 骨骼肌外观长而呈圆柱形;在显微镜下观察时,骨骼肌组织呈条纹或条纹外观。 条纹是由收缩蛋白(肌动蛋白和肌球蛋白)的规则排列引起的。 肌动蛋白是一种球状收缩蛋白,与肌球蛋白相互作用以收缩肌肉。 骨骼肌在单个细胞中还存在多个核。
平滑肌组织存在于肠道、胃和膀胱等空心器官的壁上,以及呼吸道和血管等通道周围。 平滑肌没有条纹,不受自愿控制,每个细胞只有一个核,两端呈锥形,被称为非自愿肌肉。
心肌组织仅存在于心脏中,心脏收缩会将血液泵送到全身并维持血压。 与骨骼肌一样,心肌是横纹肌的,但与骨骼肌不同,心肌无法被有意识地控制,被称为非自愿肌肉。 它每个细胞有一个核,是分支的,其特点是存在嵌合的圆盘。
骨骼肌纤维结构
每根骨骼肌纤维都是骨骼肌细胞。 这些电池非常大,直径可达 100 µm,长度可达 30 cm。 骨骼肌纤维的质膜被称为 s arcolemma。 肌瘤是动作电位传导的部位,它会触发肌肉收缩。 每根肌肉纤维中都有 myofibrils,即与肌肉纤维平行的长圆柱结构。 肌纤维贯穿肌肉纤维的整个长度,由于它们的直径只有大约 1.2 微米,因此在一根肌肉纤维中可以找到数百到数千个。 它们的末端附着在肌瘤上,因此随着肌原纤维的缩短,整个肌肉细胞会收缩(图\(\PageIndex{2}\))。
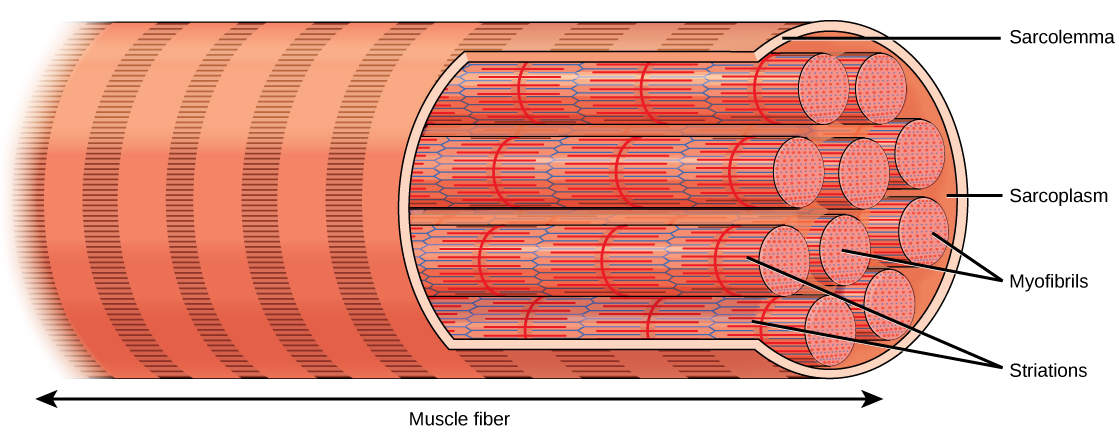
骨骼肌组织的条纹外观是肌原纤维长度上存在的肌动蛋白和肌球蛋白重复带的结果。 Dark A bands 和 light I 带沿着肌原纤维重复,细胞中肌原纤维的对齐会导致整个细胞呈条纹或带状状。
每个 I 波段都有一条垂直穿过中间的密集线,称为 Z 盘或 Z 线。 Z 盘标志着被称为 sarcomeres 的单位的边界,这些单位是骨骼肌的功能单位。 一个 sarcomere 是两个连续的 Z 光盘之间的空间,包含整个 A 波段和 I 波段的两半,一个 A 波段的两侧。 myofibril 由沿其长度延伸的许多肉瘤组成,当肉瘤单独收缩时,肌原纤维和肌肉细胞会缩短(图\(\PageIndex{3}\))。
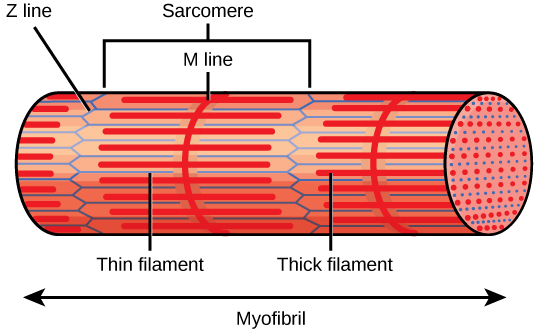
