
28.3: Superphylum Lophotrochozoa
- Page ID
- 203238

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 描述扁虫、轮虫、Nemertea、软体动物和环节动物的独特解剖和形态学特征
- 描述 extrocoelomic 腔的发育过程
- 讨论真体分割的优势
- 解释 Platyhelminthes 的主要特征及其作为寄生虫的重要性
- 描述在 phylum Annelida 中分类的动物的特征
属于 superphylum Lophotrochozoa 的动物是原生体,其中 blastopore 或外胚层或外部细菌层的融合点变成了通往消化道的嘴巴。 这被称为原生造口术或 “第一口”。 在原生造口术中,固体细胞群从内胚层或内部生殖层分裂出来,形成中心中胚层细胞。 这层成倍成一个波段,然后在内部分裂形成 coelom;因此,这个原生的 coelom 被称为 s chizocoelom。
作为 lophotrochozoans,这个 superphylum 中的生物要么拥有 lophophore 要么是 trochophore 幼虫。 lophophores 包括因存在 lophore 而团结在一起的群体,lophophore 是一组围绕嘴巴的纤毛触手。 Lophophorata 包括扁虫和其他几种 phyla。 在比较 RNA 序列时,这些进化枝得以维持。 Trochophore 幼虫的特点是身体周围有两条纤毛带。
lophotrochozoans 是 triploblastic 的,在二倍细胞刺胞动物中发现的外胚层和内胚层之间夹着一个胚胎中胚层。 这些 phyla 也是双边对称的,这意味着纵向截面会将它们分成对称的左右两侧。 这也意味着头脑化的开始,即生物体头部中神经组织和感觉器官浓度的演变,这是它第一次遇到环境的地方。
Phylum Platyhelminthes
扁虫是 acoelomate 生物,包括许多自由生活和寄生形式。 大多数扁虫被归类为 superphylum Lophotrochozoa,其中还包括软体动物和环节动物。 Platyhelminthes 由两个血统组成:Catenulida 和 Rhabditophora。 Catenulida 或 “链虫” 是一个由 100 多个物种组成的小型进化枝。 这些蠕虫通常通过出芽进行无性繁殖。 但是,后代并不完全依附于父母,在外表上就像一条链子。 这里讨论的所有剩余扁虫都是 Rhabditophora 的一部分。 许多扁虫是寄生虫,包括人类的重要寄生虫。 扁虫有三个胚胎组织层,它们产生的表面覆盖组织(来自外胚层)、内部组织(来自中胚层)和消化系统(来自内胚层)。 表皮组织是单层细胞或一层融合细胞(syncytium),覆盖一层纵向肌肉上方的一层圆形肌肉。 中胚层组织包括间充质细胞,这些细胞含有胶原蛋白并支持在表面分泌粘液和其他物质的分泌细胞。 扁虫是 acoelomates,因此它们的身体在消化系统的外表面和腔体之间是固体的。
扁虫的生理过程
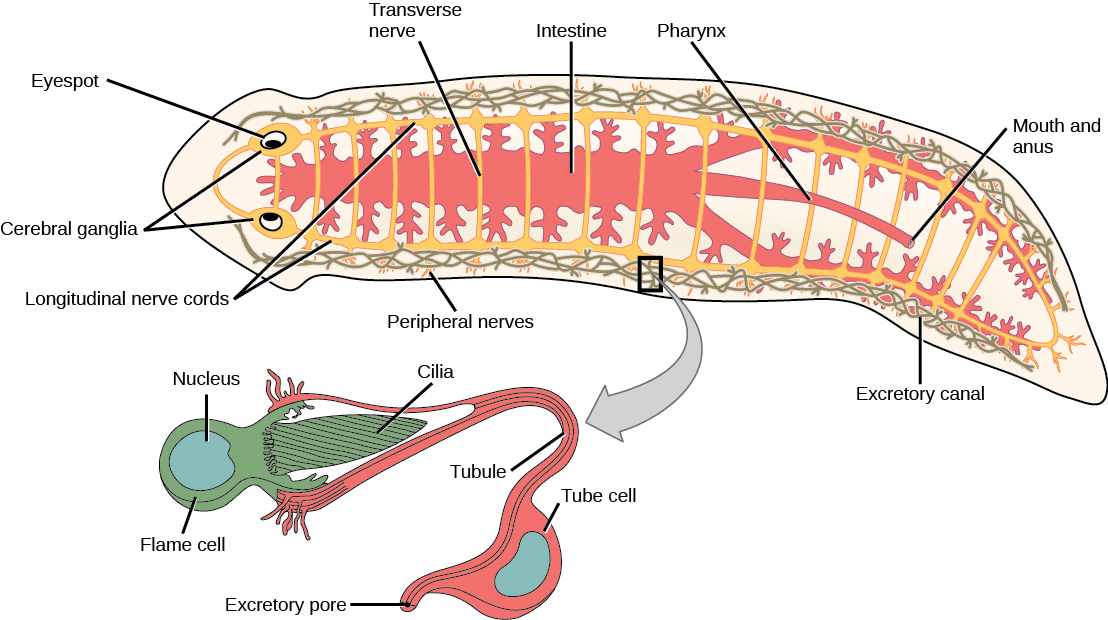
扁虫的自由生活物种是捕食者或食腐动物。 寄生虫以宿主的组织为食。 大多数扁虫,例如图中所示的 planarian\(\PageIndex{1}\),都有胃血管腔而不是完整的消化系统。 在这种动物中,“嘴巴” 还用于从消化系统中排出废物。 有些物种也有肛门开口。 肠道可能是一个简单的囊或高度分支。 消化是细胞外的,消化的物质通过吞噬作用被吸收到肠壁细胞中。 其中一个群体,即 cestodes,缺乏消化系统。 扁虫有一个排泄系统,全身都有小管网络,通向环境和附近的火焰细胞,火焰细胞的纤毛会跳动,将浓缩在小管中的废液引出体外。 该系统负责调节溶解盐和含氮废物的排泄。 神经系统由一对贯穿全身长度的神经绳组成,它们与蠕虫前端的大神经节或神经集中,那里可能还会有光感和化学感官细胞的集中。
