
20.3: 关于系统发育树的观点
- Page ID
- 202187

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 描述水平基因转移
- 说明原核生物和真核生物如何水平转移基因
- 识别系统发育关系的网络和环形模型,并描述它们与最初的系统发育树概念有何不同
系统发育建模的概念在不断变化。 它是所有生物学中最具活力的研究领域之一。 在过去的几十年中,新的研究挑战了科学家关于生物之间关系的观念。 已经提出了这些关系的新模式供科学界考虑。
许多系统发育树已被证明是物种之间进化关系的模型。 系统发育树起源于查尔斯·达尔文(Charles Darwin),他在1837年绘制了第一棵系统发育树(图\(\PageIndex{1}\) a),这是一个多世纪以来的后续研究模式。 系统发育树的概念,其单个树干代表共同祖先,树枝代表物种与该祖先的差异,这与许多常见树木的结构非常吻合,例如橡树(图\(\PageIndex{1}\) b)。 但是,来自现代DNA序列分析和新开发的计算机算法的证据引起了科学界对标准树模型有效性的怀疑。

经典模型的局限性
经典树模型中包含的关于原核生物进化的经典思维是,物种是克隆进化的。 也就是说,它们自己只通过随机突变产生后代,从而下降到科学界已知的现代灭绝物种中。 这种观点在有性繁殖的真核生物中有些复杂,但孟德尔遗传学定律再次将后代的变异解释为物种内部突变的结果。 直到最近,基因在不相关的物种之间转移的概念才被认为是可能的。 水平基因转移(HGT),也称为横向基因转移,是指不相关物种之间的基因转移。 事实证明,HGT 是一种永远存在的现象,许多进化论者假设这个过程在进化中起着重要作用,因此使简单的树模型变得复杂。 事实证明,使用标准系统发育,基因可以在仅有远距离关联的物种之间传递,从而使对系统发育关系的理解增加了一层复杂性。
HGT 在原核生物中出现的各种方式对于理解系统发育非常重要。 尽管目前人们认为 HGT 对真核生物进化并不重要,但 HGT 也出现在这个领域。 最后,作为终极基因转移的一个例子,有人提出了共生或内共生生物之间的基因组融合理论,以解释一个非常重要的事件——第一个真核细胞的进化,没有这个真核细胞,人类就不可能存在。
水平基因转移
水平基因转移(HGT)是指通过其他机制将遗传物质从一个物种引入另一个物种,而不是从亲本到后代的垂直传播。 这些转移甚至允许远距离相关的物种共享基因,从而影响其表型。 人们认为HGT在原核生物中更为普遍,但只有大约2%的原核生物基因组可以通过这种过程转移。 一些研究人员认为,这样的估计还为时过早:必须将HGT对进化过程的实际重要性视为一项正在进行的工作。 随着对这种现象的调查更加彻底,可以发现这种现象更为常见。 许多科学家认为,HGT 和突变似乎是(尤其是在原核生物中)遗传变异的重要来源,而遗传变异是自然选择过程的原料。 这些转移可能发生在任何两个有亲密关系的物种之间(表\(\PageIndex{1}\))。
