
26.1: 种子植物的进化
- Page ID
- 203289

培养技能
- 解释种子植物何时首次出现以及裸子植物何时成为主要植物群
- 描述允许种子植物在没有水的情况下繁殖的两项主要创新
- 讨论花粉颗粒和种子的用途
- 描述既有花又有果实的被子植物的重要性
第一批在陆地上殖民的植物很可能与现代苔藓(苔藓植物)密切相关,据信大约在5亿年前出现。 其次是肝草(也是 bryophytes)和原始维管植物(pterophytes),现代蕨类植物来自于此。 苔藓植物和紫叶植物生命周期的特点是世代交替,例如裸子植物和被子植物;苔藓植物和叶子植物与裸子植物和被子植物的不同之处在于它们对水的繁殖需求。 完成苔藓植物和叶子植物生命周期需要水,因为雄配子体会释放精子,精子必须在鞭毛的推动下游动,才能到达雌配子或卵子并使之受精。 受精后,合子成熟并长成孢子体,而孢子体反过来又会形成孢子管或 “孢子血管”。 在孢子囊中,母细胞经历减数分裂并产生单倍体孢子。 在合适的环境中释放孢子将导致发芽和新一代配子体的产生。
在种子植物中,进化趋势导致孢子体生成占主导地位,同时配子体大小的系统性缩小:从显眼的结构变为封闭在孢子体组织中的微观细胞群。 低维管植物,例如梭状苔藓和蕨类植物,大多是同孔植物(只产生一种孢子),而所有种子植物或精子植物都是异孔的。 它们形成两种类型的孢子:megaspores(雌性)和微孢子(雄性)。 Megaspores 会发展成产生卵子的雌性配子体,而微孢子则成长为产生精子的雄性配子体。 因为配子体在孢子内成熟,所以它们不能自由生活,其他无核维管植物的配子体也是如此。 异孔无核植物被视为种子植物的进化先驱。
种子和花粉——对干旱和不需要水的繁殖的两种关键适应措施——将种子植物与其他(无籽)维管植物区分开来。 这两种改编都是苔藓植物及其祖先开始的土地殖民化所必需的。 化石将最早的不同种子植物置于大约3.5亿年前。 裸子植物的第一个可靠记录可以追溯到大约3.19亿年前的宾夕法尼亚时期(图\(\PageIndex{1}\))。 裸子植物之前出现了 p rogymnosperms,这是第一批裸种子植物,大约在 3.8 亿年前出现。 Progymnosperms 是一组过渡植物,表面上类似于针叶树(锥形携带者),因为它们从血管组织的次生生长中生产木材;但是,它们仍然像蕨类植物一样繁殖,将孢子释放到环境中。 裸子植物在早期(三叠纪)和中期(侏罗纪)中生代占据了主导地位。 在中生代晚期,被子植物在白垩纪中期(大约1亿年前)超过了裸子植物,如今已成为大多数陆地生物群落中最丰富的植物群。
花粉和种子是创新的结构,它使种子植物能够打破对水的依赖来繁殖和发育胚胎,征服陆地。 花粉颗粒是雄配子体,它含有植物的精子(配子)。 小型单倍体(1 n)细胞被包裹在保护涂层中,以防止干燥(干燥)和机械损坏。 花粉颗粒可以远离其原始孢子体,传播植物的基因。 种子为胚胎提供保护、营养,并提供维持数万年甚至数千年的休眠机制,确保在生长条件最佳时可以发芽。 因此,种子使植物能够在时空中分散下一代。 凭借这样的进化优势,种子植物已成为最成功和最熟悉的植物群体,部分原因是它们的大小和引人注目的外观。
