
32.2: 授粉和受精
- Page ID
- 202353

培养技能
- 描述植物施肥必须发生的事情
- 解释异花授粉及其发生方式
- 描述导致种子发育的过程
- 定义双重受精
在被子植物中,授粉的定义是将花粉从花药放置或转移到同一朵花或另一朵花的柱头上。 在裸子植物中,授粉涉及花粉从雄锥转移到雌锥体。 转移后,花粉会发芽形成花粉管和用于使卵子受精的精子。 自格雷戈尔·孟德尔时代以来,授粉得到了很好的研究。 孟德尔成功地在园豌豆中进行了自授和异花授粉,同时研究了如何将特征从一代传给下一代。 今天的农作物是植物育种的结果,植物育种采用人工选择来生产当今的品种。 今天的玉米就是一个很好的例子,它是从其祖先泰奥辛特开始多年繁殖的结果。 古代玛雅人最初开始种植的 teosinte 种子很小,与当今相对较大的玉米穗有很大的不同。 有趣的是,尽管这两种植物看起来完全不同,但它们之间的遗传差异微乎其微。
授粉有两种形式:自授粉和异花授粉。 当花药中的花粉沉积在同一朵花的柱头上或同一株植物上的另一朵花上时,就会发生@@ 自授粉。 异花授粉是将花粉从一朵花的花药转移到另一朵花的柱头上,转移到同一物种的另一个个体上。 自授粉发生在花朵中,雄蕊和 carpel 同时成熟,其位置使花粉可以落在花柱上。 这种授粉方法不需要植物投资即可提供花蜜和花粉作为传粉媒介的食物。
链接到学习
浏览这个互动网站查看自授粉和异花授粉:http://passel.unl.edu/pages/animatio...erbreeding.swf
活物种旨在确保其后代的生存;失败的物种就会灭绝。 因此,需要遗传多样性,以便在不断变化的环境或压力条件下,一些后代能够存活。 自授粉会导致产生遗传多样性较少的植物,因为来自同一植物的遗传物质被用来形成配子,最终形成合子。 相比之下,异花授粉或交叉授粉会带来更大的遗传多样性,因为微配子体和巨型配子体来自不同的植物。
由于异花授粉允许更多的遗传多样性,因此植物已经开发出许多避免自授粉的方法。 在某些物种中,花粉和子房在不同的时间成熟。 这些花使自授粉几乎不可能。 当花粉成熟并脱落时,这朵花的柱头已经成熟,只能由另一朵花的花粉授粉。 有些花已经形成了防止自授粉的物理特征。 报春花就是这样一朵花。 报春花已经进化出两种花型,花药和柱头长度各不相同:针眼花的花药位于花粉管的中点,而长眼花的柱头同样位于中点。 昆虫在寻找花粉管底部的花蜜时很容易进行异花授粉。 这种现象也被称为异性现象。 许多植物,例如黄瓜,在植物的不同部位都有雄花和雌花,因此难以自授粉。 在其他物种中,雄花和雌花生在不同的植物上(雌雄异株)。 所有这些都是自授粉的障碍;因此,植物依靠传粉媒介转移花粉。 大多数传粉媒介是生物因子,例如昆虫(如蜜蜂、苍蝇和蝴蝶)、蝙蝠、鸟类和其他动物。 其他植物物种是由风和水等非生物剂授粉的。
