
25.1: 早期植物生命
- Page ID
- 203378

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 讨论在陆地上种植生命所面临的挑战
- 描述允许植物在陆地上殖民的适应措施
- 描述植物进化的时间表以及陆地植物对其他生物的影响
Plantae 王国构成了庞大而多样的生物群体。 有超过 300,000 种编目植物。 其中,超过26万株是种子植物。 苔藓、蕨类植物、针叶树和开花植物都是植物界的成员。 大多数生物学家也认为绿藻是植物,尽管其他人则将所有藻类排除在植物界之外。 这种分歧的原因是只有绿藻 Charophytes 与陆地植物具有共同的特征(例如使用叶绿素 a 和 b 加上与植物比例相同的胡萝卜素)。 这些特征在其他类型的藻类中不存在。
进化联系:藻类和光合作用的进化路径
一些科学家认为所有藻类都是植物,而另一些科学家则断言只有夏洛植物属于植物王国。 这些不同的观点与不同类型的藻类选择的不同光合作用进化路径有关。 尽管所有藻类都是光合作用的,也就是说,它们含有某种形式的叶绿体,但它们并不是都通过相同的路径变成光合作用。
大约16.5亿年前,绿藻的祖先通过内共生一种绿色光合细菌而成为光合作用。 那条藻系演变为 Charophytes,最终演变成现代苔藓、蕨类植物、裸子植物和被子植物。 它们的进化轨迹相对直截了当,是单系的。 相比之下,另一种藻类 —— 红色、棕色、金色、stramenopiles 等 —— 都通过次生甚至第三次内共生事件变成了光合作用;也就是说,它们内共生了已经与蓝细菌内共生的细胞。 这些光合作用的后来者在自养方面与 Charophytes 相似,但它们的扩张程度与 Charophytes 不同,也没有在陆地上殖民。
关于是否所有藻类都是 Plantae 的不同看法源于如何看待这些进化路径。 只追踪进化直线(即单系)的科学家,只将 Charophytes 视为植物。 对于那些对具有共同特征的生物(在本例中为光合真核生物)投下广泛网络的生物学家来说,所有藻类都是植物。
链接到学习
访问这个互动网站,更深入地了解 Charophytes。
植物对陆地生活的适应
随着生物适应陆地生活,它们不得不应对陆地环境中的多项挑战。 水被描述为 “生命之物”。 细胞内部是水汤:在这种培养基中,大多数小分子会溶解和扩散,代谢的大多数化学反应都发生了。 对于暴露在空气中的生物来说,干燥或干燥是一种持续的危险。 即使植物的一部分靠近水源,空中结构也可能变干。 水还为生物提供浮力。 在陆地上,植物需要在无法提供与水相同的升力的介质中形成结构支撑。 该生物还会受到诱变辐射的轰击,因为空气无法过滤阳光中的紫外线。 此外,雄配子必须使用新的策略到达雌配子,因为不再可能游泳。 因此,必须保护配子和合子免受干燥。 成功的陆地植物制定了应对所有这些挑战的策略。 并非所有改编作品都同时出现。 有些物种从未远离水生环境,而另一些物种则继续征服地球上最干燥的环境。
为了平衡这些生存挑战,陆地生活具有多种优势。 首先,阳光充足。 水充当过滤器,改变光合色素叶绿素吸收的光谱质量。 其次,二氧化碳在空气中比在水中更容易获得,因为它在空气中的扩散速度更快。 第三,陆地植物在陆地动物之前进化;因此,在陆地被动物殖民之前,没有捕食者威胁植物生命。 随着动物从水中出来并以已建立的植物群中丰富的营养来源为食,这种情况发生了变化。 反过来,植物制定了阻止捕食的策略:从刺和刺到有毒化学物质。