既没有循环系统也没有呼吸系统,气体和养分交换依赖于扩散和细胞-细胞连接。 这必然限制了这些生物体内的身体厚度,将它们限制为 “扁平” 蠕虫。
大多数扁虫是雌雄异株的,受精通常是内部的。 无性繁殖在某些群体中很常见。
扁虫的多样性
传统上,Platyhelminthes 分为四类:Turbellaria、Monogenea、Trematoda 和 Cestoda(图\(\PageIndex{2}\))。 如上所述,正在重新评估这些阶层成员之间的关系,尤其是现在将turbellarians视为副系群体,这个群体没有一个共同的祖先。
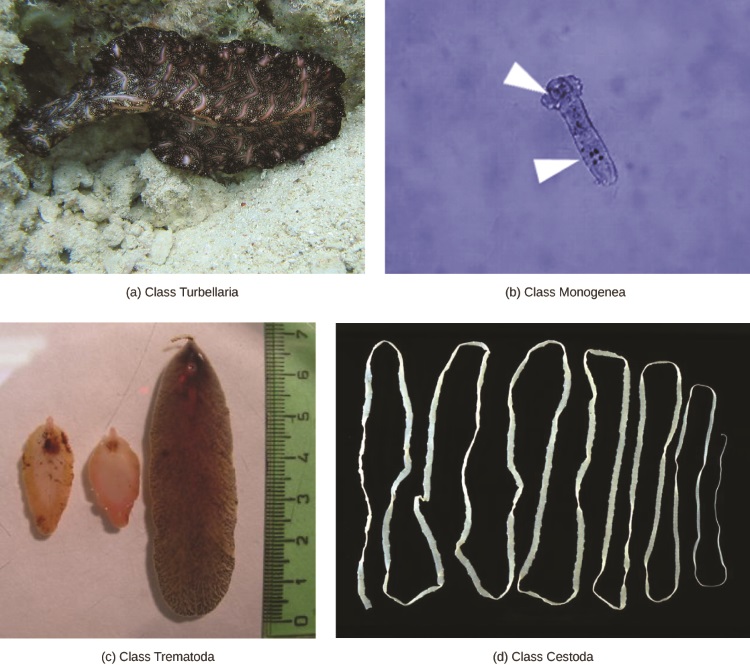
Turbellaria 类别主要包括自由生活的海洋物种,尽管有些物种生活在淡水或潮湿的陆地环境中。 turbellarians 的腹侧表皮是纤毛的,有助于他们的运动。 一些 turbellarians 能够取得非凡的再生壮举,即使是从一小块碎片中也可以使身体再生。
单基因是体外寄生虫,主要是鱼类,其生命周期很简单,包括一只自由游泳的幼虫,它附着在鱼身上,开始转化为寄生成虫形态。 寄生虫只有一个宿主,而该宿主通常只有一个物种。 蠕虫可能产生消化宿主组织的酶,或者只是在表面粘液和皮肤颗粒上吃草。 大多数单基因是雌雄同体,但雄配子首先发育,因此交叉受精很常见。
吸虫或吸虫是软体动物和包括人类在内的许多其他群体的内部寄生虫。 吸虫具有复杂的生命周期,涉及发生有性生殖的主要宿主,以及发生无性繁殖的一个或多个次要宿主。 主要宿主几乎总是软体动物。 吸虫是人类严重疾病的原因,包括血吸虫病,一种血吸虫。 据估计,这种疾病在热带地区感染了2亿人,导致器官损伤和疲劳等慢性症状。 当人类进入水中,从原生蜗牛宿主释放的幼虫找到并穿透皮肤时,就会发生感染。 这种寄生虫会感染人体的各个器官,并在繁殖之前以红细胞为食。 许多卵在粪便中释放出来,然后进入水道,在那里它们能够重新感染主要的蜗牛宿主。
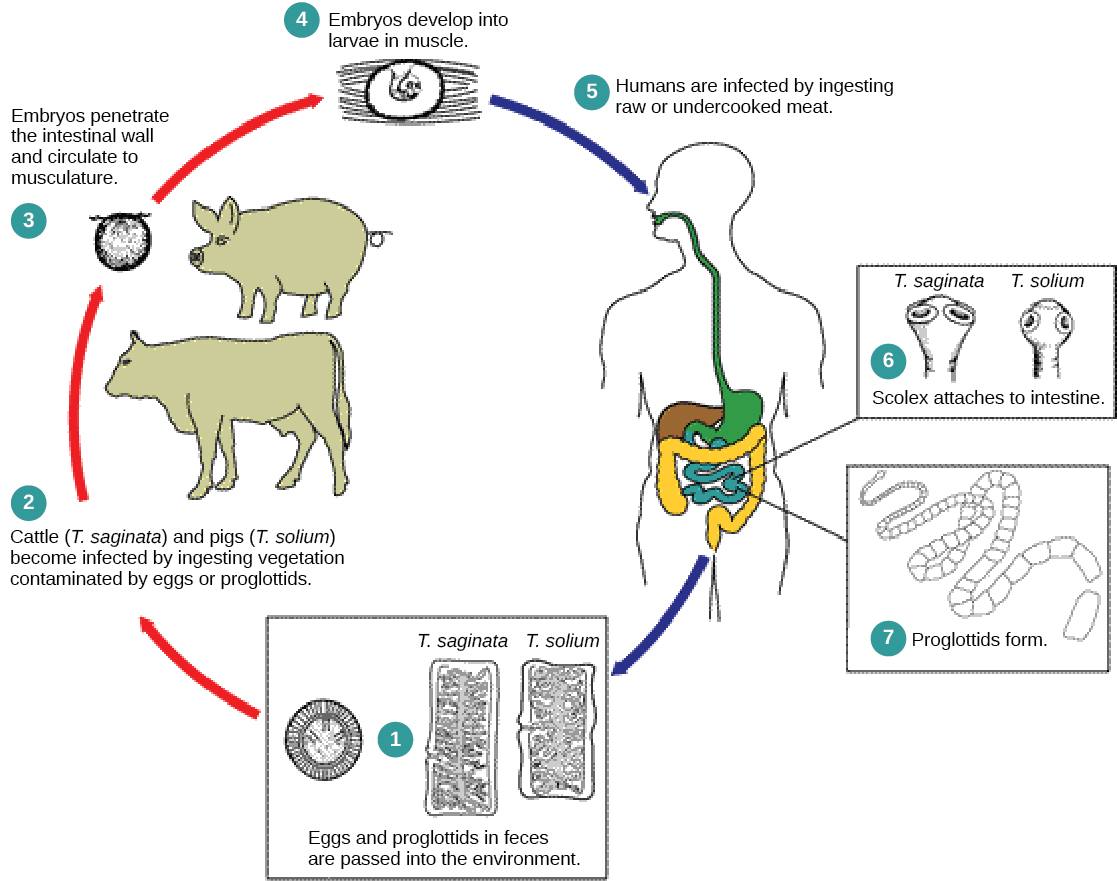
cestodes 或绦虫也是内部寄生虫,主要是脊椎动物(图\(\PageIndex{3}\))。 绦虫生活在主要宿主的肠道中,使用绦虫体前端的吸盘或 scolex 保持固定。 绦虫的剩余身体由一系列被称为 proglottids 的单位组成,每个单位都可能含有含有火焰细胞的排泄系统,但含有雄性和雌性的生殖结构。 绦虫没有消化系统;相反,它们从宿主肠道中传递的食物中吸收营养。
Proglottids 是在 scolex 产生的,然后逐渐迁移到绦虫的末端;此时,它们已经 “成熟” 了,除受精卵以外的所有结构都退化了。 大多数繁殖是通过交叉受精进行的。 proglottid 会脱离蠕虫体内并释放到生物体的粪便中。 鸡蛋被中间宿主吃掉。 幼虫感染中间宿主并居住,通常在肌肉组织中。 当肌肉组织被主要宿主吃掉时,循环就完成了。 人类有几种绦虫寄生虫是通过吃未煮过或煮得不好的猪肉、牛肉和鱼传播的。
Phylum Rotifera
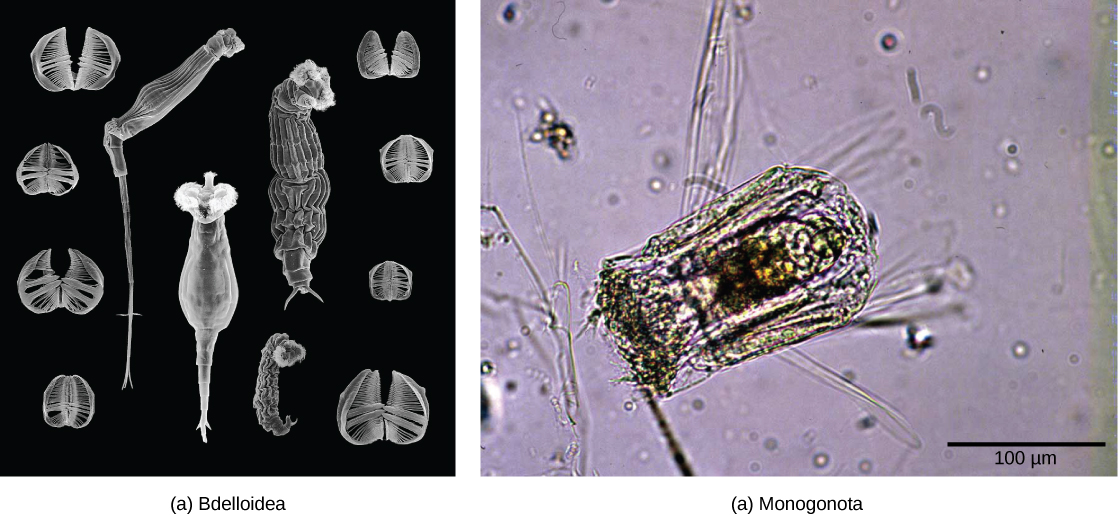
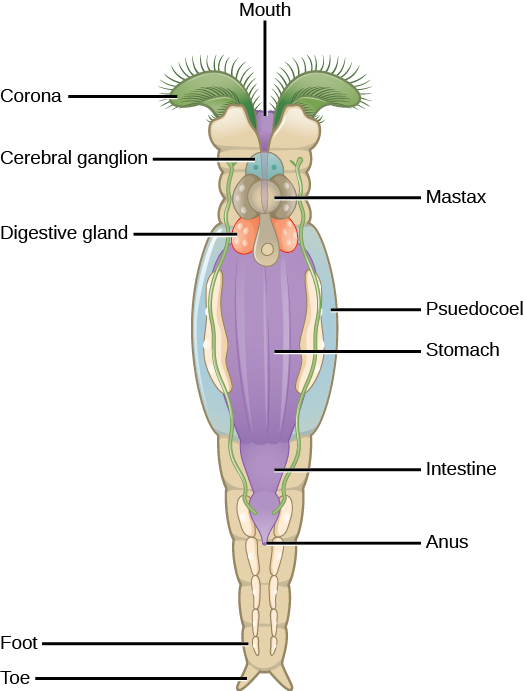
轮虫是一个微观的(大约 100 微米到 30 毫米)群体,主要由水生生物组成,它们的名字来自日冕,这是一种旋转的轮状结构,前端覆盖着纤毛(图\(\PageIndex{4}\))。 尽管轮虫的分类法目前正在不断变化,但有一种疗法将轮虫分为三类:Bdelloidea、Monogonta 和 Seisonidea。 但是,随着越来越多的系统发育证据的出现,该群体的分类目前正在修订中。 目前存在于 phylum Aanthocephala 中的 “刺头蠕虫” 将来可能会被纳入这个群体。