Myofibr ils 由称为肌丝的较小结构组成。 细丝主要有两种类型:粗丝和细丝;每种都有不同的成分和位置。 粗细丝仅出现在肌原纤维的 A 带中。 细丝附着在 Z 盘中一种叫做 alpha-actinin 的蛋白质上,存在于 I 带的整个长度和 A 带的中途。 粗丝和细丝重叠的区域具有密集的外观,因为细丝之间的空间很小。 细丝不会一直延伸到 A 波段中,留下 A 带的中心区域仅包含粗细丝。 A 波段的这个中心区域看起来比 A 波段的其余部分稍轻,被称为 H 区。 H 区的中间有一条叫做 M 线的垂直线,辅助蛋白在这条线上将粗细的细丝聚集在一起。 Z 盘和 M 线都将肌丝固定在适当的位置,以保持肌原纤维的结构排列和分层。 Myofibrils 通过附着在 Z 盘上的中间细丝或 desmin 细丝相互连接。
粗丝和细丝本身由蛋白质组成。 粗丝由蛋白质肌球蛋白组成。 肌球蛋白分子的尾部与其他肌球蛋白分子相连,在 M 线附近形成一根粗丝的中心区域,而头部对齐在细丝重叠的粗丝的两侧。 细丝的主要成分是肌动蛋白。 细丝的另外两个成分是原肌球蛋白和肌钙蛋白。 肌动蛋白具有肌球蛋白附着的结合位点。 当肌肉处于静止状态时,原肌球蛋白链会阻断结合位点并防止肌动蛋白与肌球蛋白的相互作用。 肌钙蛋白由三个球状亚基组成。 一个亚基与原肌球蛋白结合,一个亚基与肌动蛋白结合,一个亚基结合 Ca 2+ 离子。
链接到学习
观看这个展示肌肉纤维组织结构的动画。
滑丝收缩模型
要使肌肉细胞收缩,肉瘤必须缩短。 但是,粗丝和细丝(肉瘤的成分)不会缩短。 相反,它们互相滑动,导致肉瘤缩短,而细丝长度保持不变。 肌肉收缩的滑丝理论的开发是为了适应在不同程度的肌肉收缩和松弛下肉瘤上观察到的命名带中观察到的差异。 收缩机制是肌球蛋白与肌动蛋白结合,形成交叉桥,产生细丝运动(图\(\PageIndex{4}\))。
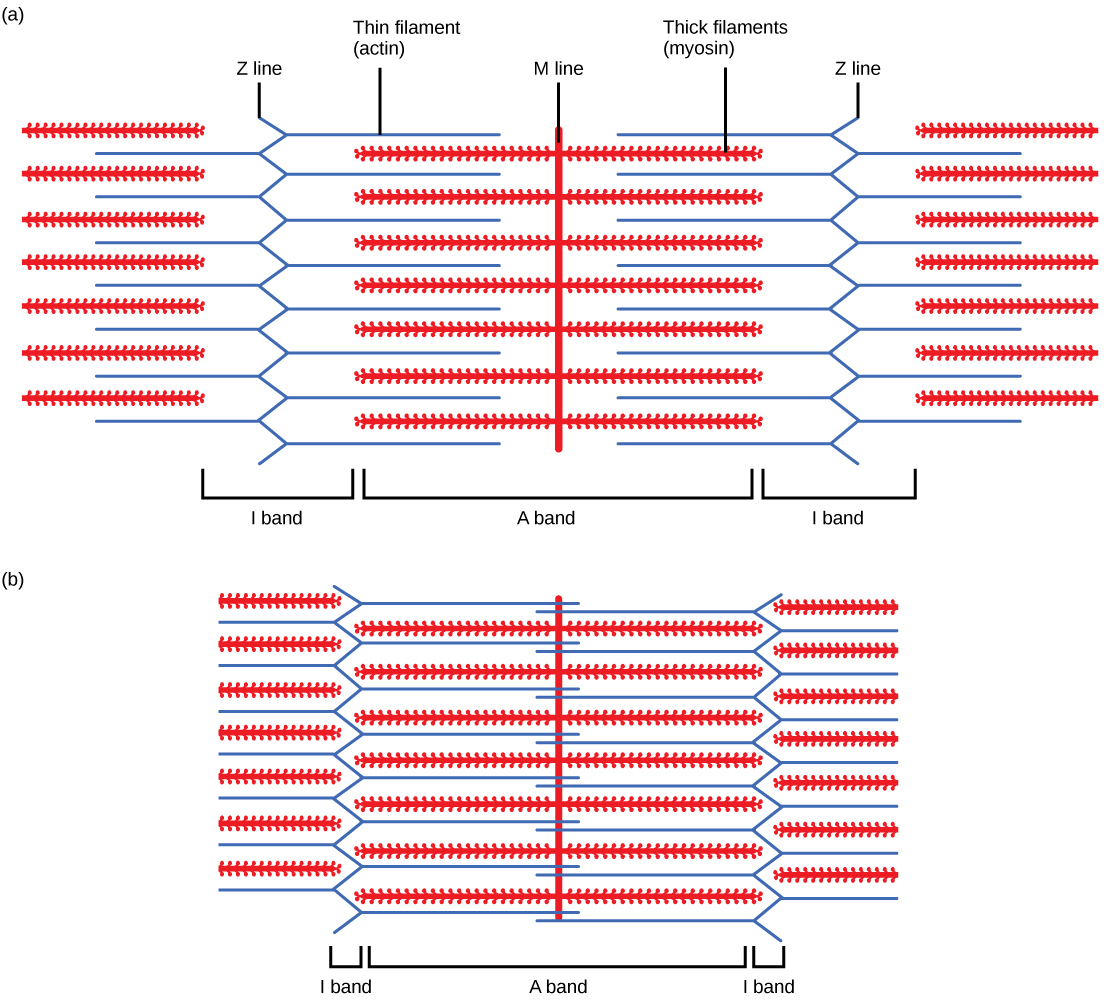
当肉瘤缩短时,一些区域会缩短,而另一些区域则保持不变。 肉瘤定义为两个连续的 Z 盘或 Z 线之间的距离;当肌肉收缩时,Z 盘之间的距离会缩小。 H 区(A 区的中心区域)仅包含粗丝,在收缩时会缩短。 I 波段仅包含细丝,还能缩短。 A 波段不会缩短 —— 长度保持不变 —— 但是 A 带在收缩过程中会靠得更近,最终消失。 细丝被粗细的细丝拉向肉瘤的中心,直到 Z 盘接近粗细的细丝。 重叠区域随着细丝向内移动而增加,其中细丝和粗细丝占据相同的面积。