早期的陆地植物,比如早期的陆地动物,生活在离充足水源不远的地方,因此制定了抗旱的生存策略。 其中一种策略叫做宽容。 例如,许多苔藓会变干成棕色而脆的垫子,但是一旦雨水或洪水使水可用,苔藓就会吸收水并恢复健康的绿色外观。 另一种策略是在湿度高的环境中殖民,在这些环境中,干旱并不常见。 蕨类植物被认为是植物的早期谱系,它们在潮湿和凉爽的地方生长,例如温带森林的地下。 后来,植物通过抵抗干燥而不是耐受性而远离潮湿或水生环境。 这些植物,如仙人掌,可以最大限度地减少水分流失,使其能够在极其干燥的环境中生存。
最成功的适应解决方案是开发新的结构,使植物在新的干燥环境中殖民时具有优势。 所有陆生植物都有四种主要适应性:世代交替、孢子在其中形成的孢子、产生单倍体细胞的配子体以及根和芽中的顶端分生组织。 蜡状角质层和含有木质素的细胞壁的进化也为陆地植物的成功做出了贡献。 密切相关的绿藻明显缺乏这些改编,这是关于它们在植物界中的地位的争论的另一个原因。
世代交替
世代交替描述了一个生命周期,在这个周期中,生物体既有单倍体又有二倍体多细胞阶段(图\(\PageIndex{1}\))。
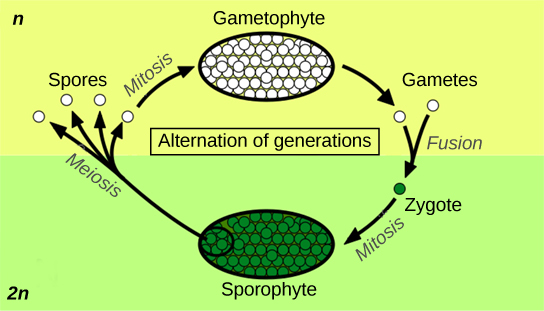
Haplontic 是指存在占主导地位的单倍体阶段的生命周期,而 diplontic 是指以二倍体为主要生命阶段的生命周期。 人类是 diplontic。 大多数植物表现出世代交替,这被描述为单倍体:被称为配子体的单倍体多细胞形式,在发育顺序中紧随其后的是多细胞二倍体生物:孢子体。 配子体通过有丝分裂产生配子(生殖细胞)。 这可能是植物生命周期中最明显的阶段,比如在苔藓中,也可能发生在高等植物(维管植物的常用统称)中的微观结构中,例如花粉颗粒。 孢子体阶段在低层植物(苔藓、肝草和地衣等植物群的统称)中几乎不明显。 高耸的树木是红杉和松树等植物生命周期中的二元化阶段。
保护胚胎是陆地植物的主要要求。 必须保护脆弱的胚胎免受干燥和其他环境危害。 在无核植物和种子植物中,雌配子体在胚胎发育成新一代孢子体时为胚胎提供保护和营养。 陆地植物的这一显著特征使该群体有了胚胎植物的别名。
无核植物中的孢子兰
无核植物的孢子体是二倍体,由两个配子的同婚(融合)产生。 孢子体带有 sporangia(单数,sporangium):最早出现在陆地植物中的器官。 “sporangia” 一词的字面意思是 “血管中的孢子”,因为它是一个含有孢子的生殖囊\(\PageIndex{2}\)。 在多细胞孢子囊内,二倍体孢子细胞或母细胞通过减数分裂产生单倍体孢子,其中 2 n 染色体数量减少到 1 n(请注意,许多植物孢子体是多倍体:例如,硬质小麦是四倍体,面包小麦是六倍体,有些蕨类植物是 1000 倍体)。 孢子后来被孢子囊释放出来并分散在环境中。 陆地植物会产生两种不同类型的孢子,导致在生命周期的不同阶段出现性别分离。 无籽非维管植物只产生一种孢子,被称为同孔。 配子体阶段在这些植物中占主导地位。 从孢子中发芽后,由此产生的配子体会产生雄性和雌性配子体,通常在同一个个体上。 相比之下,异孔植物会产生两种形态上不同类型的孢子。 雄性孢子之所以被称为微孢子,是因为它们的大小较小,会发育成雄性配子体;相对较大的超级孢子会发展成雌配子体。 在一些无籽维管植物和所有种子植物中都观察到异孢子。