裸子植物的进化
化石植物 Elkinsia polymorph a是泥盆纪时期(大约4亿年前)的 “种子蕨类植物”,被认为是迄今为止已知的最早的种子植物。 种子蕨类植物(图\(\PageIndex{2}\))在没有专门结构的情况下沿着树枝生产种子。 它们之所以成为第一批真正的种子植物,是因为它们开发了称为吸盘的结构来封闭和保护胚珠(雌配子体和相关组织),胚珠在受精后会发育成种子。 在石炭纪时期的煤沼中,类似于现代树蕨的种子植物变得越来越多和多样化。

化石记录表明,第一批裸子植物(progymnosperms)很可能起源于古生代,即泥盆纪中期:大约3.9亿年前。 在密西西比和宾夕法尼亚潮湿的时期(以巨型蕨类植物为主)之后,二叠纪时期处于干燥状态。 这为种子植物提供了繁殖优势,它们更适合在干旱时期中生存。 Ginkgoales 是一组裸子植物,只有一种存活物种——银杏叶,是侏罗纪下游最早出现的裸子植物。 裸子植物在中生代(大约2.4亿年前)扩张,取代了景观中的蕨类植物,并在此期间达到了最大的多样性。 侏罗纪时期既是苏铁(棕榈树状裸子植物)的时代,也是恐龙时代。 银杏树和比较熟悉的针叶树也点缀着景观。 尽管被子植物(开花植物)是大多数生物群落中植物生命的主要形式,但裸子植物仍然在一些生态系统中占主导地位,例如针叶林(北方森林)和高山高山森林(图\(\PageIndex{3}\)),因为它们适应寒冷和干燥的生长条件。

种子和花粉对旱地的进化适应
与苔藓植物和蕨类孢子(它们是依赖水分来快速发育配子体的单倍体细胞)不同,种子含有二倍体胚胎,会发芽成孢子体。 维持生长的储存组织和保护性外套为种子提供了卓越的进化优势。 几层硬化的组织可防止干燥,并且由于需要持续的水供应而自由繁殖。 此外,在生长条件变得有利之前,种子一直处于休眠状态,这是由干燥和脱落酸激素引起的。 无论是被风吹过、漂浮在水面上,还是被动物带走,种子都会散落在不断扩大的地理范围内,从而避免了与母株的竞争。
花粉颗粒(图\(\PageIndex{4}\))是雄性配子体,由风、水或传粉媒介携带。 整个结构不受干燥影响,可以在不依赖水的情况下到达女性器官。 雄配子通过花粉管到达雌配子体和卵细胞配子:花粉颗粒内细胞的延伸。 现代裸子植物的精子缺乏鞭毛,但在苏铁和银杏中,精子仍然具有鞭毛,可以让它们沿着花粉管游到雌配子中;但是,它们被花粉颗粒所包围。
被子植物的进化
无可争议的化石记录显示,被子植物的大规模出现和多样化出现在中生代中后期。 被子植物(“血管中的种子”)会产生一朵含有雄性和/或雌性生殖结构的花。 化石证据(图\(\PageIndex{5}\))表明,开花植物最早出现在下白垩纪,大约在1.25亿年前,并在大约1亿年前在中白垩纪迅速多样化。 早期的被子植物的痕迹很少。 从侏罗纪地质材料中回收的化石花粉被归因于被子植物。 一些早期的白垩纪岩石上有明显的树叶痕迹,类似于被子植物的叶子。 到了白垩纪中期,化石记录中挤满了数量惊人的各种开花植物。 同一地质时期还以许多现代昆虫群体的出现为标志,包括在生态和开花植物进化中起关键作用的授粉昆虫。
