
21.1: 病毒进化、形态学和分类
- Page ID
- 203192

培养技能
- 描述病毒最初是如何被发现的,以及它们是如何被检测出来的
- 讨论关于病毒如何进化的三个假设
- 识别病毒的基本形状
- 了解过去和新兴的病毒分类系统
病毒是多种多样的实体。 它们的结构、复制方法和目标主机各不相同。 几乎所有形式的生命——从细菌和古细菌到植物、动物和真菌等真核生物——都有感染它们的病毒。 尽管大多数生物多样性可以通过进化史来理解,例如物种如何适应条件和环境,但有关病毒起源和进化的许多信息仍然未知。
发现和检测
病毒是在开发出一种名为Chamberland-Pasteur过滤器的瓷过滤器之后首次发现的,该过滤器可以从任何液体样本中去除显微镜中可见的所有细菌。 1886年,阿道夫·迈耶(Adolph Meyer)证明,一种烟草植物病,即烟草花叶病,可以通过液体植物提取物从患病植物转移到健康植物。 1892年,德米特里·伊万诺夫斯基表明,即使在张伯兰-巴斯德过滤器从提取物中去除了所有活细菌之后,这种疾病仍可能以这种方式传播。 尽管如此,许多年后才证明这些 “可过滤” 的传染病原体不仅是非常小的细菌,而且是一种新型的非常小的致病颗粒。
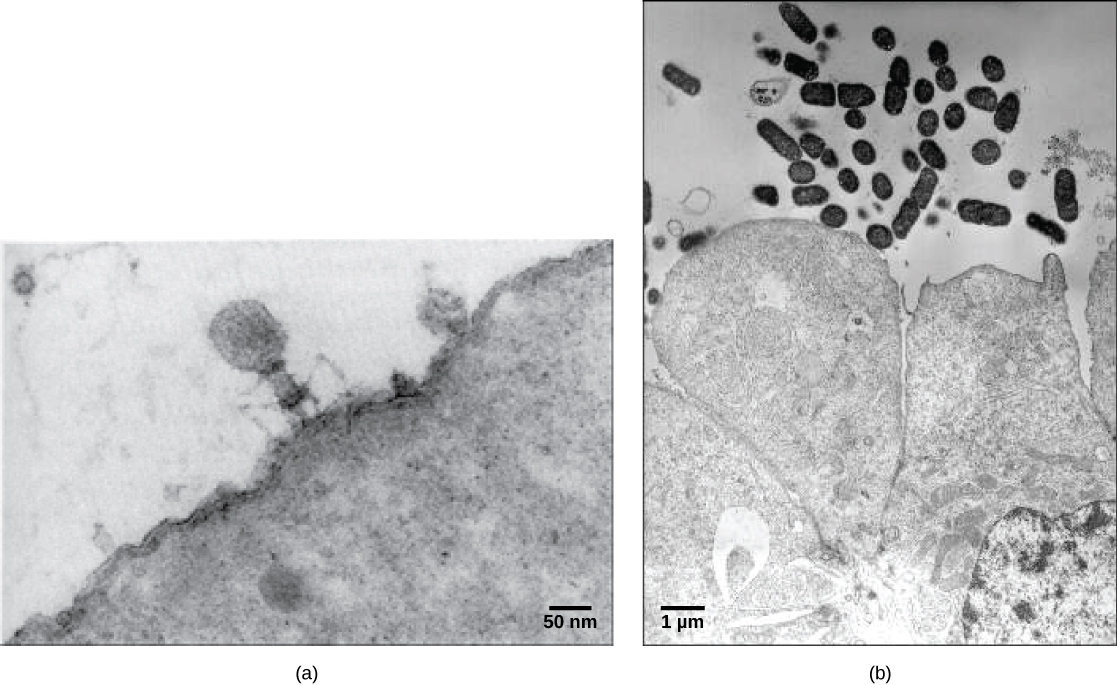
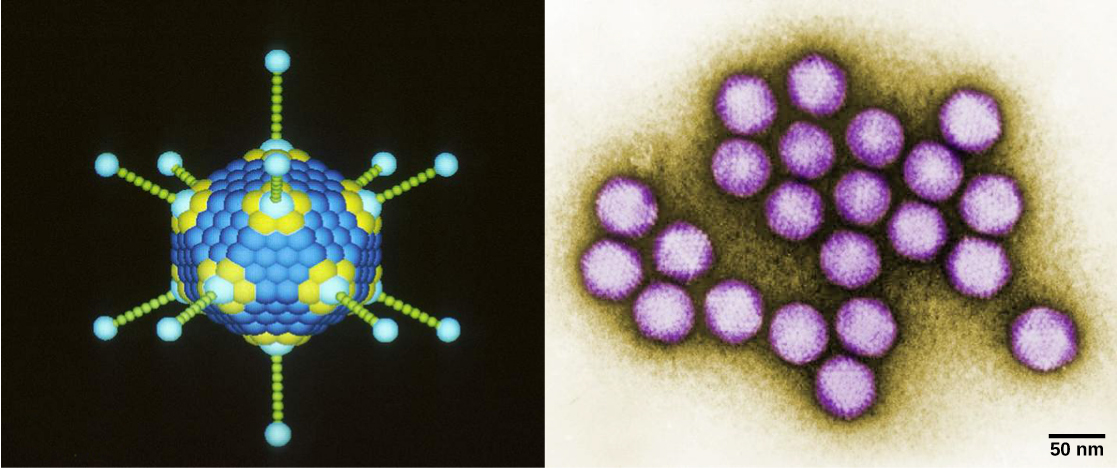
病毒体,单一病毒颗粒,非常小,直径约为20—250纳米。 这些单独的病毒颗粒是宿主细胞外病毒的传染形式。 与细菌(大约大100倍)不同,除了痘病毒家族的一些大型病毒体外,我们无法用光学显微镜看到病毒。 直到 20 世纪 30 年代末电子显微镜的开发后,科学家们才首次很好地了解了烟草花叶病毒(TMV)(图\(\PageIndex{1}\))和其他病毒(图\(\PageIndex{1}\))的结构。 病毒体的表面结构可以通过扫描和透射电子显微镜观察,而病毒的内部结构只能在透射电子显微镜的图像中观察。 这些技术的使用使人们得以发现各种活生物体的许多病毒。 它们最初是按共享形态分组的。 后来,根据病毒所含核酸的类型(DNA或RNA)以及它们的核酸是单链还是双链对病毒组进行了分类。 最近,病毒复制周期的分子分析进一步完善了其分类。