ATP 和肌肉收缩
肌肉缩短的运动发生在肌球蛋白头部与肌动蛋白结合并将肌动蛋白向内拉动时。 此动作需要能量,由 ATP 提供。 肌球蛋白在球状肌动蛋白的结合位点与肌动蛋白结合。 肌球蛋白还有另一个与ATP结合的位点,在该位点的酶活性将ATP水解为ADP,从而释放出无机磷酸盐分子和能量。
ATP 结合导致肌球蛋白释放肌动蛋白,使肌动蛋白和肌球蛋白相互分离。 发生这种情况之后,新结合的 ATP 会转化为 ADP 和无机磷酸盐 P i。 肌球蛋白结合位点的酶被称为 ATPase。 ATP 水解过程中释放的能量将肌球蛋白头部的角度变为 “竖起” 位置。 然后,肌球蛋白头可以进一步移动,拥有潜在能量,但是 ADP 和 P i 仍然附着。 如果肌动蛋白结合位点被覆盖且不可用,则肌球蛋白将在ATP水解后保持高能量配置,但仍会附着。
如果肌动蛋白结合位点被发现,就会形成交叉桥;也就是说,肌球蛋白头跨越肌动蛋白和肌球蛋白分子之间的距离。 然后释放 P i,允许肌球蛋白消耗储存的能量作为构象变化。 肌球蛋白头向 M 线移动,同时拉动肌动蛋白。 当肌动蛋白被拉动时,细丝向 M 线移动大约 10 nm。 这种运动被称为强力冲程,因为它是产生力的步骤。 当肌动蛋白向 M 线拉动时,肉瘤会缩短,肌肉收缩。
当肌球蛋白头 “竖起” 时,它含有能量并且处于高能量配置。 这种能量是在肌球蛋白头部通过强力冲程时消耗的;在强力冲程结束时,肌球蛋白头处于低能量位置。 强力冲击后,ADP 被释放;但是,形成的交叉桥仍然存在,肌动蛋白和肌球蛋白结合在一起。 然后 ATP 可以附着在肌球蛋白上,这允许跨桥周期重新开始,肌肉可能会进一步收缩(图\(\PageIndex{5}\))。
链接到学习
观看这段视频,解释肌肉收缩是如何发出信号的。
艺术连接
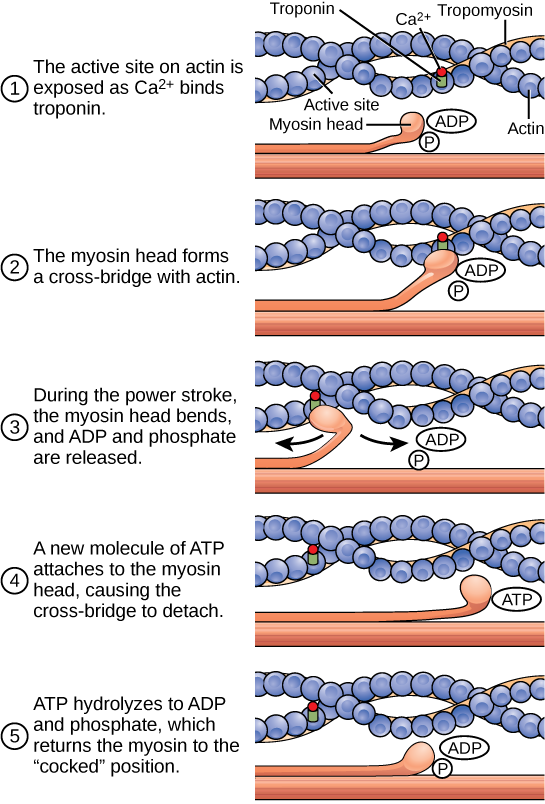
以下关于肌肉收缩的陈述中哪一项是正确的?
- 当 ATP 水解成 ADP 和磷酸盐时,就会发生强力冲击。
- 当 ADP 和磷酸盐与肌球蛋白头分离时,就会发生强力冲击。
- 当 ADP 和磷酸盐与肌动蛋白活性位点分离时,就会发生强力中风。
- 当 Ca 2+ 结合钙头时,就会发生强力冲击。
链接到学习
观看这个跨桥肌肉收缩的动画。
调节蛋白