轮虫的身体形态由头部(包含日冕)、躯干(包含器官)和脚组成。 轮虫通常是自由游泳,是真正的浮游生物,但脚趾或脚的延伸部分会分泌一种粘性物质,形成支撑物,帮助它们粘附在表面。 头部包含双叶大脑形式的感觉器官和日冕附近的小眼点。
轮虫是过滤器喂食器,会吃掉死物、藻类和其他微观活生物,因此是水生食物网中非常重要的组成部分。 轮虫获得的食物是由日冕运动产生的电流引导到口腔的。 食物颗粒进入口腔并传播到乳房(咽部有下巴状结构)。 然后,食物通过消化腺和唾液腺,进入胃,然后进入肠道。 消化道和排泄废物在排出肛门之前被收集在阴沟膀胱中。
轮虫是在世界各地的淡水和一些咸水环境中常见的伪生态动物。 该图\(\PageIndex{5}\)显示了属于 Bdelloidea 类的轮虫的解剖结构。 已经确定了大约 2,200 种轮虫。 轮虫是雌雄异株生物(具有雄性生殖器或雌性生殖器),表现出性二态性(雄性和雌性有不同的形式)。 许多物种是单性生殖的,表现出单倍体,这是一种确定性别的方法,在这种方法中,受精卵发育成雌性,未受精卵发育成雄性。 在许多雌雄异株物种中,雄性寿命短且体积较小,没有消化系统,只有一个睾丸。 雌性可以产生能够在恶劣的环境条件下处于休眠状态的卵子以提供保护。
Phylum Nemertea
Nemertea 俗称丝带虫。 phylum Nemertea 的大多数物种是海洋物种,主要是底栖生物或海底居民,估计已知有 900 种物种。 但是,在淡水和陆地栖息地中也记录了 nemertini。 大多数 nemerteans 都是食肉动物,以蠕虫、蛤蜊和甲壳类动物为食。 有些物种是食腐动物,而一些 nemertini 物种,例如 Malacobdella gross a,也与某些软体动物形成了共生关系。 一些物种破坏了蛤蜊和螃蟹的商业捕鱼。 Nemerteans 几乎没有捕食者,有两种物种被当作鱼饵出售。
形态学
丝带蠕虫的大小从1厘米到几米不等。 它们显示出双向对称性和显著的收缩特性。 由于它们的收缩性,它们可以根据环境线索改变形态表现。 phylum Nemertea 中的动物呈现扁平的形态,也就是说,它们从前到后都是扁平的,就像扁平的管子一样。 Nemertea 是柔软且未分段的动物(图\(\PageIndex{6}\))。

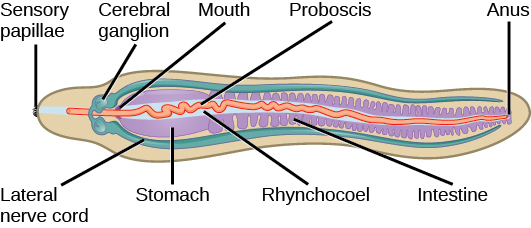
这个 phylum 的一个独特特征是存在封闭在 rhynchocoel 中的长鼻。 长鼻用于捕捉食物,在某些物种中可能会用倒钩装饰。 rhynchocoel 是一个充满液体的空腔,从头部延伸到这些动物肠道长度的近三分之二(图\(\PageIndex{7}\))。 长鼻可能会被附着在 rhynchocoel 壁上的牵开器肌肉伸展或缩回。
消化系统
nemertini 的消化系统非常发达。 rhynchocoel 腹侧的嘴巴通向前肠,然后进入肠道。 肠道以憩室袋的形式存在,末端是通过肛门打开的直肠。 性腺穿插在肠道憩室袋中,通过生殖器毛孔向外开放。 循环系统由一对侧向血管的闭环组成。 循环系统来自胚胎的 coelomic 腔。 有些动物除了侧向血管外,可能还有交叉连接的血管。 尽管它们被称为血管,但由于它们起源于 coelomic,因此循环液是无色的。 有些物种含有血红蛋白以及其他黄色或绿色色素。 血管与 rhynchocoel 相连。 体壁肌肉的收缩促进了这些血管中的液体流动。 这些动物体内存在一对质子肾或原始肾脏,以促进渗透调节。 在 nemertini 中通过皮肤进行气态交换。
神经系统
Nemertini 的前端有一个神经节或 “大脑”,位于口腔和前肠之间,围绕着消化系统和 rhynchocoel。 被称为 “神经节” 的四个神经团环构成了这些动物的大脑。 成对的纵向神经绳从脑神经节中冒出并延伸到后端。 Ocelli 或 eyespots 成对出现,身体前部以二的倍数出现。 据推测,眼斑来自神经组织,而不是表皮。