日常连接:花中的不相容基因
近几十年来,在许多被子植物物种中都发现了不相容的基因,这些基因会阻止花粉发芽或长成花的柱头。 如果植物没有相容的基因,花粉管就会停止生长。 自我不相容由 S(无菌)轨迹控制。 花粉管必须先通过柱头和样式的组织生长,然后才能进入胚珠。 carpel 允许在里面生长的花粉类型是有选择性的。 这种相互作用主要发生在花粉和柱状表皮细胞之间。 在某些植物中,例如卷心菜,花粉在柱头表面被排出,不需要的花粉不会发芽。 在其他植物中,花粉管长度为花粉管长度的三分之一后就会停止发芽,从而导致花粉管死亡。 花粉管死亡要么是由于细胞凋亡(程序性细胞死亡),要么是花粉管 RNA 的降解。 降解是由S位点编码的核糖核酸酶的活性造成的。 核糖核酸酶是从细胞外基质中的同类细胞中分泌出来的,细胞外基质位于不断增长的花粉管旁边。
总而言之,自相容性是一种阻止许多开花植物物种自我受精的机制。 这种自不相容机制的运作对植物育种者具有重要影响,因为它抑制了近交和杂交植物的产生。
昆虫授粉
蜜蜂可能是许多园林植物和大多数商业果树中最重要的传粉媒介(图\(\PageIndex{1}\))。 最常见的蜜蜂种类是大黄蜂和蜜蜂。 由于蜜蜂看不到红色,蜜蜂授粉的花通常有蓝色、黄色或其他颜色的阴影。 蜜蜂收集能量丰富的花粉或花蜜以满足其生存和能量需求。 他们拜访的花朵在白天开放,颜色鲜艳,有强烈的香气或气味,呈管状的花朵,通常会有花蜜向导。 花蜜指南包括花瓣上只有蜜蜂才能看见的区域,人类看不见;它有助于将蜜蜂引导到花的中心,从而提高授粉过程的效率。 花粉粘在蜜蜂模糊的头发上,当蜜蜂拜访另一朵花时,一些花粉会转移到第二朵花上。 最近,有许多关于蜜蜂数量减少的报道。 如果蜜蜂消失,许多花将保持未授粉状态,也不会播种。 对商业水果种植者的影响可能是毁灭性的。

许多苍蝇会被有腐烂气味或肉味腐烂的花所吸引。 这些花会产生花蜜,通常颜色暗淡,例如棕色或紫色。 它们存在于尸花或伏都教百合(Amorphallus)、龙阿鲁姆(Dracunculus)和腐肉花(Stapleia、Rafflesia)上。 花蜜提供能量,而花粉提供蛋白质。 黄蜂也是重要的昆虫传粉媒介,可以为许多种类的无花果授粉。
蝴蝶,例如君主,为许多园林花卉和野花授粉,这些花卉和野花通常成群出现。 这些花颜色鲜艳,有浓郁的香味,白天开放,并有花蜜指南,可以更轻松地获得花蜜。 花粉被捡起并带到蝴蝶的四肢上。 另一方面,飞蛾在傍晚和深夜为花朵授粉。 飞蛾授粉的花是浅色或白色的,是扁平的,使飞蛾能够着陆。 飞蛾授粉植物的一个经过充分研究的例子是丝兰植物,它由丝兰蛾授粉。 花和飞蛾的形状已经进行了调整,可以成功授粉。 飞蛾将花粉沉积在粘稠的柱头上,以便稍后进行受精。 雌蛾还会将卵子沉积到子房中。 当卵发育成幼虫时,它们会从花中获取食物并发育种子。 因此,在这种共生关系中,昆虫和花都相互受益。 玉米耳虫蛾和高拉植物有相似的关系(图\(\PageIndex{2}\))。