表\(\PageIndex{1}\):原核生物和真核生物 HGT 机制摘要
| 机制 | 传输方式 | 示例 | |
|---|---|---|---|
| 原核生物 | 转型 | DNA 吸收 | 许多原核生物 |
| 转导 | 噬菌体(病毒) | 细菌 | |
| 共轭 | pilus | 许多原核生物 | |
| 基因转移剂 | 噬菌体样颗粒 | 紫色非硫细菌 | |
| 真核生物 | 来自食物生物 | 未知 | 蚜虫 |
| 跳跃基因 | 转座子 | 水稻和小米植物 | |
| 附生植物/寄生虫 | 未知 | 紫杉树真菌 | |
| 来自病毒感染 |
原核生物中的 HGT
事实证明,HGT 的机制在细菌和古细菌的原核结构域中非常常见,极大地改变了人们对它们进化的看法。 大多数进化模型,例如Endosymbiont理论,都认为真核生物来自多个原核生物,这使得HGT对于理解所有现存和灭绝物种的系统发育关系变得更加重要。
基因在普通细菌之间转移的事实已为微生物学专业的学生所熟知。 物种之间的这些基因转移是细菌获得抗生素耐药性的主要机制。 通常,人们认为这种类型的转移是由三种不同的机制发生的:
- 转化:裸露的 DNA 被细菌吸收
- 转导:使用病毒转移基因
- 共轭:使用一种叫做 pilus 的空心管在生物体之间转移基因
最近,发现了原核生物之间基因转移的第四种机制。 称为基因转移剂 (GTA) 的小病毒样颗粒将随机基因组片段从一种原核生物转移到另一种原核生物。 事实证明,GTA是造成遗传变化的原因,与其他进化过程相比,有时频率很高。 第一个 GTA 的特征是在 1974 年使用紫色的非硫细菌。 这些 GTA 被认为是丧失自身繁殖能力的噬菌体,它们将随机的 DNA 片段从一个生物体携带到另一个生物体。 在使用海洋细菌的对照研究中,GTA具有高频作用的能力。 据估计,仅在地中海,海洋原核生物中的基因转移事件,无论是GTA还是病毒,每年就高达10 13 次。 GTA 和病毒被认为是高效的 HGT 载体,对原核生物进化有重大影响。
由于这种现代DNA分析,真核生物直接从古细菌进化的想法已经失宠了。 尽管真核生物具有许多细菌所不具备的特征,例如TATA盒(存在于许多基因的启动子区域),但发现某些真核生物基因与细菌DNA比古细菌DNA更同源,这使得这个想法不那么站得住脚。 此外,有人提出,通过内共生融合来自古细菌和细菌的基因组是真核生物进化的终极事件。