病毒的进化
尽管生物学家已经积累了大量关于当今病毒如何演变的知识,但对病毒最初是如何起源的知之甚少。 在探索大多数生物的进化史时,科学家可以查看化石记录和类似的历史证据。 但是,病毒不会化石,因此研究人员必须通过调查当今的病毒如何演变以及使用生化和遗传信息来创建推测性病毒历史来进行猜测。
尽管大多数发现都认为病毒没有一个共同的祖先,但学者们尚未找到该领域完全接受的关于病毒起源的单一假设。 其中一个假设叫做权力下放或回归假说,它提出通过暗示病毒是由自由生活的细胞进化而来解释病毒的起源。 但是,这个过程可能发生的许多组成部分还是个谜。 第二个假设(称为逃避现实或渐进假设)解释了具有RNA或DNA基因组的病毒,并表明病毒起源于从宿主细胞中逃脱的RNA和DNA分子。 第三种假设假设的自我复制系统与其他自我复制分子的系统相似,很可能与它们作为宿主所依赖的细胞一起进化;对一些植物病原体的研究支持了这一假设。
随着技术的进步,科学家可能会开发和完善进一步的假设来解释病毒的起源。 被称为病毒分子系统学的新兴领域试图通过比较测序的遗传物质来做到这一点。 这些研究人员希望有一天能更好地了解病毒的起源,这一发现可能会导致病毒所产生的疾病的治疗取得进展。
病毒形态学
病毒是脱细胞的,这意味着它们是没有细胞结构的生物实体。 因此,它们缺乏细胞的大部分成分,例如细胞器、核糖体和质膜。 病毒体由核酸核心、外部蛋白质涂层或衣壳组成,有时还包括由源自宿主细胞的蛋白质和磷脂膜制成的外包膜。 病毒还可能含有其他蛋白质,例如酶。 病毒家族成员之间最明显的区别是它们的形态,这种形态非常多样化。 病毒复杂性的一个有趣特征是,宿主的复杂性与病毒体的复杂性无关。 在噬菌体中观察到一些最复杂的病毒体结构,噬菌体是感染最简单的活生物体细菌的病毒。
形态学
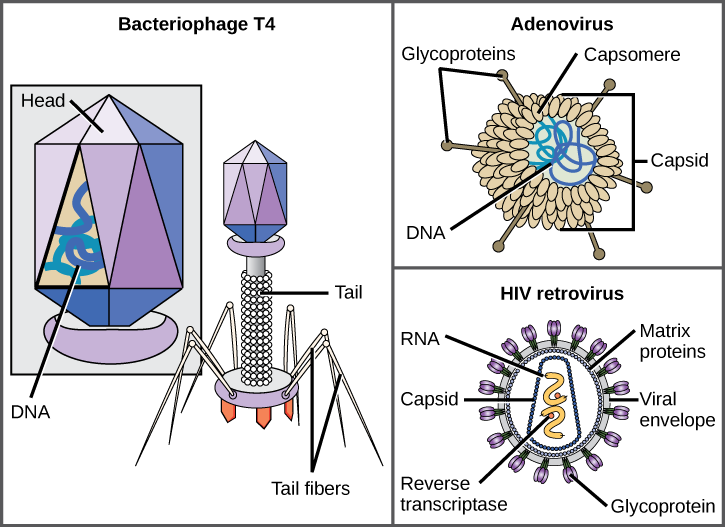