蝙蝠授粉
在热带和沙漠中,蝙蝠通常是龙舌兰、番石榴和牵牛花等夜间活动花朵的传粉媒介。 花朵通常又大又白或浅色;因此,可以将它们与夜晚的黑暗环境区分开来。 这些花具有浓郁的果香或麝香味,会产生大量的花蜜。 它们自然很大,嘴巴很宽,可以容纳蝙蝠的头部。 当蝙蝠寻找花蜜时,它们的脸和头部被花粉覆盖,然后花粉被转移到下一朵花上。
鸟类授粉
许多种类的小鸟,例如蜂鸟(图\(\PageIndex{3}\))和太阳鸟,都是兰花和其他野花等植物的传粉媒介。 鸟儿拜访的花通常很坚固,其方向可以让鸟儿待在花朵附近,而不会让翅膀被附近的花朵缠住。 花通常呈弯曲的管状形状,可以让鸟的喙进入。 白天开着的色彩鲜艳、无味的花朵由鸟类授粉。 当一只鸟寻找能量丰富的花蜜时,花粉会沉积在鸟的头部和脖子上,然后转移到它访问的下一朵花上。 众所周知,植物学家通过从同一地点收集和鉴定具有200年历史的鸟类标本中的花粉来确定灭绝植物的范围。

风中授粉
大多数种类的针叶树和许多被子植物,例如草、枫树和橡树,都是通过风授粉的。 松果是棕色的,没有香味,而风授粉的被子植物的花通常是绿色的,很小,花瓣可能很小或没有花瓣,并且会产生大量的花粉。 与典型的昆虫授粉花不同,适应风授粉的花不会产生花蜜或气味。 在风授粉的物种中,微孢子囊在花中悬挂,随着风的吹来,轻质花粉随之携带(图\(\PageIndex{4}\))。 花朵通常在春季初出现,在落叶之前,这样叶子就不会阻挡风的移动。 花粉沉积在花朵裸露的羽毛状柱头上(图\(\PageIndex{5}\))。


用水授粉
有些杂草,例如澳大利亚海草和池塘杂草,是由水授粉的。 花粉漂浮在水上,当它与花接触时,它会沉积在花中。
进化连接:通过欺骗进行授粉
兰花是价值很高的花朵,有许多稀有品种(图\(\PageIndex{6}\))。 它们生长在一系列特定的栖息地中,主要是在亚洲、南美洲和中美洲的热带地区。 已经确定了至少25,000种兰花。

花通常以花蜜的形式吸引传粉媒介,并获得食物奖励。 但是,某些种类的兰花是该标准的例外:它们已经进化出不同的方式来吸引所需的传粉媒介。 他们使用一种被称为食物欺骗的方法,即提供鲜艳的色彩和香水,但不提供食物。 Anacamptis morio,俗称绿翅兰花,带有鲜艳的紫色花朵,散发出强烈的气味。 大黄蜂是它的主要传粉媒介,它之所以被花所吸引,是因为它具有强烈的气味(通常表示蜜蜂的食物),在此过程中,它会吸收花粉运送到另一朵花中。