真核生物中的 HGT
尽管很容易看出原核生物是如何通过 HGT 交换遗传物质的,但最初人们认为真核生物中不存在这种过程。 毕竟,原核生物不过是直接暴露于环境中的单细胞,而多细胞生物的性细胞通常被隔离在人体的受保护部位。 从这个想法可以看出,多细胞真核生物之间的基因转移应该更加困难。 事实上,人们认为这种过程在真核生物中更为罕见,其进化影响也比在原核生物中小得多。 尽管如此,已经在几种真核生物中证明了远距离相关的生物之间的 HGT,将来有可能发现更多的例子。
在植物中,已经观察到无法通过正常方式进行异花授粉的物种的基因转移。 转座子或 “跳跃基因” 已被证明可以在水稻和小米植物物种之间转移。 此外,以紫杉树为食的真菌物种(抗癌药物 TAXOL® 源自树皮)已经获得了自己制造紫杉醇的能力,这是基因转移的明显例子。
在动物体内,HGT 的一个特别有趣的例子出现在蚜虫物种中(图\(\PageIndex{2}\))。 蚜虫是根据类胡萝卜素含量不同颜色的昆虫。 类胡萝卜素是由各种植物、真菌和微生物产生的色素,它们在动物身上起着多种作用,动物从食物中获取这些化学物质。 人类需要类胡萝卜素来合成维生素A,而我们通过吃橙色水果和蔬菜来获得它们:胡萝卜、杏子、芒果和红薯。 另一方面,蚜虫已经获得了自己制造类胡萝卜素的能力。 根据DNA分析,这种能力是由于HGT将真菌基因转移到昆虫体内,大概是因为昆虫食用真菌作为食物。 一种叫做去饱和酶的类胡萝卜素酶是某些蚜虫体内出现红色的原因,而且研究进一步表明,当这种基因因突变而失活时,蚜虫会恢复到更常见的绿色(图\(\PageIndex{2}\))。

基因组融合与真核生物的进化
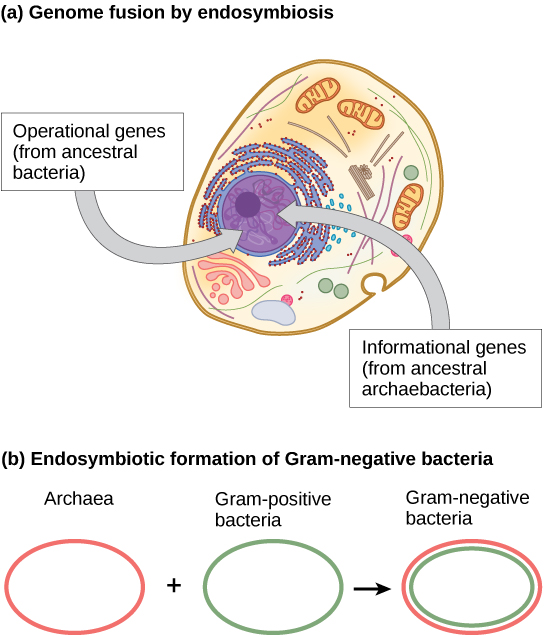
科学家们认为,当两种共生生物变成内共生生物时,HGT 的终极目标发生在不同种类的原核生物之间的基因组融合。 当一个物种被提取到另一个物种的细胞质中时,就会发生这种情况,这最终会产生一个由来自内共生体和宿主的基因组成的基因组。 这种机制是 Endosymbiont 理论的一个方面,大多数生物学家都认为该理论是真核细胞获得线粒体和叶绿体的机制。 但是,内共生在细胞核发育中的作用更具争议性。 核DNA和线粒体DNA被认为具有不同(单独的)进化起源,线粒体DNA来自被古代原核细胞吞没的细菌的循环基因组。 线粒体DNA可以被认为是最小的染色体。 有趣的是,线粒体DNA只能从母亲那里继承。 当精子在受精卵中降解时,或者在其他情况下,当位于精子鞭毛中的线粒体无法进入卵子时,线粒体 DNA 会在精子中降解。