繁殖
phylum Nemertea 中的动物表现出性二态性,尽管淡水物种可能是雌雄同体。 卵子和精子被释放到水中,受精发生在外部。 然后合子发育成扁平状幼虫。 在某些 nemertine 物种中,pilidium 幼虫可能会在幼虫体内从一系列虚构的椎间盘中发育。 这种幼虫形态的特点是形状像猎鹿帽子,它会吞噬幼虫的组织以求生存,然后变成类似成虫的形态。
Phylum 软体动物
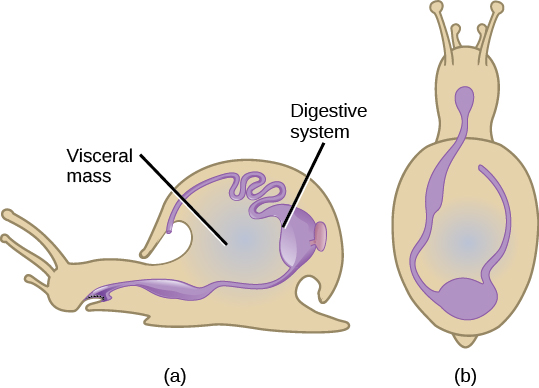
Phylum Mollusca 是海洋环境中主要的 phylum。 据估计,在所有已知的海洋物种中,有23%是软体动物;描述的物种超过75,000种,使它们成为动物中第二多样化的物种。 “软体动物” 这个名字表示软体,因为对软体动物的最早描述来自对无壳墨鱼的观察。 软体动物主要是海洋动物群体;但是,众所周知,它们栖息在淡水和陆地栖息地中。 软体动物在每个类别和亚类中表现出各种各样的形态,但有一些共同的关键特征,包括肌肉足、含有内脏器官的内脏肿块以及可能分泌也可能不会分泌碳酸钙壳的地幔(图\(\PageIndex{8}\))。

练习\(\PageIndex{1}\)
以下关于软体动物解剖结构的陈述中哪一项是错误的?
- 软体动物有用于研磨食物的 radula。
- 消化腺与胃相连。
- 外壳下的组织被称为地幔。
- 消化系统包括 gizzard、胃、消化腺和肠道。
- 回答
-
d
软体动物的足部肌肉发达,用于运动和锚固,其形状和功能各不相同,具体取决于正在研究的软体动物的类型。 在带壳软体动物中,这只脚的大小通常与贝壳开口的大小相同。 脚既是可伸缩的器官,也是可伸缩的风琴。 脚是最腹侧的器官,而地幔是限制性背部器官。 软体动物是 eucoelomate,但是 coelomic 腔仅限于成年动物心脏周围的空腔。 地幔腔独立于 coelomic 腔发育。
内脏肿块存在于脚上方的内脏驼峰中。 这包括消化系统、神经系统、排泄系统、生殖系统和呼吸系统。 完全是水生的软体动物有鳃用于呼吸,而一些陆地物种则有肺用于呼吸。 此外,许多物种中都存在一种叫做 radula 的舌状器官,它带有几丁质牙齿状的装饰,用于切碎或刮掉食物。 地幔(也称为 pallium)是软体动物的背表皮;带壳软体动物专门分泌几丁质和坚硬的钙质壳。
大多数软体动物是雌雄异株动物,受精发生在外部,尽管在陆生软体动物(例如蜗牛和蛞蝓)或头足类动物中并非如此。 在某些软体动物中,合子在成为幼虫之前会孵化并经历两个幼虫阶段:trochophore 和 veliger;双壳类动物可能会出现第三个幼虫阶段,即 glochidia。
Phylum 软体动物的分类
Phylum Mollusca 是一个非常多样化(85,000 种)的群体,主要由海洋物种组成。 软体动物的形态千差万别,从大型掠食性鱿鱼和章鱼(其中一些表现出很高的智力)到带有精心雕刻和彩色贝壳的放牧形态。 这个 phylum 可以分为七类:Aplacophora、Monoplacophora、Polyplacophora、Bivalvia、Gastropoda、Cephalopoda 和 Scaphopoda。
Aplacophora 类(“不带板块”)包括主要存在于海底海洋栖息地的蠕虫状动物。 这些动物没有钙质壳,但表皮上有文石尖刺。 它们的地幔腔很基本,没有眼睛、触手和肾脏(排泄器官)。 Monoplacophora 等级(“带一块盘子”)的成员拥有一个封闭身体的帽状外壳。 贝壳和底层动物的形态可能从圆形到卵形不等。 这些动物体内存在循环的消化系统、多对排泄器官、许多鳃和一对性腺。 据信 monoplacophorans 已经灭绝,在 1952 年发现 Neoplina galathae a 之前,只有通过化石记录才知道。 今天,科学家们已经确定了近二十种现存物种。