其他兰花使用性欺骗手段。 Chiloglottis trapezifor mis 释放出的化合物闻起来与雌性黄蜂为吸引雄性黄蜂而释放的信息素相同。 雄性黄蜂被这种气味所吸引,落在兰花上,并在此过程中转移花粉。 有些兰花,例如澳大利亚锤兰,在另一种性欺骗策略中使用气味和视觉诡计来吸引黄蜂。 这种兰花的花模仿雌性黄蜂的外观并散发出费洛蒙。 雄性黄蜂试图与看似雌性黄蜂的交配,并在此过程中捡起花粉,然后将其转移到下一个假冒的伴侣身上。
双重受精
花粉沉积在柱头上后,它必须发芽并在样式中生长才能到达胚珠。 微孢子或花粉包含两个细胞:花粉管细胞和生成细胞。 花粉管细胞长成花粉管,生成细胞通过花粉管传播。 花粉管的发芽需要水、氧气和某些化学信号。 当花粉管穿过样式到达胚囊时,花粉管的生长由该样式的组织支持。 同时,如果生成细胞尚未分裂成两个细胞,它现在会分裂成两个精子细胞。 花粉管由胚囊中存在的协同作用分泌的化学物质引导,然后通过微孔进入胚珠囊。 在这两个精子细胞中,一个精子使卵细胞受精,形成二倍体合子;另一个精子与两个极性核融合,形成发育成胚乳的三倍体细胞。 被子植物中的这两个受精事件加在一起被称为双重受精(图\(\PageIndex{7}\))。 受精完成后,其他精子无法进入。 受精的胚珠形成种子,而子房的组织变成果实,通常包裹着种子。
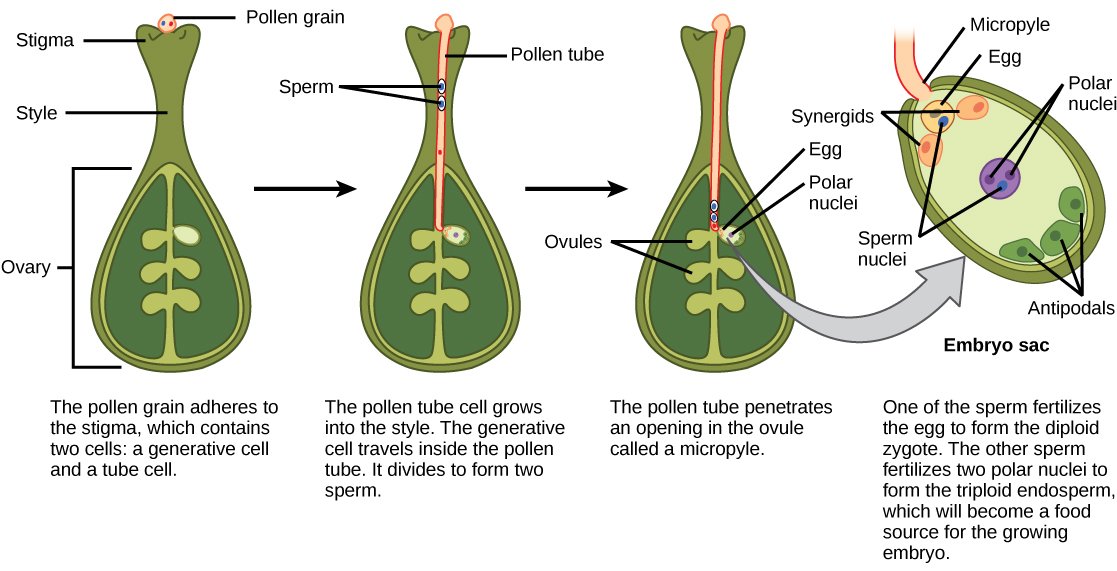
受精后,合子分裂形成两个细胞:上部细胞或末端细胞和下部细胞或基底细胞。 基底细胞的分裂产生悬浮液,悬浮物最终与母体组织建立联系。 悬浮液为营养从母株输送到成长中的胚胎提供了途径。 末端细胞也会分裂,产生球状的前胚胎(图\(\PageIndex{8}\) a)。 在 dicots(eudicots)中,由于存在两个基本子叶,发育中的胚胎呈心形(图\(\PageIndex{8}\) b)。 在非内精子双子体中,例如 Capsella burs a,胚乳最初发育,但随后被消化,食物储备被转移到两个子叶中。 随着胚胎和子叶的扩大,它们在发育中的种子内部空间不足,被迫弯曲(图\(\PageIndex{8}\) c)。 最终,胚胎和子叶填满了种子(图\(\PageIndex{8}\) d),种子已准备好传播。 胚胎发育在一段时间后暂停,只有种子发芽后才能恢复生长。 在第一组叶子开始光合作用之前,发育中的幼苗将依赖子叶中储存的食物储备。
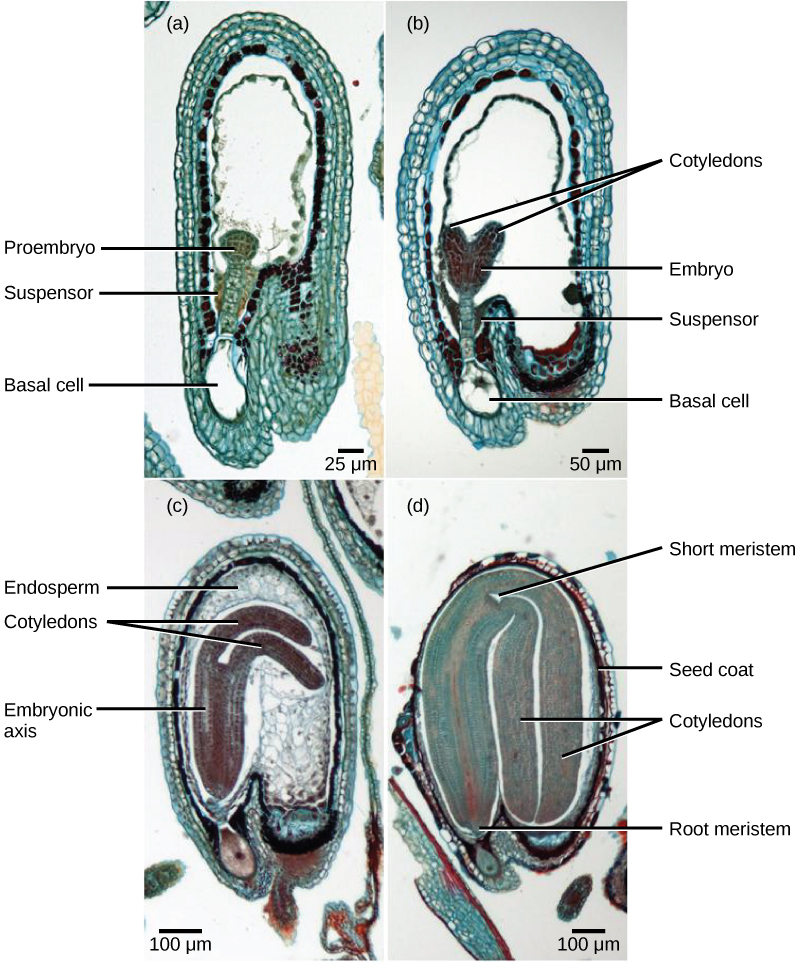
种子的发育
成熟的胚珠发育成种子。 典型的种子包含种皮、子叶、胚乳和单个胚胎(图\(\PageIndex{9}\))。
艺术连接