当肌肉处于静止状态时,肌动蛋白和肌球蛋白会分离。 为了防止肌动蛋白与肌球蛋白上的活性位点结合,调节蛋白会阻断分子结合位点。 原@@ 肌球蛋白阻断肌动蛋白分子上的肌球蛋白结合位点,防止交叉桥形成并防止没有神经输入的肌肉收缩。 肌@@ 钙蛋白与原肌球蛋白结合并帮助将其定位在肌动蛋白分子上;它还结合钙离子。
为了实现肌肉收缩,原肌球蛋白必须改变构象,在肌动蛋白分子上发现肌球蛋白结合位点并允许交叉桥形成。 这种情况只有在存在钙的情况下才会发生,钙在肌质中的浓度保持在极低的水平。 如果存在,钙离子会与肌钙蛋白结合,导致肌钙蛋白的构象变化,从而使原肌球蛋白能够远离肌动蛋白上的肌球蛋白结合位点。 一旦原肌球蛋白被移除,肌动蛋白和肌球蛋白之间就会形成交叉桥梁,从而触发收缩。 跨桥循环一直持续到 Ca 2+ 离子和 ATP 不再可用,原肌球蛋白再次覆盖肌动蛋白的结合位点。
激收耦合
兴奋-收缩耦合是肌肉瘤中产生的动作电位与肌肉收缩开始之间的联系(转导)。 钙从肌质网释放到肌质的触发因素是神经信号。 每根骨骼肌纤维都由运动神经元控制,运动神经元将信号从大脑或脊髓传导到肌肉。 肌肉纤维上与神经元相互作用的肌肉瘤区域称为运动端板。 神经元轴突的末端被称为突触末端,它实际上并不接触马达端板。 一个叫做 synaptic cleft 的小空间将突触端子与电机端板分开。 电信号沿着神经元的轴突传播,轴突穿过肌肉并在神经肌肉交界处连接到单个肌肉纤维。
细胞的电通信能力要求细胞消耗能量在细胞膜上产生电梯度。 这种电荷梯度由离子携带,离子在膜上分布差异。 每个离子都会施加电气影响和浓度影响。 正如牛奶最终无需搅拌即可与咖啡混合一样,如果允许的话,离子也会均匀分布。 在这种情况下,不允许它们恢复到均匀混合的状态。
钠-钾 ATPase 利用细胞能量在细胞内移动 K + 离子,将 Na + 离子移到细胞外。 仅此一项就会积累少量电荷,但浓度梯度很大。 细胞中有很多 K +,细胞外有很多 Na +。 钾能够通过 K + 通道离开细胞,这些通道在 90% 的时间都是开放的,而且确实如此。 但是,Na + 通道很少开放,因此 Na + 仍留在细胞外。 当 K + 离开细胞,服从其浓度梯度时,实际上会留下负电荷。 因此,静止时,Na + 进入细胞的浓度梯度很大,细胞中会积累大量的负电荷。 这是静息膜电位。 在这种情况下,潜力意味着分离能够起作用的电荷。 它以伏特为单位测量,就像电池一样。 但是,跨膜电位要小得多(0.07 V);因此,小值以毫伏(mV)或 70 mV 表示。 由于电池内部与外部相比为负,因此负号表示电池内部的负电荷过多,−70 mV。