Polyplacophora(“携带许多盘子”)类的动物通常被称为 “甲壳素”,带有类似盔甲的八层镀外壳(图\(\PageIndex{9}\))。 这些动物有宽阔的腹足,适合吸入岩石和其他基质,地幔以腰带的形式延伸到贝壳之外。 腰带上可能存在钙质刺,以保护他们免受掠食者的侵害。 腹侧存在的 ctenidia()促进呼吸。 这些动物拥有一只经过改装后可以刮擦的 radula。 神经系统很基本,前端只有颊部或 “脸颊” 神经节。 这些动物身上没有眼斑。 存在一对用于排泄的肾病。

Bivalvia 类(“双壳”)包括蛤蜊、牡蛎、贻贝、扇贝和象拔鸭。 该类的成员存在于海洋和淡水栖息地中。 顾名思义,双壳类动物被封闭在一对贝壳(瓣膜通常称为 “贝壳”)中,它们的背端由贝壳韧带和贝壳齿铰接(图\(\PageIndex{10}\))。 整体形态横向变平,头部区域发育不佳。 某些物种可能没有眼斑和静态囊肿。 由于这些动物是悬浮饲养物,因此此类软体动物中没有 radula。 一对 ctenidia 促进呼吸,而排泄和渗透调节是由一对肾脏引起的。 双壳类动物通常有一个很大的地幔腔。 在某些物种中,地幔的后缘可能会融合形成两个虹吸管,用于吸收和排出水分。

地幔的功能之一是分泌外壳。 一些双壳类动物,例如牡蛎和贻贝,具有在可能进入地幔腔的异物颗粒周围分泌和沉积钙质珍珠层或 “珍珠母” 的独特能力。 该财产已被商业开发用于生产珍珠。
Gastropoda(“胃足”)类动物包括著名的软体动物,例如蜗牛、蛞蝓、海螺、海兔和海蝶。 Gastropoda 包括携带贝壳的物种以及贝壳减少的物种。 这些动物是不对称的,通常呈现盘绕的外壳(图\(\PageIndex{11}\))。 贝壳可能是 planospira l(比如花园水管缠绕起来),常见于花园蜗牛中,也可以是圆锥形的(如螺旋楼梯),常见于海螺中。

带壳物种中的内脏肿块显示出围绕足部中心的垂直轴线的扭转,这是该群体的关键特征,还有一只经过改装后可以爬行的脚(图\(\PageIndex{12}\))。 大多数腹足动物的头部有触手、眼睛和风格。 消化系统使用复杂的 radula,有助于食物的摄入。 某些腹足动物物种可能没有眼睛。 地幔腔围住了 ctenidia 和一对 nephridia。
日常连接:蜗牛毒液可以用作药理止痛药吗?
Conus 属的海蜗牛(图\(\PageIndex{13}\))用毒刺攻击猎物。 释放的毒素被称为 conotoxin,是一种具有内部二硫化物连接的肽。 Conotoxins 会导致人类瘫痪,这表明这种毒素会攻击神经系统靶点。 一些玉米毒素已被证明可以阻断神经元离子通道。 这些发现促使研究人员研究了可能的医疗应用的玉米毒素。
Conotoxins 是一个令人兴奋的潜在药理开发领域,因为这些肽可能会被修饰并在特定的医学条件下使用,以抑制特定神经元的活性。 例如,这些毒素可用于在特定的健康应用中诱发肌肉麻痹,类似于肉毒杆菌毒素的使用。 由于玉米毒素的全部范围及其作用机制尚不完全清楚,因此对其潜在应用的研究仍处于起步阶段。 迄今为止,大多数研究都集中在它们用于治疗神经系统疾病上。 它们在缓解慢性疼痛以及与坐骨神经痛和带状疱疹等疾病相关的疼痛方面也显示出一定的功效。 生物毒素(源自活生物体的毒素)的研究和使用是生物科学应用于现代医学的一个很好的例子。

头足类(“头足” 动物)包括章鱼、鱿鱼、墨鱼和鹦鹉螺。 Cephalopods 是一类携带贝壳的动物,也是贝壳缩小的软体动物。 它们呈现鲜艳的色彩,通常出现在鱿鱼和章鱼身上,用于伪装。 该类中的所有动物都是肉食性捕食者,前端有喙状的下巴。 所有头足类动物都表现出非常发达的神经系统和眼睛以及封闭的循环系统。 脚被裂开并发育成触手,还有一个漏斗,用作它们的运动方式。 章鱼和鱿鱼的触手上有吸盘。 Ctenidia 被封闭在一个大地幔腔中,由大型血管提供服务,每个血管都有自己的心脏与之相关;地幔有促进水交换的虹吸团。