病毒有多种形状和大小,但每个病毒家族的形状和大小都是一致且不同的。 所有病毒体都有一个被称为衣壳的保护层覆盖的核酸基因组。 衣壳由称为 caps omeres 的蛋白质亚基组成。 有些病毒衣壳是简单的多面体 “球体”,而另一些则在结构上相当复杂。
一般而言,病毒的形状分为四组:丝状、等距(或二十面体)、包膜以及头部和尾部。 丝状病毒长而圆柱形。 许多植物病毒是丝状的,包括TMV。 等距病毒的形状大致呈球形,例如脊髓灰质炎病毒或疱疹病毒。 包膜病毒的衣壳周围有膜。 动物病毒,例如艾滋病毒,经常被包裹。 头尾病毒感染细菌,其头部类似于二十面体病毒,尾部形状像丝状病毒。
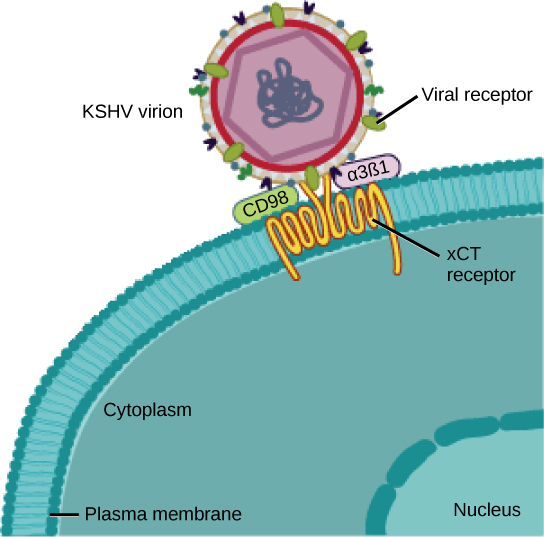
许多病毒使用某种糖蛋白通过细胞上称为病毒受体的分子附着在宿主细胞上(图\(\PageIndex{2}\))。 对于这些病毒来说,附着是日后穿透细胞膜的必要条件,这样它们才能在细胞内完成复制。 病毒使用的受体是通常存在于细胞表面并具有自身生理功能的分子。 病毒只是进化为利用这些分子进行自身复制。 例如,HIV使用T淋巴细胞上的CD4分子作为其受体之一。 CD4是一种称为细胞粘附分子的分子,其作用是在产生T淋巴细胞免疫反应的过程中使不同类型的免疫细胞彼此靠近。
在已知的最复杂的病毒体中,感染大肠埃希氏菌的T4噬菌体具有病毒用来附着在宿主细胞上的尾部结构和容纳其DNA的头部结构。
腺病毒是一种非包膜动物病毒,可导致人类呼吸道疾病,它使用从其胶囊中伸出的糖蛋白尖刺附着在宿主细胞上。 非包膜病毒还包括导致脊髓灰质炎(脊髓灰质炎病毒)、足底疣(乳头瘤病毒)和甲型肝炎(甲型肝炎病毒)的病毒。
像艾滋病的病原体HIV这样的包膜病毒由核酸(HIV的RNA)和被磷脂双层包膜及其相关蛋白包围的衣壳蛋白组成。 嵌入病毒包膜中的糖蛋白用于附着在宿主细胞上。 其他包膜蛋白是稳定包膜的基质蛋白,通常在后代病毒体的组装中发挥作用。 水痘、流感和腮腺炎就是由带有包膜的病毒引起的疾病的例子。 由于包膜的脆弱性,非包膜病毒比包膜病毒更能抵抗温度、pH 值和某些消毒剂的变化。
总体而言,病毒体的形状和包膜的存在与否几乎没有告诉我们病毒可能导致什么疾病或它可能感染什么物种,但它们仍然是开始病毒分类的有用手段(图\(\PageIndex{3}\))。