如果一个事件将膜的渗透性改变为钠离子,它们就会进入细胞。 这将改变电压。 这是一种电事件,称为动作电位,可用作蜂窝信号。 神经和肌肉之间通过神经递质进行交流。 神经元动作电位导致神经递质从突触末端释放到突触裂缝,然后它们可以在突触裂缝中扩散并与运动端板上的受体分子结合。 运动端板具有交界折叠,即肉瘤中的折叠,为神经递质与受体结合形成较大的表面积。 受体实际上是钠通道,当它们接收到神经递质信号时,它们会打开以允许 Na + 进入细胞。
乙酰胆碱(acH)是由运动神经元释放的一种神经递质,与运动端板中的受体结合。 当动作电位沿运动神经元轴突向下移动时,就会发生神经递质释放,从而导致突触末端膜的渗透性改变和钙的涌入。 Ca 2+ 离子允许突触囊泡移动到突触前膜(在神经元上)并与之结合,并将神经递质从囊泡释放到突触裂缝中。 一旦被突触末端释放,acH 就会通过突触裂缝扩散到马达端板,在那里它与 acH 受体结合。 当神经递质结合时,这些离子通道打开,Na + 离子穿过膜进入肌肉细胞。 这减少了电池内部和外部之间的电压差,这称为去极化。 当 acH 绑定在电机端板上时,这种去极化被称为端板电位。 然后,去极化沿着肌瘤扩散,在与初始去极化位点相邻的钠通道感知电压的变化并开路时产生动作电位。 动作电位在整个细胞中移动,形成去极化浪潮。
acH 被乙酰胆碱酯酶 (acHE) 酶分解成乙酰和胆碱。 acHe 存在于突触裂缝中,可分解 acH,使其不会与 acH 受体结合,这会导致不必要的肌肉收缩延长(图\(\PageIndex{6}\))。
艺术连接
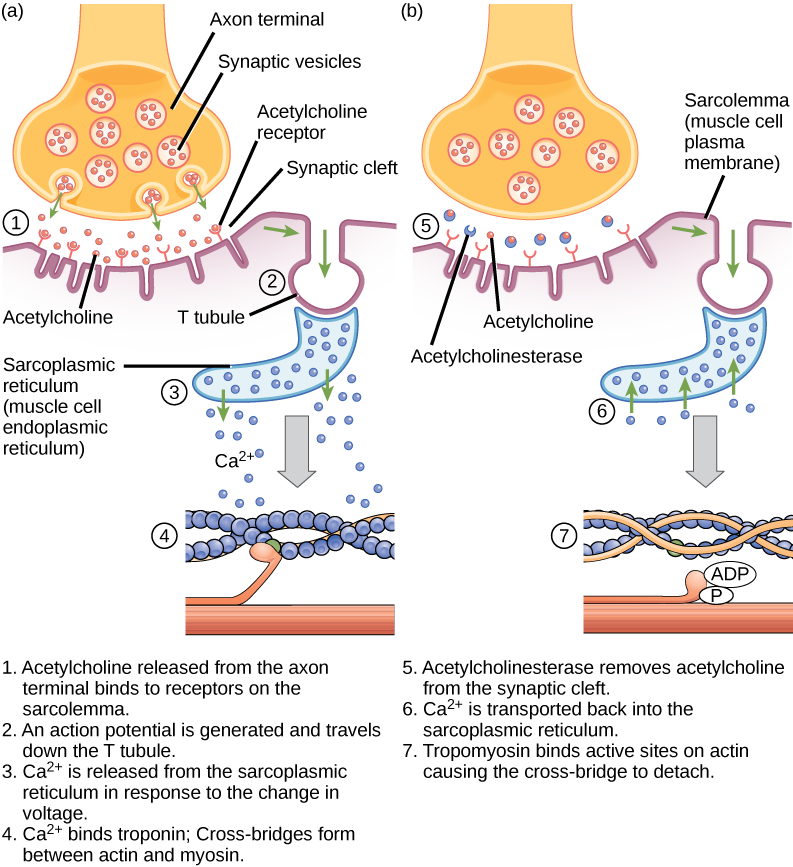
致命的神经毒气沙林不可逆转地抑制乙酰胆碱酯酶。 沙林会对肌肉收缩产生什么影响?