通过喷射水流进行推进,可以促进头足类动物的运动。 这被称为 “喷气式推进”。 地幔腔内存在一对肾炎。 在这类动物中可以看到性二态性。 物种的成员交配,然后雌性在一个隐蔽且受保护的壁垒中产卵。 某些物种的雌性会长时间照顾卵子,最终可能会在这段时间内死亡。 鱿鱼和章鱼等头足类动物也会产生棕褐色或深色墨水,喷射到捕食者身上以帮助他们快速逃跑。
头足类动物的繁殖与其他软体动物的不同之处在于,卵在没有经过 trochophore 和 veliger 幼虫阶段的情况下孵化出幼体成虫。
在装有贝壳的 Nautilus spp. 中,螺旋壳是多腔的。 这些腔室充满气体或水以调节浮力。 鱿鱼和墨鱼的贝壳结构减少了,内部分别以鱿鱼圈和墨鱼骨的形式存在。 示例如图所示\(\PageIndex{14}\)。

图\(\PageIndex{14}\):(a)鹦鹉螺、(b)巨型墨鱼、(c)礁鱿鱼和(d)蓝环章鱼都是 Cephalopoda 类的成员。 (来源 a:J. Baecker 对作品的修改;来源 b:Adrian Mohedano 对作品的修改;来源 c:Silke Baron 对作品的修改;来源 d:安杰尔·威廉姆斯对作品的修改)
Scaphopoda 类(“船脚”)的成员俗称 “象牙壳” 或 “牙壳”,在检查剩下的为数不多的 scaphopod 属之一 D entalium 时可以明显看出(图\(\PageIndex{15}\))。 Scaphopods 通常被埋在沙子里,前开口暴露在水中。 这些动物只有一个圆锥形的外壳,两端都是开着的。 头部很基本,从炮弹的后端伸出来。 这些动物没有眼睛,但它们有 radula,还有一只脚改装成带有球根末端的触手,称为 captaculae。 Captaculae 用于捕捉和操纵猎物。 这些动物中没有 Ctenidia。
图\(\PageIndex{15}\):Antalis vulgaris 展示了经典的 Dentaliidae 形状,它赋予了这些动物的通用名称 “象牙壳”。 (来源:Georges Jansoone)
Phylum Annelida
Phylum Annelida 包括分段蠕虫。 这些动物存在于海洋、陆地和淡水栖息地中,但水或湿度的存在是它们生存的关键因素,尤其是在陆地栖息地中。 phylum 的名字源自拉丁语 annellus,意思是小戒指。 这个 phylum 中的动物表现出与栖息地中其他物种的寄生和共生共生关系。 在 phylum Annelida 中描述了大约 16,500 个物种。 phylum 包括蚯蚓、polychaete 蠕虫和水蛭。 环节动物在胚胎阶段表现出原生发育,通常被称为 “分段蠕虫”,因为它们的关键特征是同种异体或真正的分割。
形态学
环节动物表现出双向对称性,整体形态类似蠕虫。 环节动物具有分段的身体计划,其中内部和外部形态特征在每个身体部位中重复。 Metamerism 允许动物通过添加 “隔间” 来变大,同时提高它们的移动效率。 人们认为,这种异构体源于胚胎阶段的相同的 teloblast 细胞,它们产生相同的中胚层结构。 整体身体可分为头部、身体和 pygidium(或尾巴)。 阴蒂是一种生殖结构,它产生粘液,有助于精子转移,并产生蚕茧,在其中进行受精;它在动物的前三分之一以融合带的形式出现(图\(\PageIndex{16}\))。

图\(\PageIndex{16}\):阴蒂在这里被视为一个突出的部分,其颜色与身体其他部位不同,是一种有助于环节动物繁殖的结构。 (来源:Rob Hille)
解剖学