练习\(\PageIndex{1}\)
以下关于病毒结构的陈述中哪一项是正确的?
- 所有病毒都被包裹在病毒膜中。
- capsomere 由称为衣壳的小蛋白质亚基组成。
- DNA是所有病毒中的遗传物质。
- 糖蛋白帮助病毒附着在宿主细胞上。
- 回答
-
D
核酸的种类
与几乎所有使用DNA作为遗传物质的活生物不同,病毒可能使用DNA或RNA作为其遗传物质。 病毒核心包含病毒的基因组或总遗传含量。 病毒基因组往往很小,只包含那些编码病毒无法从宿主细胞获得的蛋白质的基因。 这种遗传物质可以是单链或双链。 它也可以是线性的或圆形的。 虽然大多数病毒含有单个核酸,但其他病毒的基因组有多个核酸,称为片段。
在 DNA 病毒中,病毒 DNA 引导宿主细胞的复制蛋白合成病毒基因组的新拷贝,并将该基因组转录并转化为病毒蛋白。 DNA病毒会导致人类疾病,例如水痘、乙型肝炎和一些性病,例如疱疹和尖锐湿疣。
RNA 病毒仅含有 RNA 作为其遗传物质。 为了在宿主细胞中复制其基因组,RNA病毒对可以将RNA复制到DNA中的酶进行编码,而宿主细胞无法做到这一点。 这些 RNA 聚合酶比 DNA 聚合酶更容易出现复制错误,因此在转录过程中经常会出错。 因此,RNA病毒的突变比在DNA病毒中更频繁地发生。 这使他们能够更快地改变和适应宿主。 由RNA病毒引起的人类疾病包括丙型肝炎、麻疹和狂犬病。
病毒分类
要了解不同病毒组之间的共同特征,必须制定分类方案。 但是,由于人们认为大多数病毒不是从共同的祖先演变而来的,因此科学家用来对生物进行分类的方法并不是很有用。 过去,生物学家根据不同病毒的形态和遗传学使用了多种分类系统。 但是,这些早期的分类方法根据病毒的哪些特征对病毒进行分类,对病毒进行了不同的分组。 当今最常用的分类方法称为巴尔的摩分类方案,它基于每种特定类型的病毒中信使RNA(mRNA)的生成方式。
过去的分类系统
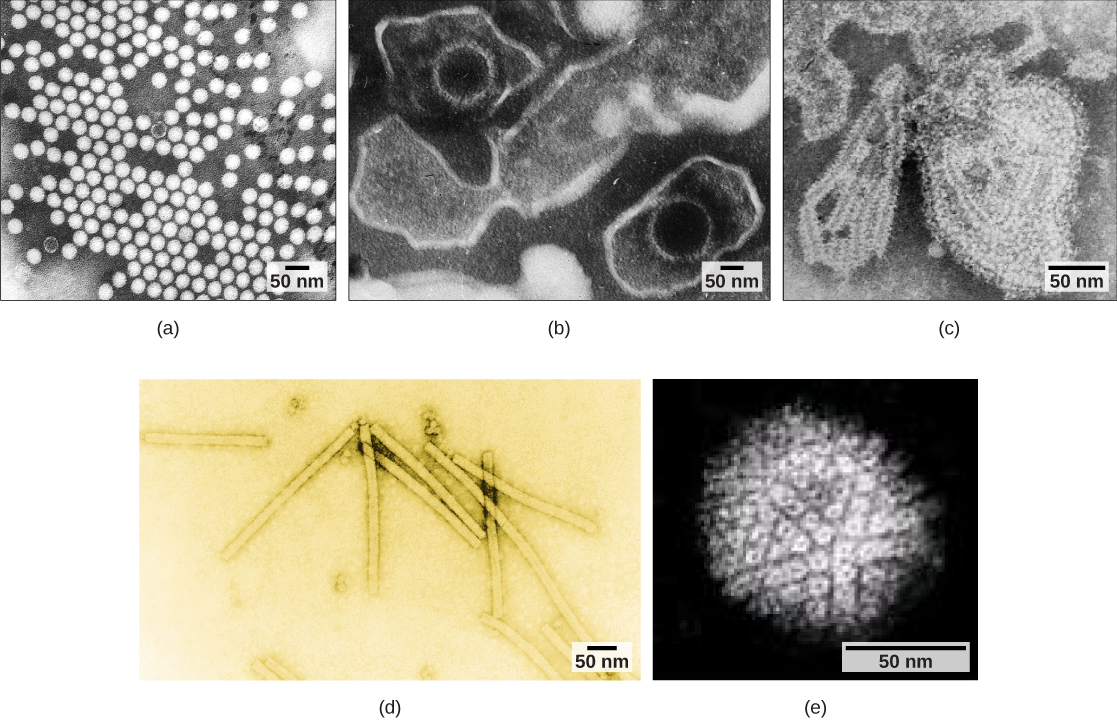