以下陈述中哪一项是正确的?
- 单子叶和双子叶都有胚乳。
- 根发育成根。
- 羽毛是表心的一部分。
- 胚乳是胚胎的一部分。
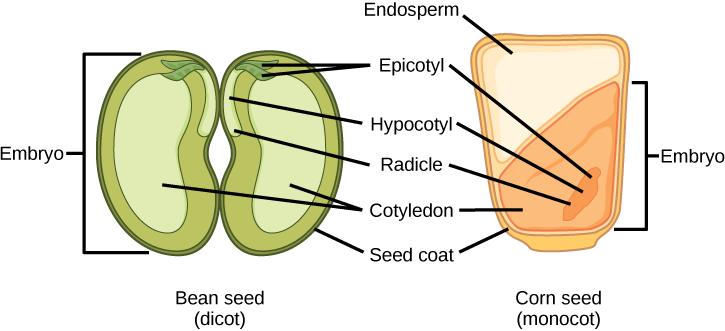
被子植物种子中食物储备的储存因单子植物和双子叶植物而异。 在玉米和小麦等单子叶中,单子叶被称为 scutellum;scutellum 通过血管组织(木质部和韧皮)直接与胚胎相连。 食物储备储存在大型胚乳中。 发芽后,酶由 aleurone 分泌,这是一层位于种皮内的单层细胞,围绕着胚乳和胚胎。 这些酶会降解储存的碳水化合物、蛋白质和脂质,其产物被 scutellum 吸收并通过血管系统链运送到发育中的胚胎。 因此,scutellum 可以看作是吸收器官,而不是储藏器官。
双子种子中的两个子叶也与胚胎有血管连接。 在胚精二子体中,食物储备储存在胚乳中。 因此,在发芽过程中,这两个子叶充当吸收器官,吸收酶释放的食物储备,就像单子叶一样(顾名思义,单子叶也有子精子种子)。 烟草(Nicotiana tabaccum)、番茄(Solanum lycopersicum)和胡椒(C apsicum annum)就是精子内分泌双胞胎的例子。 在非内精子双子体中,三倍体胚乳在双重受精后正常发育,但胚乳的食物储备很快被重新调动并转移到发育中的子叶中进行储存。 花生种子(Arachis hypogaea)的两半和豌豆汤中的豌豆(P isum sativum)是充满食物储备的单独子叶。
种子和胚珠受到由胚珠囊外皮形成的种皮的保护。 在 dicots 中,种皮进一步分为称为 testa 的外层和称为 te gmen 的内层。
胚胎轴由三部分组成:羽毛、根和下胚轴。 子叶附着点和根之间的胚胎部分被称为下胚轴(下胚轴的意思是 “子叶下方”)。 胚胎轴终止于根部(胚胎根),这是根部发育的区域。 在 dicots 中,hypocotyls 延伸到地上,产生植物的茎。 在单子叶植物中,下胚轴不显示在地上,因为单子叶子植物不表现出茎伸长率。 胚胎轴中伸出子叶上方的部分被称为 e picotyl。 羽毛由 epicotyl、幼叶和芽顶分生体组成。
在 dicot 种子中发芽后,epicotyl 的形状像钩子,羽毛指向下方。 这种形状被称为 plumule hook,只要在黑暗中发芽,它就会持续存在。 因此,当 epicotyl 穿过坚硬而具有磨蚀性的土壤时,羽毛会受到保护,免受损坏。 暴露在光线下,下胚轴钩会伸直,幼树叶面对阳光膨胀,epicotyl 继续伸长。 在这段时间里,根也在生长并产生初级根。 当它向下生长形成水龙头根时,侧根会向四面分支,产生典型的双子龙头根系统。
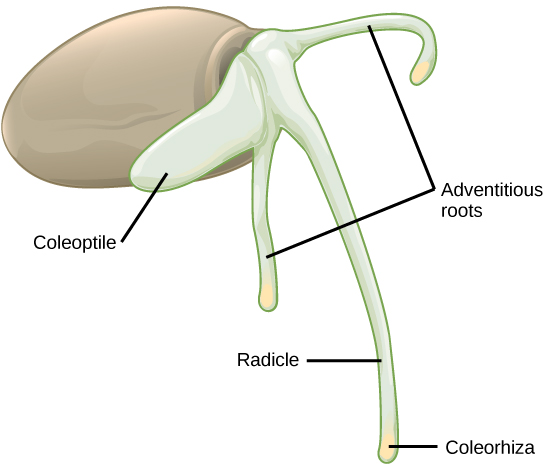