当单倍体孢子在热情好客的环境中发芽时,它会通过有丝分裂产生多细胞配子体。 配子体支持配子融合形成的合子和由此产生的幼孢子体(营养形态)。 然后,循环重新开始。
无核植物的孢子周围环绕着厚厚的细胞壁,其中含有一种叫做 s poropollenin 的坚硬聚合物。 这种复杂物质的特征是与脂肪酸和类胡萝卜素相关的长链有机分子:因此,大多数花粉呈黄色。 Sporopollenin 对化学和生物降解具有异常的抵抗力。 在使用花粉将雄性精子转移到雌卵的种子植物中,孢子花素的韧性解释了保存完好的花粉化石的存在。 Sporopollenin 曾经被认为是陆地植物的创新;但是,绿藻 Coleochaetes 会形成含有 sporopollenin 的孢子。
无核植物中的 Gametangia
Gametangia(单数,gametangium)是在多细胞单倍体配子体上观察到的结构。 在 gametangia 中,前体细胞通过有丝分裂产生配子。 雄性 gametangium(antheridium)会释放精子。 许多无核植物会产生带有鞭毛的精子,使它们能够在潮湿的环境中游到 archegonia:雌性 gametangium。 胚胎作为孢子体在 archegonium 内部发育。 Gametangia 在无核植物中很突出,但在种子植物中很少见。
Apical Meristems
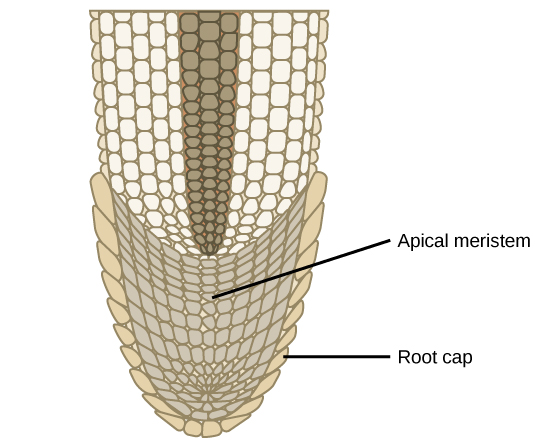
植物的芽和根的长度通过在称为顶端分生体的组织中的快速细胞分裂而增加,顶端分生体是位于芽尖或根尖的一小块细胞(图\(\PageIndex{3}\))。 顶端分生体由未分化的细胞组成,这些细胞在植物的整个生命周期中继续增殖。 分生细胞产生生物体的所有特殊组织。 枝条和根部的伸长使植物能够获得额外的空间和资源:芽是光线,根部是水和矿物质。 一个单独的分生体称为横向分生体,它会产生增加树干直径的细胞。
其他陆地植物改编
随着植物适应旱地并独立于潮湿栖息地中持续存在的水,新的器官和结构出现了。 早期的陆地植物在离地面不超过几英寸的地方生长,正在争夺这些低矮的地垫上的光线。 通过长出枝条并长得更高,单株植物可以捕获更多的光线。 由于空气提供的支撑比水少得多,陆地植物在其茎(以及后来的树干)中掺入了更多的硬分子。 在单细胞藻类等小型植物中,简单的扩散就足以在整个生物体中分配水分和养分。 但是,要使植物进化出更大的形态,血管组织的进化以分配水和溶质是先决条件。 血管系统包含木质部和韧皮部组织。 木质部将从土壤中吸收的水分和矿物质输送到芽中,而韧皮部则将光合作用产生的食物输送到整个植物。 根系进化为吸收土壤中的水分和矿物质,并将越来越高的芽固定在土壤中。
在陆地植物中,一种叫做角质层的蜡质防水覆盖物可以保护叶子和茎免受干燥。 但是,角质层还可以阻止摄入通过光合作用合成碳水化合物所需的二氧化碳。 为了克服这个问题,植物从潮湿的环境转移到更干燥的栖息地时,会出现打开和关闭以调节气体和水蒸气流量的气孔或毛孔。
水过滤紫外线 B (UVB) 光,这对所有生物都有害,尤其是那些必须吸收光才能生存的生物。 这种过滤不适用于陆地植物。 这给陆地定植带来了额外的挑战,合成保护性类黄酮和其他化合物的生物合成途径的演变应对了这一挑战:吸收紫外线波长的光并保护植物空中部分免受光动力学损伤的色素。