去极化后,膜恢复到静止状态。 这称为复极化,在此期间,电压门控钠通道关闭。 钾通道继续保持在 90% 的电导率。 由于质膜钠-钾 ATPase 总是输送离子,因此静止状态(内部相对于外部带负电荷)得以恢复。 在神经或肌肉中传递冲动之后的时期,即神经元或肌肉细胞恢复传递另一种冲动的能力,称为难治期。 在耐火期,膜无法产生另一种作用电位。 耐火期允许对电压敏感的离子通道恢复到静止状态。 钠钾 ATPase 不断将 Na + 移回细胞,然后 K + 移回细胞,然后 K + 泄漏出来,留下负电荷。 膜很快就会复极化,因此可以再次去极化。
控制肌肉紧张
神经控制启动肌动蛋白—肌球蛋白交叉桥的形成,导致肌肉收缩所涉及的肉瘤缩短。 这些收缩从肌肉纤维延伸到结缔组织,拉动骨骼,导致骨骼运动。 肌肉施加的拉力称为张力,这种张力产生的力量可能会有所不同。 这使得同样的肌肉能够移动非常轻的物体和非常重的物体。 在单个肌肉纤维中,产生的张力取决于肌肉纤维的横截面积和神经刺激的频率。
肌动蛋白和肌球蛋白之间形成的交叉桥的数量决定了肌肉纤维可以产生的张力。 交叉桥只能在粗丝和细丝重叠的地方形成,从而允许肌球蛋白与肌动蛋白结合。 如果形成更多的交叉桥,更多的肌球蛋白会拉动肌动蛋白,从而产生更大的张力。
在产生最大张力期间,肉瘤的理想长度发生在粗丝和细丝最大程度重叠时。 如果静止的肉瘤被拉伸到理想的静止长度以上,则粗细丝不会在最大程度上重叠,可以形成的交叉桥也更少。 这样可以减少肌球蛋白头部拉动肌动蛋白,减少产生的张力。 随着肉瘤的缩短,当细丝到达由肌球蛋白尾巴组成的 H 区时,重叠区域就会缩小。 因为肌球蛋白头形成交叉桥,所以肌动蛋白不会与该区域的肌球蛋白结合,从而降低这种肌纤维产生的张力。 如果肉瘤进一步缩短,细丝就会开始相互重叠,从而进一步减少交叉桥的形成,产生的张力甚至更小。 相反,如果肉瘤被拉伸到粗细丝根本不重叠的程度,则不会形成交叉桥,也不会产生张力。 这种程度的拉伸通常不会发生,因为辅助蛋白、内部感觉神经和结缔组织会阻碍极度伸展。
决定力产生的主要变量是肌肉内从控制该纤维的神经元获得动作电位的肌纤维的数量。 使用二头肌拿起铅笔时,大脑的运动皮层仅向二头肌的几个神经元发出信号,只有少数肌纤维有反应。 在脊椎动物中,每根肌纤维在受到刺激时都会充分反应。 拿起钢琴时,运动皮层向二头肌中的所有神经元发出信号,每根肌纤维都参与其中。 这接近肌肉可以产生的最大力。 如上所述,增加动作电位的频率(每秒的信号数)可以进一步增加力,因为原肌球蛋白中充斥着钙。
摘要