表皮受到脱细胞外部角质层的保护,但它比在溃疡中发现的角质层薄得多,不需要周期性脱落即可生长。 圆形和纵向肌肉位于表皮内部。 每个部位都存在着锚定在表皮中并从角质层伸出的几丁质头发状接发,称为 setae/ch aetae。 Annelids 显示存在真正的 coelom,它源自胚胎中胚层和原生造口术。 因此,它们是最先进的蠕虫。 蚯蚓(oligochaetes)中存在发育完善且完整的消化系统,有嘴巴、肌肉发达的咽部、食道、农作物和 gizzard。 gizzard 通向肠道并以肛门开口结束。 图中显示了蚯蚓(一种陆地类环节动物)身体部分的横截面图\(\PageIndex{17}\);每个部分都受到膜状隔膜的限制,该隔膜将 coelomic 腔分成一系列隔间。
环节动物拥有封闭的循环系统,由与消化道平行延伸的背侧和腹侧血管以及为单个组织服务的毛细血管组成。 此外,这些船只在每个段中都通过横向环路相连。 这些动物缺乏发达的呼吸系统,气体交换发生在潮湿的体表上。 一对 metanephridia(一种由复杂的小管和开放的纤毛漏斗组成的原始 “肾脏”)促进排泄,它们存在于腹侧的每个部分。 环节动物显示出发育良好的神经系统,咽部周围存在融合神经节的神经环。 神经绳位于腹侧,每个节段都有肿大的节点或神经节。
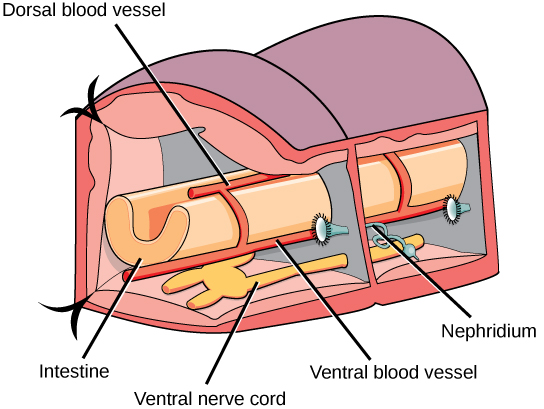
环节动物可以是具有永久性腺的雌雄异株(如蚯蚓、水蛭),也可以是雌雄异株,有临时或季节性性腺发育(如多毛类动物)。 但是,在雌雄同体动物中,交叉受精是首选。 这些动物还可能同时表现出雌雄同体,并在排队进行交配时同时参与精子交换。
Phylum Annelida 的分类
Phylum Annelida 包含 Polychaeta 类(polychaetes)和 Oligochaeta 类(蚯蚓、水蛭及其亲属)。
蚯蚓 Oligochaeta 类中最丰富的成员,其特点是存在阴蒂以及少量减少的 chaetae(“oligo-= “few”;-chaetae = “hairs”)。 与 polychaetes(pol y =many,chaetae = 头发)相比,Oligochaeta 中 chaetae 的数量和大小大大减少了。 许多多毛类动物也排列在肉质、扁平、成对的附属物中,这些附属物从每个名为 parapodia 的片段伸出来,可能专门用于多毛类动物中的不同功能。 Hirudinea 子类包括水蛭,例如 Hirudo medicinalis 和 Hemiclepsis marginata。 Oligochaeta 类包括 Hirudinia 子类和 Brachiobdella 子类。 水蛭和其他环节动物之间的显著区别在于前端和后端会出现吸盘,并且缺乏 chaetae。 此外,体壁的分割可能与 coelomic 腔的内部分割不符。 当水蛭从宿主脊椎动物身上摄取大量血液时,这种适应可能会帮助它们伸长。 Brachiobdella 子类包括 B ranchiobdella balcanica sketi 和 B ranchiobdella astaci 等物种,这些蠕虫与水蛭和寡毛类动物表现出相似之处。

图\(\PageIndex{18}\):(a)蚯蚓、(b)水蛭和(c)featherduster 都是环节动物。 (来源 a:修改 S. Shepherd 的作品;来源 b:“Sarah G...” 对作品的修改 /Flickr;credit c:NOAA Chris Gotschalk 对作品的修改)
摘要
Phylum Annelida 包括 vermiform、分段动物。 在内部解剖学中也可以看到分割,这被称为同质论。 环节动物是原生体。 这些动物具有发育良好的神经元和消化系统。 有些物种有专门的片段,称为阴蒂。 Annelids 显示存在许多被称为 chaetae 的几丁质投影,而 polychaetes 拥有 parapodia。 傻瓜是按顺序看见的 Hirudinea。 生殖策略包括性二态性、雌雄同体和连续雌雄同体。 Hirudinea 课堂中没有内部分割。