在过去的十年中,加州大学洛杉矶分校/美国宇航局天体生物学研究所的詹姆斯·莱克提出了通过内共生进行基因组融合的过程,该过程负责第一批真核细胞的进化(图\(\PageIndex{3}\) a)。 他的实验室使用DNA分析和一种称为条件重建(CR)的新数学算法,提出真核细胞是由两个物种之间的内共生基因融合而成的,一个是古细菌,另一个是细菌。 如前所述,一些真核生物基因与古细菌的基因相似,而另一些则类似于来自细菌的真核基因。 像莱克所提出的那样,内共生聚变事件将清楚地解释这一观察。 另一方面,这项工作是新的,CR算法相对没有得到证实,这使许多科学家抵制这一假设。
莱克最近的研究(图\(\PageIndex{3}\) b)提出,革兰氏阴性细菌在其结构域中是独一无二的,因为它们含有两个脂质双层膜,确实是由古生物和细菌物种的内共生融合产生的。 双膜将是 endosymbiosis 的直接结果,endosymbiont 在宿主内部化时会从宿主那里吸收第二层膜。 这种机制也被用来解释线粒体和叶绿体中发现的双膜。 莱克的作品并非没有疑问,生物科学界仍在争论这些想法。 除了莱克的假设外,关于真核生物的起源,还有其他几种相互竞争的理论。 真核细胞核是如何进化的? 一种理论认为,原核细胞在细菌染色体周围产生了额外的膜。 有些细菌的DNA被两个膜包围;但是,没有证据表明存在核仁或核孔。 其他变形杆菌也有膜结合的染色体。 如果真核核以这种方式进化,我们预计这两种原核生物中的一种与真核生物的关系会更加密切。
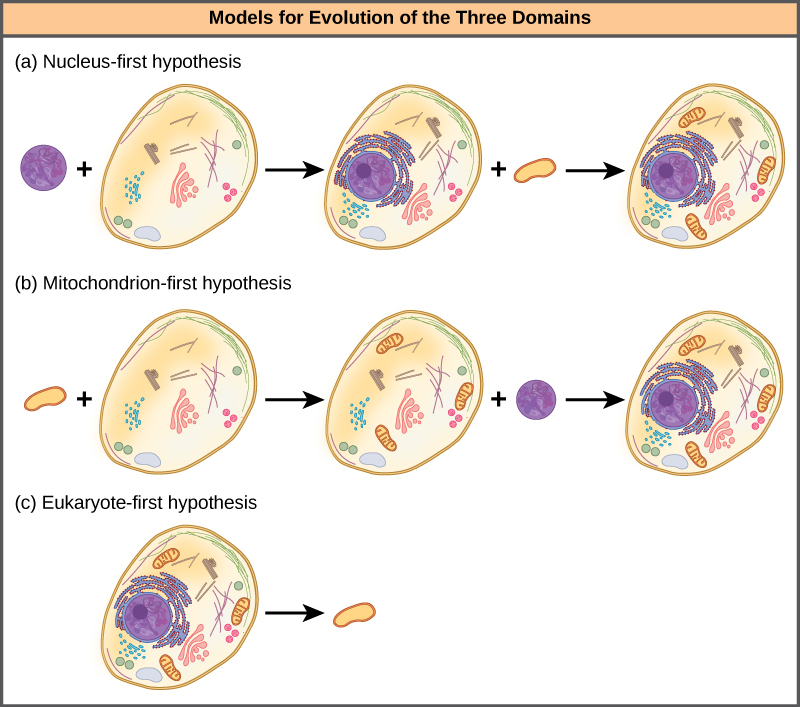
核优先假设提出,原子核首先在原核生物中进化(图\(\PageIndex{4}\) a),然后是新的真核生物与成为线粒体的细菌融合。 线粒体优先假设提出,线粒体最初是在原核生物宿主中建立的(图\(\PageIndex{4}\) b),该宿主随后通过聚变或其他机制获得了一个核,成为第一个真核细胞。 最有趣的是,真核生物优先假设提出,原核生物实际上是通过失去基因和复杂性而从真核生物进化而来的(图\(\PageIndex{4}\) c)。 所有这些假设都是可以检验的。 只有时间和更多的实验才能确定哪种假设最能得到数据的支持。
网络和网络模型
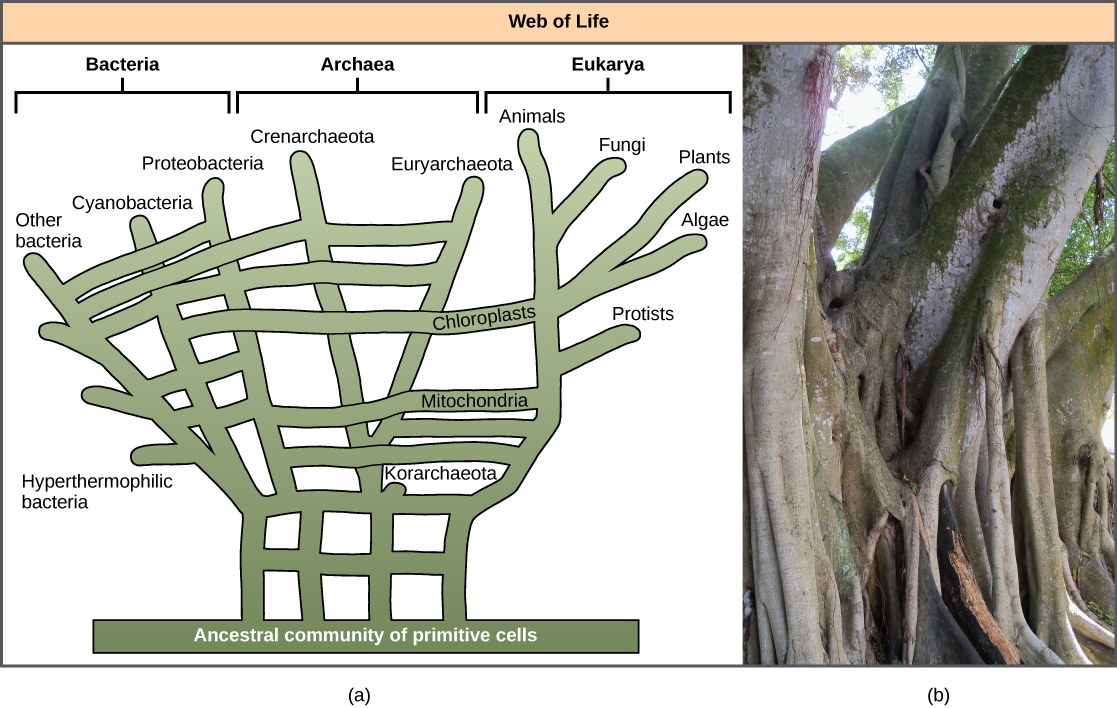
由于认识到HGT的重要性,尤其是在原核生物进化中的重要性,一些人提议放弃经典的 “生命之树” 模型。 1999 年,W. Ford Doolittle 提出了一种系统发育模型,它比树更像网络或网络。 假设真核生物不是从单一的原核生物祖先进化而来的,而是从许多通过HGT机制共享基因的物种库中进化而来的。 如图\(\PageIndex{5}\) a 所示,一些原核生物负责将导致线粒体发育的细菌转移到新的真核生物身上,而其他物种则将产生叶绿体的细菌转移到新真核生物身上。 这种模型通常被称为 “生命之网”。 为了挽救树的比喻,有人提议使用榕树(图\(\PageIndex{5}\) b)及其多根树干作为系统发育来表示 HGT 进化作用的减弱。