人体包含三种类型的肌肉组织:骨骼肌、心肌和平滑肌。 骨骼肌肉组织由肉瘤组成,肉瘤是肌肉组织的功能单位。 当肉瘤缩短时,就会发生肌肉收缩,因为粗细丝相互滑过,这被称为肌肉收缩的滑丝模型。 ATP 为跨桥形成和灯丝滑动提供能量。 调节蛋白,例如肌钙蛋白和原肌球蛋白,控制交叉桥的形成。 激发收缩耦合通过乙酰胆碱将神经元的电信号转化为肌肉膜上的电信号,从而启动力产生。 肌肉纤维收缩的数量决定了整块肌肉产生的力。
艺术联系
图\(\PageIndex{5}\):以下关于肌肉收缩的陈述中哪一项是正确的?
- 当 ATP 水解成 ADP 和磷酸盐时,就会发生强力冲击。
- 当 ADP 和磷酸盐与肌球蛋白头分离时,就会发生强力冲击。
- 当 ADP 和磷酸盐与肌动蛋白活性位点分离时,就会发生强力中风。
- 当 Ca 2+ 结合钙头时,就会发生强力冲击。
- 回答
-
B
图\(\PageIndex{6}\):致命的神经毒气沙林不可逆转地抑制乙酰胆碱酯酶。 沙林会对肌肉收缩产生什么影响?
- 回答
-
在沙林存在的情况下,乙酰胆碱不会从突触中去除,从而持续刺激肌肉质膜。 起初,肌肉活动剧烈且不受控制,但离子梯度会消散,因此 T 型管中的电信号不再可能。 结果是瘫痪,导致窒息死亡。
词汇表
- 肌动蛋白
- 与肌球蛋白相互作用以收缩肌肉的球状收缩蛋白
- 乙酰胆碱酯酶
- (acHe) 将乙酰胆碱分解成乙酰和胆碱的酶
- 心肌
- 组织肌肉组织仅存在于心脏中;心脏收缩将血液泵送到全身并维持血压
- 电机端板
- 与神经元相互作用的肌肉纤维的肌肉瘤
- myofibril
- 平行于肌肉纤维的长圆柱结构
- myofilament
- 构成肌原纤维的小结构
- 肌球蛋白
- 与肌动蛋白相互作用以收缩肌肉收缩的收缩蛋白
- sarcolemma
- 骨骼肌纤维的质膜
- 肉瘤
- 骨骼肌的功能单位
- 骨骼肌组织
- 形成骨骼肌,骨骼肌附着在骨骼上并控制运动和任何可以有意识控制的运动
- 平滑肌
- 组织存在于肠道、胃和膀胱等空心器官的壁上,以及呼吸道和血管等通道周围
- 厚灯丝
- 一组肌球蛋白分子
- 细灯丝
- 两种肌动蛋白聚合物与原肌球蛋白和肌钙蛋白一起缠绕在一起
- 原肌球蛋白
- 起到阻断肌动蛋白分子上肌球蛋白结合位点的作用,防止交叉桥形成并在肌肉收到神经元信号之前防止收缩
- 肌钙蛋白
- 与原肌球蛋白结合并帮助将其定位在肌动蛋白分子上,还可以结合钙离子