植物无法避免被动物吃掉。 相反,它们合成了大量有毒的次生代谢物:复杂的有机分子,例如生物碱,其有毒气味和难闻的味道会阻止动物。 这些有毒化合物还可能导致严重疾病甚至死亡,从而阻碍捕食。 几个世纪以来,人类一直将其中许多化合物用作药物、药物或香料。 相比之下,随着植物与动物共同进化,甜味和营养代谢物的发育诱使动物在分散花粉颗粒、水果或种子方面提供宝贵的帮助。 数亿年来,植物一直在以这种方式招募动物作为它们的助手。
陆地植物的演变
如果不简要回顾地质时代的时间表,就无法讨论陆地上植物的演变。 早期时代被称为古生代,分为六个时期。 它始于寒武纪时期,其次是奥陶纪、志留纪、泥盆纪、石炭纪和二叠纪。 5亿多年前,纪念奥陶纪的重大事件是现代陆地植物的祖先对土地的殖民。 早期陆地植物的化石细胞、角质层和孢子的历史可以追溯到古生代早期的奥陶纪时期。 已知最古老的维管植物是在泥盆纪的沉积物中发现的。 最丰富的信息来源之一是 Rhynie chert,这是在苏格兰瑞尼发现的沉积岩矿床(图\(\PageIndex{4}\)),在那里发现了一些最早的维管植物的嵌入化石。

古植物学家区分了已灭绝的物种(如化石)和仍然活着的现存物种。 已灭绝的维管植物被归类为 zosterophylls 和 trimerophytes,很可能缺少真正的叶子和根,形成了大小与现代苔藓相似的低植被垫,尽管一些 trimetophytes 的高度可能达到一米。 后来的库克索尼亚属在志留纪时期蓬勃发展,已从保存完好的例子中进行了广泛的研究。 Cooksonia 的印记显示了细长的分支茎以看似孢子囊的结尾。 从回收的标本中,无法确定库克索尼亚是否有血管组织。 化石表明,到泥盆纪末期,蕨类植物、马尾草和种子植物居住在景观中,树木和森林正在崛起。 这种茂盛的植被有助于丰富大气中的氧气,使呼吸空气的动物更容易在陆地上殖民。 植物还与真菌建立了早期的共生关系,从而产生了菌根:在这种关系中,细丝的真菌网络提高了植物根系的效率,而植物则为真菌提供了光合作用的副产品。
职业联系:古植物学家
生物如何获得使它们能够在新环境中殖民的特征,以及当代生态系统是如何形成的,是进化的基本问题。 Paleobotany(对灭绝植物的研究)通过分析从实地研究中提取的化石标本来解决这些问题,重构了很久以前消失的生物的形态。 古植物学家通过跟踪植物形态的变化来追踪植物的进化:通过识别具有相同特征的共同祖先来揭示现有植物之间的联系。 该领域旨在寻找过渡物种,弥合现代生物发育道路上的空白。 当生物被困在沉积物或保存其形状的环境中时,就会形成化石。 古植物学家在野外收集化石标本,并将其置于地质沉积物和周围其他化石生物的背景下。 这项活动需要格外小心,以保持精细化石和发现它们的岩石层的完整性。
古植物学最近最激动人心的发展之一是使用分析化学和分子生物学来研究化石。 保存分子结构需要一个没有氧气的环境,因为物质通过微生物的活性氧化和降解取决于其存在。 使用分析化学和分子生物学的一个例子是鉴定齐墩果烷,这是一种能阻止害虫的化合物。 到目前为止,齐墩果烷似乎是开花植物所独有的;但是,它现在已经从二叠纪的沉积物中回收了,比目前给出的第一批开花植物出现的日期要早得多。 古植物学家还可以通过分析和比较灭绝植物的DNA序列与活体和相关生物的DNA序列来研究化石DNA,从而产生大量信息。 通过这种分析,可以为植物谱系建立进化关系。
一些古植物学家对分子化石分析得出的结论持怀疑态度。 例如,感兴趣的化学材料在最初的隔离期间以及进一步的操作中暴露在空气中时会迅速降解。 外来物质(主要来自微生物)污染标本的风险总是很高。 尽管如此,随着技术的不断完善,对化石植物DNA的分析将提供有关植物进化及其适应不断变化的环境的宝贵信息。