在单子叶种子中(图\(\PageIndex{10}\)),种皮的 testa 和 tegmen 融合在一起。 随着种子的发芽,初级根会出现,受到根尖覆盖物的保护:coleorhiza。 接下来,主要射击出现了,受 coleoptile 的保护:射击尖端的遮盖物。 暴露在光线下(即当羽毛离开土壤并且不再需要保护性的 coleoptile 时),coleoptile 的伸长停止,叶子会膨胀和展开。 在胚胎轴的另一端,初级根很快就会死亡,而其他不定根(不是来自通常位置的根,即根)则从茎的底部出现。 这为单子叶提供了纤维根系。
种子发芽
许多成熟种子进入不活跃期,或新陈代谢活性极低的时期:这种过程被称为休眠,可能持续数月、数年甚至几个世纪。 休眠有助于在不利条件下保持种子的存活力。 恢复到有利条件后,种子就会发芽。 有利的条件可能多种多样,例如湿度、光线、寒冷、火灾或化学处理。 暴雨过后,出现了许多新的幼苗。 森林火灾还导致新幼苗的出现。 有些种子需要春化(冷处理)才能发芽。 这保证了植物在温带气候下产生的种子要到春天才会发芽。 在炎热气候中生长的植物的种子可能需要热处理才能发芽,以避免在炎热干燥的夏天发芽。 在许多种子中,厚厚的种皮的存在会阻碍发芽能力。 划痕,包括软化种皮的机械或化学过程,通常在发芽之前使用。 也可以使用在热水中预浸泡或穿过酸性环境,例如动物的消化道。
根据种子的大小,幼苗出苗所需的时间可能会有所不同。 种子大的物种有足够的食物储备,可以在地下深处发芽,并且仍然可以将其表皮一直延伸到土壤表面。 小种子种的种子通常需要光作为发芽线索。 这样可以确保种子只在土壤表面(光线最充足的地方)或附近发芽。 如果它们在地表之下发芽得太远,发育中的幼苗就没有足够的食物储备来照射阳光。
水果和水果类型的开发
受精后,花的子房通常会发育成果实。 水果通常与甜味有关;但是,并非所有水果都是甜的。 从植物学上讲,“水果” 一词用于成熟的子房。 在大多数情况下,受精的花朵会长成果实,而未受精的花朵则不会。 有些果实是从子房发育出来的,被称为真正的果实,而另一些则是从雌配子体的其他部分发育而来的,被称为辅助果实。 果实包围种子和发育中的胚胎,从而为其提供保护。 水果有多种类型,取决于它们的来源和质地。 黑莓的甜组织、番茄的红肉、花生的外壳和玉米壳(吃爆米花时会卡在牙齿里的坚硬而薄的部分)都是水果。 随着果实的成熟,种子也会成熟。