病毒可通过多种方式进行分类:按核心含量(表\(\PageIndex{1}\)和图\(\PageIndex{2}\))、衣壳结构以及是否有外包膜等因素进行分类。 遗传物质(DNA 或 RNA)的类型及其结构(单链或双链、线性或圆形、分段或非分段)用于对病毒核心结构进行分类。
| 核心分类 | 示例 |
|---|---|
|
|
|
|
|
|
|
|
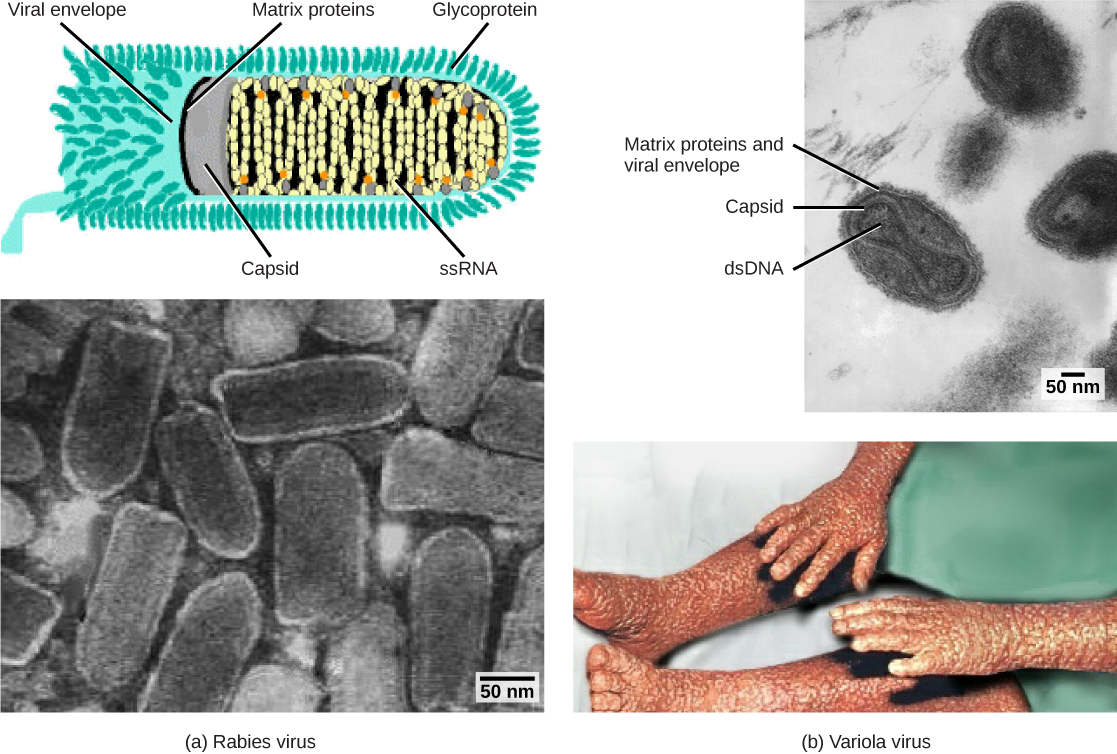
病毒也可以根据其衣壳的设计进行分类(图\(\PageIndex{3}\)和图\(\PageIndex{4}\))。 衣壳分为裸二十面体、包封的二十面体、包络的螺旋、裸螺旋和复杂(图\(\PageIndex{5}\)和图\(\PageIndex{6}\))。 遗传物质(DNA或RNA)的类型及其结构(单链或双链、线性或圆形、分段或非分段)用于对病毒核心结构进行分类(表\(\PageIndex{2}\))。
| 衣壳分类 | 示例 |
|---|---|
| 裸体二十面体 | 甲型肝炎病毒、脊髓灰质炎病毒 |
| 封闭的二十面体 | 爱泼斯坦-巴尔病毒、单纯疱疹病毒、风疹病毒、黄热病病毒、HIV-1 |
| 封装螺旋线 | 流感病毒、流行性腮腺炎病毒、麻疹病毒、狂犬病毒 |
| 裸螺旋 | 烟草花叶病毒 |
| 与许多蛋白质复合;有些具有二十面体和螺旋衣壳结构的组合 | 疱疹病毒、天花病毒、乙型肝炎病毒、T4 噬菌体 |
巴尔的摩分类
最常用的病毒分类系统是由诺贝尔奖得主生物学家戴维·巴尔的摩在20世纪70年代初开发的。 除了上述形态学和遗传学的差异外,巴尔的摩分类方案还根据病毒复制周期中mRNA的产生方式对病毒进行分组。
第一组病毒以双链 DNA (dsDNA) 作为其基因组。 它们的 mRNA 是通过转录产生的,其产生方式与细胞 DNA 大致相同。 第二组病毒以单链 DNA (ssDNA) 作为其基因组。 在转录为 mRNA 之前,他们将其单链基因组转化为 dsDNA 中间体。 第三组病毒使用 dsRNA 作为其基因组。 这些链分离,其中一条用作使用病毒编码的依赖RNA的RNA聚合酶生成mRNA的模板。 IV 组病毒以 ssRNA 作为其基因组,具有正极性。 