陆地植物的主要分部
绿藻和陆地植物被组合成一个叫做 Streptophytina 的 subphylum,因此被称为 Streptophytes。 在进一步划分中,根据血管组织的不存在或存在,陆地植物分为两大类,详见图\(\PageIndex{5}\)。 缺乏维管组织的植物被称为非维管植物,维管组织由专门的细胞组成,用于输送水分和养分。 Liverworts、mosses 和 hornworts 是无籽的非维管植物,很可能出现在陆地植物进化的早期。 维管植物形成了一个传导水和溶质的细胞网络。 第一批维管植物出现在奥陶纪晚期,可能与番茄红素相似,包括棍状苔藓(不要与苔藓混淆)和翼生植物(蕨类植物、马尾和打蛋蕨类植物)。 Lycophytes 和 pterophytes 被称为无核维管植物,因为它们不产生种子。 种子植物或种子植物构成所有现有植物中最大的群体,因此在景观中占据主导地位。 种子植物包括裸子植物,最著名的是产生 “裸种子” 的针叶树(裸子植物),以及所有植物中最成功的开花植物(被子植物)。 被子植物保护在花中央的腔室内的种子;腔室的墙壁后来会长成果实。
艺术连接
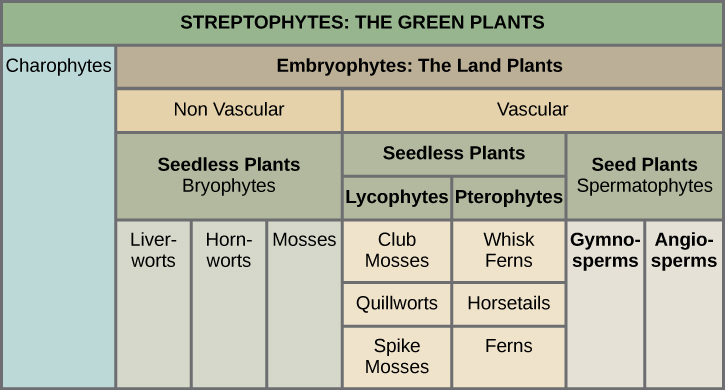
以下关于工厂分部的陈述中哪一项是错误的?
- 番茄红素和紫叶植物是无核维管植物。
- 所有维管植物都会产生种子。
- 所有非血管胚胎植物都是苔藓植物。
- 种子植物包括被子植物和裸子植物。
摘要
陆地植物获得的特征使得在陆地上殖民并在水中生存成为可能。 所有陆地植物都具有以下特征:世代交替,单倍体植物称为配子体,二倍体植物称为孢子体;保护胚胎、在孢子中形成单倍体孢子、在配子体中形成配子,以及顶端分生体。 血管组织、根、叶、角质层覆盖物以及保护孢子的坚硬外层有助于植物适应旱地。 陆地植物出现在大约5亿年前的奥陶纪时期。
艺术联系
图\(\PageIndex{5}\):以下关于工厂分部的陈述中哪一项是错误的?
- 番茄红素和紫叶植物是无核维管植物。
- 所有维管植物都会产生种子。
- 所有非血管胚胎植物都是苔藓植物。
- 种子植物包括被子植物和裸子植物。
- 回答
-
B。
词汇表
- antheridium
- 雄性 gametangium
- archegonium
- 雌性 gametangium
- charophyte
- 绿藻的另一个术语;被认为是陆地植物的最近亲属
- diplontic
- 二倍体阶段是占主导地位的阶段
- 胚胎植物
- 陆地植物的别称;胚胎受到孢子体的保护和滋养
- 现行的
- 仍然活着的物种
- 绝种
- 物种已不复存在
- gametangium
- 产生配子的配子体上的结构
- haplodiplodontic
- 单倍体和二倍体阶段交替出现
- Haplontic
- 单倍体阶段是占主导地位的阶段
- 异孔
- 产生两种类型的孢子
- 同孔的
- 产生一种孢子
- megaspore
- 女性孢子
- microspore
- 雄性孢子
- 非维管植物
- 缺乏血管组织的植物,血管组织由专门的细胞组成,用于输送水分和养分
- 无籽维管植物
- 不产生种子的植物
- 孢子细胞
- 通过减数分裂产生孢子的二倍体细胞
- sporopollenin
- 孢子周围有坚硬的聚合物
- 维管植物
- 含有细胞网络的植物,通过生物体传导水和溶质