水果可分为简单水果、聚合水果、多重水果或附属水果,具体取决于其来源(图\(\PageIndex{11}\))。 如果果实由单个子房的单个心脏发育而成,则它被称为简单的水果,如坚果和豆类所示。 聚合果实是指由多个 carpel 发育而成的果实,但它们都在同一朵花中:成熟的 carpels 融合在一起形成整个果实,如覆盆子所示。 多个果实由花序或一簇花发育而成。 菠萝就是一个例子,花朵融合在一起形成果实。 辅助果实(有时称为假果)不是来自子房,而是来自花的另一部分,例如容器(草莓)或 hypanthium(苹果和梨)。

水果通常分为三部分:外果皮(最外层的皮肤或覆盖物)、中果皮(果实的中间部分)和内果皮(果实的内部)。 这三者加在一起被称为 pericarp。 中果皮通常是水果中肉质、可食用的部分;但是,在某些水果中,例如杏仁,内果是可食用的部分。 在许多水果中,两层或全部三层是融合的,成熟时无法区分。 水果可以是干的也可以是肉质的。 此外,水果可以分为开裂型或不结实型。 豌豆等破碎的水果很容易释放种子,而桃子等不熟的水果则依靠腐烂来释放种子。
水果和种子传播
水果只有一个用途:种子传播。 果实中所含的种子需要分散在远离母株的地方,因此它们可能会找到有利且竞争力较弱的发芽和生长条件。
有些水果具有内置机制,因此它们可以自行分散,而另一些则需要风、水和动物等物质的帮助(图\(\PageIndex{12}\))。 种子结构、成分和大小的改变有助于传播。 被风吹散的水果很轻,可能有翅膀状的附属物,可以让它们被风吹走。 有些具有类似降落伞的结构以保持漂浮。 有些水果——例如蒲公英——具有毛茸茸的失重结构,适合通过风传播。
被水分散的种子包含在轻盈有活力的果实中,使它们能够漂浮。 众所周知,椰子能够漂浮在水面上,到达可以发芽的陆地。 同样,柳树和银桦会产生轻质水果,可以漂浮在水面上。
动物和鸟吃水果,未被消化的种子会在一段距离之外的粪便中排出体外。 有些动物,比如松鼠,会埋葬含种子的水果供以后使用;如果松鼠找不到藏匿的果实,如果条件有利,种子就会发芽。 有些水果,比如 cocklebur,有钩子或粘性结构,可以粘在动物的外套上,然后被运送到另一个地方。 当人类将水果运送到新的地方并扔掉含有种子的不可食用部分时,他们在传播种子方面也起着重要作用。
上述所有机制都允许种子在太空中分散,就像动物的后代可以移动到新的位置一样。 如前所述,种子休眠允许植物通过时间分散其后代:这是动物无法做到的。 休眠种子可能需要等待数月、数年甚至数十年,才能为该物种的发芽和繁殖创造适当的条件。



摘要
要在被子植物中进行受精,必须将花粉转移到花的柱头上:这个过程被称为授粉。 裸子植物授粉涉及将花粉从雄锥转移到雌锥体。 当花的花粉转移到同一朵花的柱头上时,它被称为自授粉。 当花粉从一朵花转移到同一株植物上的另一朵花或另一株植物时,就会发生异花授粉。 异花授粉需要授粉剂,例如水、风或动物,并增加遗传多样性。 花粉落在柱头上后,管细胞产生花粉管,生成核通过花粉管迁移。 花粉管通过胚珠囊上的微孔进入。 生成细胞分裂形成两个精子细胞:一个与卵子融合形成二倍体合子,另一个与极性核融合形成胚乳,胚乳本质上是三倍体。 这就是所谓的双重受精。 受精后,合子分裂形成胚胎,受精的胚珠形成种子。 子宫壁形成果实,种子在其中发育。 种子成熟后将在有利的条件下发芽并产生二倍体孢子体。