正极性意味着基因组RNA可以直接用作mRNA。 dsRNA 的中间体,称为复制中间体,是在复制基因组 RNA 的过程中产生的。 由这些中间体形成多条负极性(与正链基因组RNA互补)的全长RNA链,然后这些中间体可以作为产生具有正极性的RNA的模板,包括全长基因组RNA和较短的病毒mRNA。 V 组病毒包含极性为负的 ssRNA 基因组,这意味着它们的序列与 mRNA 互补。 与 IV 组病毒一样,dsRNA 中间体用于复制基因组并产生 mRNA。 在这种情况下,负链基因组可以直接转化为 mRNA。 此外,全长阳性 RNA 链可用作生成负链基因组的模板。 第六组病毒有二倍体(两个拷贝)ssRNA 基因组,必须使用逆转录酶将这些基因组转化为 dsDNA;然后 dsDNA 被运送到宿主细胞核并插入宿主基因组。 然后,可以通过转录整合到宿主基因组中的病毒 DNA 来产生 mRNA。 VII 组病毒具有部分 dsDNA 基因组,可制成 ssRNA 中间体,充当 mRNA,但也可以通过逆转录酶转回 dsDNA 基因组,这是基因组复制所必需的。 表中总结了巴尔的摩分类中每个组的特征,并\(\PageIndex{3}\)附有每个组的示例。
| 群组 | 特征 | mRNA 的产生模式 | 示例 |
|---|---|---|---|
| 我 | 双链 DNA | mRNA 直接从 DNA 模板中转录 | 单纯疱疹(疱疹病毒) |
| 二 | 单链 DNA | 在 RNA 被转录之前,DNA 被转化为双链形式 | 犬细小病毒(细小病毒) |
| 三 | 双链 RNA | mRNA 是从 RNA 基因组中转录的 | 儿童胃肠炎(轮状病毒) |
| IV | 单链 RNA (+) | 基因组起到 mRNA 的作用 | 普通感冒(pircornavirus) |
| V | 单链 RNA (-) | mRNA 是从 RNA 基因组中转录的 | 狂犬病(rhabdovirus) |
| 六 | 含有逆转录酶的单链 RNA 病毒 | 逆转录酶从 RNA 基因组中产生 DNA;然后 DNA 被纳入宿主基因组;mRNA 从掺入的 DNA 中转录 | 人类免疫缺陷病毒(HIV) |