尽管有人提出了几种假设来解释这种突然大量多样的开花植物,但没有一个假设得到古植物学家(研究古代植物的科学家)的共识。 但是,比较基因组学和古植物学的新数据为被子植物的进化提供了一些启示。 被子植物不是来自裸子植物,而是形成了与裸子植物并行发育的姊妹进化枝(一种物种及其后代)。 花和果实的两种创新结构代表了一种改进的生殖策略,有助于保护胚胎,同时增加遗传变异性和范围。 古植物学家争论被子植物是从小型木质灌木丛进化而来的,还是与热带草有关的基础被子植物。 这两种观点都得到了分类学研究的支持,而所谓的木质木兰假设——该假设提出被子植物的早期祖先是灌木——也提供了分子生物学证据。
最原始的活被子植物被认为是 Amborella trichopod a,这是一种原产于南太平洋岛屿新喀里多尼亚雨林的小植物。 对 A. trichopod a基因组的分析表明,它与所有现有的开花植物有关,属于被子植物家谱中最古老的已确认分支。 其他一些被子植物群称为基础被子植物,被认为是原始的,因为它们很早就从系统发育树中分支出来。 大多数现代被子植物根据其叶子和胚胎的结构被归类为单子叶或尤迪科特。 基础被子植物,例如睡莲,被认为更原始,因为它们与单子叶子植物和尤迪科斯具有共同的形态特征。

花和果实作为进化适应
被子植物在不同的器官中产生配子,这些器官通常存放在花中。 受精和胚胎发育都发生在解剖结构内,解剖结构提供了稳定的有性生殖系统,在很大程度上不受环境波动的影响。 开花植物是地球上仅次于昆虫的最多样化的 phylum;花的大小、形状、颜色、气味和排列各不相同,令人困惑。 大多数花都有互惠传粉媒介,花的独特特征反映了授粉剂的性质。 传粉媒介和花朵特征之间的关系是共同进化的典型例子之一。
卵子受精后,胚珠长成种子。 卵巢的周围组织变厚,发育成可以保护种子的果实,并通常确保其在广泛的地理范围内传播。 并非所有的果实都是从子房发育而来的;这样的结构是 “假果”。 像花一样,水果在外观、大小、气味和味道上可能有很大差异。 番茄、核桃壳和牛油果都是水果的例子。 与花粉和种子一样,水果也可以起到传播剂的作用。 有些可能会被风吹走。 许多人会吸引动物,这些动物会吃掉水果并将种子传递给消化系统,然后将种子存放在另一个地方。 Cockleburs 上覆盖着坚硬的钩状刺,它们可以钩住毛皮(或衣服),然后长距离搭乘动物。 紧紧抓住富有进取心的瑞士徒步旅行者乔治·德·梅斯特拉尔的天鹅绒长裤的 cockleburs 启发了他发明了他命名为 Velcro 的环扣和钩扣。
进化连接:通过分析 DNA 序列比对建立系统发育树