扁虫是 acoelomate 的三倍体动物。 它们缺乏循环和呼吸系统,排泄系统很基本。 这种消化系统在大多数物种中是不完整的。 有四类传统的扁虫,主要是自由生活的 turbellarians、ectoparasitic monogenans 以及 endoparasitic trematodes 和 cestodes。 吸虫具有复杂的生命周期,涉及软体动物次要宿主和进行有性生殖的主要宿主。 Cestodes 或绦虫会感染初级脊椎动物宿主的消化系统。
轮虫是微观的、多细胞的,主要是水生生物,目前正在进行分类学修订。 该群体的特点是头上有旋转、纤毛、轮状的结构,即日冕。 mastax 或 jawed pharynx 是这组生物独有的另一种结构。
nemertini 是最简单的 eucoelomates。 这些丝带状的动物带着封闭在 rhynchocoel 中的特殊长鼻。 与其他 pseudocoelomate phyla 相比,源自 coelom 的封闭循环系统的开发是该物种的显著差异。 nemertini 的消化、神经和排泄系统比不太晚期的 phyla 更发达。 nemertine 蠕虫的胚胎发育通过 planuliform 幼虫阶段进行。
Phylum Mollusca 是一大群海洋无脊椎动物。 软体动物在 phylum 内表现出各种形态变化。 这种 phylum 的不同之处在于,有些成员表现出钙质壳作为外部保护手段。 一些软体动物已经进化出精简的外壳。 软体动物是原生动物。 软体动物的背表皮经过修饰形成地幔,它包围了地幔腔和内脏器官。 这个腔与 coelomic 腔完全不同,后者在成年动物中环绕心脏。 呼吸是由被称为 ctenidia 的鳃促进的。 大多数软体动物中都存在一种叫做 radula 的几丁齿舌头。 某些物种的早期发育是通过两个幼虫阶段发生的:trochophore 和 veliger。 在这个 phylum 中,性二态性是主要的性策略。 软体动物可以分为七类,每类都有不同的形态特征。
词汇表
- 安妮丽达
- 患有 metamerism 的 vermiform 动物的 phylum
- captacula
- 触手般的投影,存在于象牙壳中,用于捕捉猎物
- clitellum
- 特殊的融合片段带,有助于繁殖
- 阴谋的
- 围绕水平轴盘绕的外壳形状
- 日冕
- 轮虫前部有轮状结构,含有纤毛,可将食物和水流向嘴巴
- ctenidium
- 软体动物的特殊鳃结构
- 地幔
- (也是 pallium)封闭所有内脏器官并分泌贝壳的特殊表皮
- mastax
- 轮虫独有的下巴咽部
- metamerism
- 一系列内部和外部相似的身体结构,例如分段
- 软体动物
- 有软体且没有分割的原生体的 phylum
- 珍珠层
- 双壳类动物产生的钙质分泌物排列在贝壳的内侧并覆盖侵入的颗粒物
- Nemertea
- 被称为丝带蠕虫的背腹扁平原生体的 phylum
- parapodium
- 肉质、扁平、附属物,成对地从每段多毛类动物中伸出
- pilidium
- 在某些 nemertine 物种中发现了幼虫形态
- planosiral
- 围绕垂直轴盘绕的外壳形状
- planuliform
- 在 phylum Nemertea 中发现了幼虫形态
- radula
- 带有几丁质装饰的舌状器官
- rhynchocoel
- 容纳长鼻的嘴上方有空腔
- schizocoelom
- coelom 由从内皮层分裂出来的细胞群形成
- seta/chaeta
- 角质层的几丁质突出
- trochophore
- 软体动物两个幼虫阶段中的第一个
- veliger
- 软体动物两个幼虫阶段中的第二个