| 七 | 含有逆转录酶的双链 DNA 病毒 | 病毒基因组是双链 DNA,但病毒 DNA 是通过 RNA 中间体复制的;RNA 可以直接作为 mRNA 或作为 mRNA 的模板 | 乙型肝炎病毒(肝炎病毒) |
摘要
病毒是微小的脱细胞实体,通常只能用电子显微镜看到。 它们的基因组包含 DNA 或 RNA(绝不会两者兼而有之),它们使用宿主细胞的复制蛋白进行复制。 病毒多种多样,感染古细菌、细菌、真菌、植物和动物。 病毒由核酸核心组成,周围环绕着带有或没有外部脂质包膜的蛋白质衣壳。 衣壳形状、包膜的存在和核心成分决定了病毒分类的某些要素。 最常用的分类方法是巴尔的摩分类,它根据病毒产生 mRNA 的方式对病毒进行分类。
词汇表
- 脱细胞的
- 缺少细胞
- 衣壳
- 病毒核心的蛋白质涂层
- capsomere
- 构成衣壳的蛋白质亚单位
- 信封
- 包裹某些病毒的脂质双层
- I 组病毒
- 带有 dsDNA 基因组的病毒
- II 组病毒
- 带有 ssDNA 基因组的病毒
- 第三组病毒
- 带有 dsRNA 基因组的病毒
- IV 组病毒
- 具有正极性 ssRNA 基因组的病毒
- V 组病毒
- 带有负极性 ssRNA 基因组的病毒
- VI 组病毒
- 带有 ssRNA 基因组的病毒通过逆转录酶转化为 dsDNA
- VII 组病毒
- 带有单链 mRNA 的病毒转化为 dsDNA 以进行基因组复制
- 基质蛋白
- 可稳定包膜的包膜蛋白,通常在后代病毒体的组装中发挥作用
- 负极性
- 具有与其 mRNA 互补基因组的 ssRNA 病毒
- 正极性
- ssRNA 病毒,其基因组包含与其 mRNA 中发现的相同碱基序列和密码子
- 复制中间体
- 在复制基因组 RNA 过程中制造的 dsRNA 中间体
- 逆转录酶
- 在巴尔的摩第 VI 组和 VII 组中发现的将单链 RNA 转化为双链 DNA 的酶
- 病毒受体
- 糖蛋白过去通过细胞上的分子将病毒附着在宿主细胞上
- 病毒体
- 宿主细胞外的单个病毒颗粒
- 病毒核心
- 包含病毒基因组