所有活生物体都表现出源于其进化史的关系模式。 系统发育学是一门描述生物之间在祖先和后代物种方面的相对联系的科学。 系统发育树,例如图中所示的植物进化史\(\PageIndex{6}\),是描绘这些关系的树状分支图。 在树枝的尖端发现了物种。 每个分支点称为节点,是单个分类组(分类单元)(例如物种)分离成两个或更多物种的点。
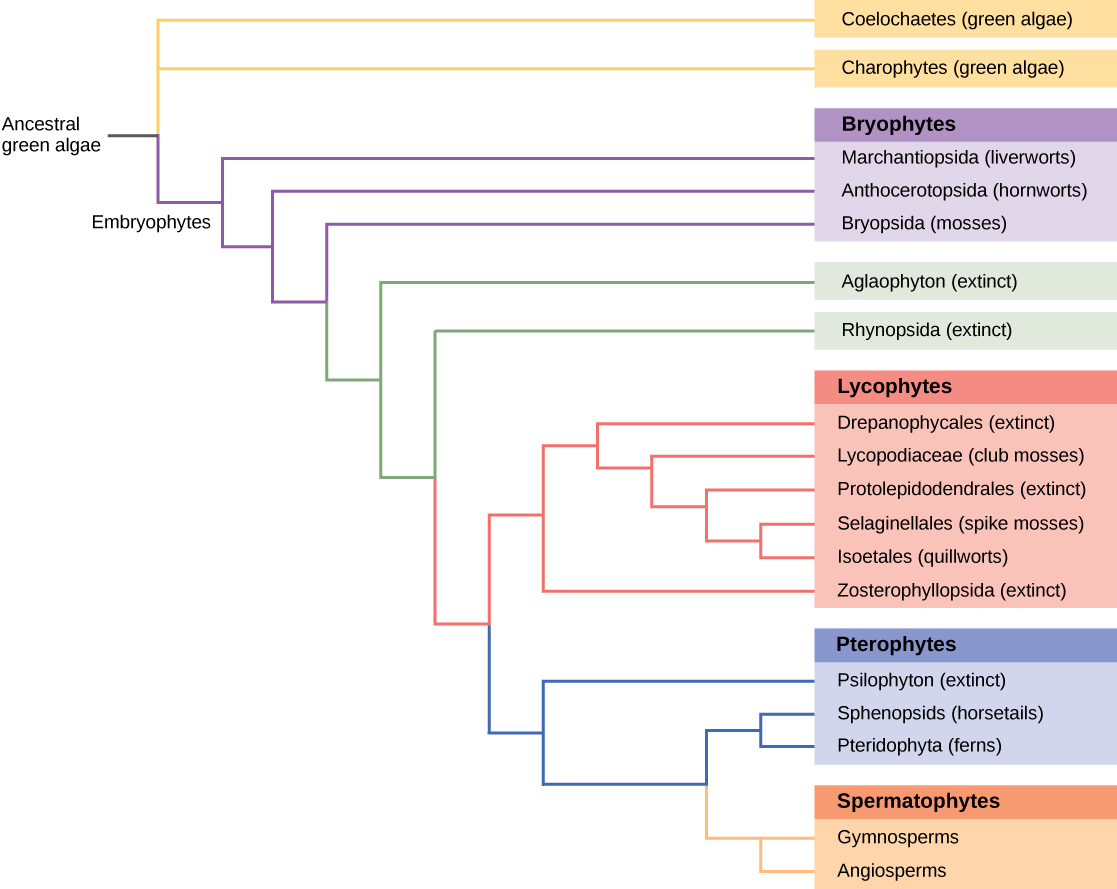
自达尔文时代以来,已经建立了系统发育树来描述物种之间的关系。 传统方法包括比较同源解剖结构和胚胎发育,假设密切相关的生物在胚胎发育过程中具有共同的解剖学特征。 一些在成年体内消失的特征存在于胚胎中;例如,人类胎儿曾经有一条尾巴。 对化石记录的研究显示了将祖先形态与其后代联系起来的中间阶段。 这些方法大多不精确,容易有多种解释。 近年来,随着分子生物学和计算分析工具的开发和完善,新一代的树木建造方法已经形成。 关键假设是,必需蛋白质或RNA结构(例如核糖体RNA)的基因本质上是保守的,因为突变(DNA序列的变化)可能会危及生物体的存活。 来自微量活生物体或化石的DNA可以通过聚合酶链反应(PCR)进行扩增和测序,靶向最有可能在物种之间保守的基因组区域。 经常选择编码来自小型 18S 亚基和质体基因的核糖体 RNA 的基因进行 DNA 比对分析。
获得感兴趣的序列后,将它们与数据库中的现有序列进行比较,例如由国家生物技术信息中心维护的GenBank。 有许多计算工具可用于对齐和分析序列。 复杂的计算机分析程序确定序列一致性或同源性的百分比。 序列同源性可用于估计两个 DNA 序列之间的进化距离,并反映自基因与共同祖先分离以来经过的时间。 分子分析彻底改变了种系发育树。 在某些情况下,形态学研究的先前结果已经得到证实:例如,证实Amborella trichopoda是已知的最原始的被子植物。 但是,由于DNA分析,一些群体和关系已被重新排列。
摘要
种子植物出现在大约一百万年前,即石炭纪时期。 两项重大创新——种子和花粉——允许种子植物在没有水的情况下繁殖。 种子植物的配子体萎缩,而孢子体成为突出的结构,二倍体阶段成为生命周期中最长的阶段。 裸子植物在三叠纪时期成为主导群体。 在这些植物中,花粉颗粒和种子可以防止干燥。 与孢子不同,种子是二倍体胚胎,周围环绕着储藏组织和保护层。 它具有延迟发芽直到生长条件达到最佳状态的能力。 被子植物既有花又有果实。 这些结构在配子和胚胎发育过程中为其提供保护。 被子植物出现在中生代,现已成为陆地栖息地的主要植物生命。
词汇表
- 花
- 在一些育种植物中发现的专门用于繁殖的树枝,含有专门的雄性或雌性器官,或者同时含有雄性和雌性器官
- 水果
- 源自子房壁的增厚组织,可在受精后保护胚胎并促进种子传播
- 胚珠
- 雌配子体
- 花粉颗粒
- 含有植物雄配子体的结构
- 花粉管
- 从花粉颗粒中延伸,将精子输送到卵细胞
- progymnoperm
- 一组类似于针叶树的过渡植物,因为它们生产木材,但仍然像蕨类植物一样繁殖
- 种子
- 包含胚胎、储藏组织和保护外套的结构
- 精子植物
- 种子植物;来自希腊精子(种子)和植物(植物)