生命之环模特
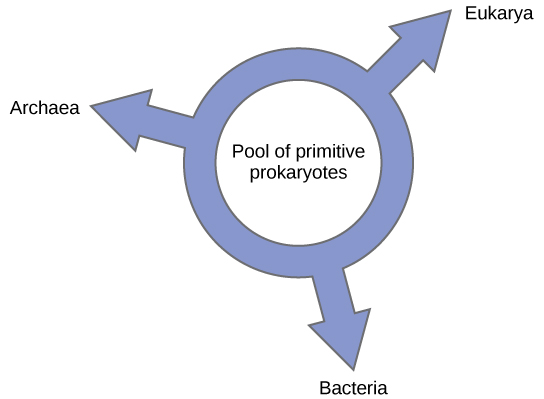
其他人则提议放弃任何树状的系统发育模型,转而采用环状结构,即所谓的 “生命之环”(图\(\PageIndex{6}\));一种系统发育模型,其中所有三个生命领域都是从原始原核生物池进化而来的。 莱克再次使用条件重建算法,提出了一种环状模型,其中所有三个结构域(古细菌、细菌和真核生物)的物种都是从单一的基因交换原核生物池进化而来的。 他的实验室提出,这种结构最适合在他的实验室中进行的大量DNA分析得出的数据,而环模型是唯一充分考虑HGT和基因组融合的模型。 但是,其他系统发育学家仍然对这个模型持高度怀疑态度。
总而言之,必须修改达尔文提出的 “生命之树” 模型,以包括HGT。 这是否意味着完全放弃树模型? 甚至莱克也认为,应该尽一切努力发现对树模型进行一些修改,使其能够准确地拟合他的数据,只有做不到这一点才会促使人们转向他的戒指提案。
这并不意味着树、网或环将与生命的系统发育关系的准确描述完全相关。 关于系统发育模型的新思维的一个结果是,人们认为达尔文最初对系统发育树的概念过于简单,但根据当时已知的情况是有道理的。 但是,寻找更有用的模型的工作仍在继续:每个模型都是需要测试的假设,有可能开发新模型。 科学就是这样发展的。 这些模型用作可视化,以帮助构建假设的进化关系并了解正在分析的大量数据。
摘要
达尔文首次使用的系统发育树是描述物种间系统发育关系的经典 “生命之树” 模型,也是当今最常用的模型。 关于HGT和基因组融合的新想法促使一些人建议修改模型,使其类似于网络或环。
词汇表
- 真核生物优先假设
- 关于原核生物是从真核生物进化而来的提议
- 基因转移剂 (GTA)
- 将随机基因组片段从一种原核生物转移到另一种原核生物的样细菌噬菌体颗粒
- 基因组融合
- 两个原核生物基因组的融合,大概是通过内共生作用进行的
- 水平基因转移 (HGT)
- (也包括横向基因转移)不相关物种之间的基因转移
- 线粒体优先假设
- 提议原核生物先获得线粒体,然后再进行核开发
- 原子核优先假设
- 提议原核生物先获得原子核,然后获得线粒体
- 生命之戒
- 系统发育模型,其中生命的所有三个领域都是从原始原核生物池进化而来的
- 生命之网
- 系统发育模型试图整合水平基因转移对进化的影响