艺术联系
图\(\PageIndex{9}\):子叶的功能是什么?
- 它发展成根。
- 它为胚胎提供营养。
- 它形成了胚胎。
- 它可以保护胚胎。
- 回答
-
B
词汇表
- 配件水果
- 源自卵巢以外的组织的果实
- 聚合水果
- 由同一朵花中的多个 carpels 发育而成的水果
- aleurone
- 种皮内部的单层细胞,在发芽时分泌酶
- coleoptile
- 芽尖的覆盖物,存在于发芽的单子种子中
- coleorhiza
- 根尖覆盖物,存在于发芽的单子种子中
- cotyledon
- 种子的肉质部分,为种子提供营养
- 异花授粉
- 将花粉从一朵花的花药转移到另一朵花的柱头上
- 休眠
- 无生长期,新陈代谢过程非常缓慢
- 双重受精
- 被子植物中有两次受精事件;一个精子与卵子融合形成合子,而另一个精子与极性核融合,形成胚乳
- endocarp
- 水果最里面的部分
- 胚乳
- 三倍体结构由精子与极性核融合而成,极性核是胚胎的营养组织
- 精子内分泌双胞胎
- 在胚乳中储存食物储备的 dicot
- exocarp
- 水果的最外层覆盖物
- epicotyl
- 子叶上方的胚芽
- gravitropism
- 植物在与重力相同方向生长的反应
- hypocyl
- 子叶上方的胚胎轴
- mesocarp
- 水果的中间部分
- 多种水果
- 由花序上的多朵花发育而成的果实
- 花蜜指南
- 花上的颜料图案引导昆虫找到花蜜
- 非精子内分泌的 dicot
- dicot 在发育中的子叶中储存食物储备
- pericarp
- 描述 exocarp、mesocarp 和 endocarp 的统称;围绕种子的结构,是果实的一部分
- plumule
- 由发芽的种子发育而成的芽
- 授粉
- 将花粉转移到柱头上
- 激进
- 由发芽种子发育出来的原始根
- 划痕
- 软化种皮的机械或化学过程
- scutellum
- 在单子叶中发现的 cotyledon 的类型,如在草种子中发现的
- 自授粉
- 将花粉从花药转移到同一朵花的柱头上
- 简单的水果
- 由单个 carpel 或融合的 carpels 发育而成的水果
- 悬架
- 成长中的胚胎的一部分,与母体组织有联系
- tegmen
- 种皮的内层
- testa
- 种皮的外层
- 春化
- 一些种子在发芽之前需要暴露在寒冷中