
18.2: 新物种的形成
- Page ID
- 202142

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 定义物种并描述如何将物种识别为不同物种
- 描述导致物种形成的遗传变量
- 识别 prezygotic 和 postzygotic 生殖屏障
- 解释同种异体和同情物种形成
- 描述自适应辐射
尽管地球上的所有生命都有不同的遗传相似之处,但只有某些生物通过有性繁殖将遗传信息结合起来,其后代可以成功繁殖。 科学家称此类生物为同一个生物物种的成员。
物种和繁殖能力
物种是一组单个生物,它们杂交并产生可育的、可存活的后代。 根据这个定义,当在自然界中,每个物种的个体之间的交配不可能产生可育的后代时,一个物种就会与另一个物种区分开来。
同一物种的成员具有外部和内部特征,这些特征是从它们的DNA中发展而来的。 两种生物之间的关系越密切,它们共有的DNA就越多,就像人及其家庭一样。 人们的DNA可能更像他们父亲或母亲的DNA,而不是表亲或祖父母的DNA。 同一物种的生物具有最高的 DNA 对齐水平,因此具有共同的特征和行为,可以成功繁殖。
物种的外观在暗示交配能力或无法交配方面可能具有误导性。 例如,尽管家犬(Canis lupus familiaris)表现出表型差异,例如大小、身材和外套,但大多数狗可以杂交并产生可以成熟和性繁殖的可行幼犬(图\(\PageIndex{1}\))。

在其他情况下,尽管个体不是同一个物种的成员,但它们可能看起来相似。 例如,尽管白头鹰(Haliaeetus leucocephalus)和非洲鱼鹰(Haliaeetus vocifer)既是鸟又是鹰,但它们都属于一个单独的物种组(图\(\PageIndex{2}\))。 如果人类进行人为干预,用非洲鱼鹰的精子使白头鹰的卵受精,而小鸡确实孵化了,那么被称为杂种(两个物种之间的杂交)的后代可能不孕——在成熟后无法成功繁殖。 不同的物种可能具有不同的活跃发育基因;因此,可能无法通过两组不同的方向培育出可存活的后代。 因此,尽管可能发生杂交,但这两个物种仍然分开。

物种群共享一个基因库:物种中所有基因变异的集合。 同样,生物群体或种群发生任何变化的基础都必须是遗传的,因为这是共享和传递特征的唯一途径。 当一个物种内部发生变异时,它们只能通过两个主要途径传给下一代:无性繁殖或有性生殖。 只要繁殖细胞具有改变的特征,这种变化就会以无性方式传播。 为了使改变的特征通过有性生殖传播,配子,例如精子或卵细胞,必须具有改变的特征。 换句话说,有性繁殖的生物体可以在其体内细胞中经历多种遗传变化,但是如果这些变化没有发生在精子或卵细胞中,那么改变的特征将永远不会传给下一代。 只有可遗传的特征才能进化。 因此,繁殖对于遗传变化在种群或物种中扎根起着至关重要的作用。 简而言之,生物必须能够相互繁殖才能将新的特征传递给后代。
形态形成
物种的生物学定义适用于有性繁殖的生物,是一组实际或可能杂交的个体。 此规则有例外。 许多物种非常相似,因此杂交后代是可能的,并且可能经常出现在自然界中,但是对于大多数物种来说,这条规则通常成立。 实际上,自然界中存在相似物种之间的杂交表明它们可能是单一杂交物种的后代,物种形成过程可能尚未完成。
鉴于地球上生命的非凡多样性,必须有物种形成机制:从一个原始物种中形成两个物种。 达尔文将这个过程设想为分支事件,并在《论物种起源》(图\(\PageIndex{3}\) a)中找到的唯一插图中描绘了这个过程。 将此插图与大象进化图(图\(\PageIndex{3}\) b)进行比较,后者显示,当一个物种随着时间的推移而发生变化时,只要种群存活下来或生物灭绝,它就会反复分支形成多个新物种。

要实现物种形成,必须从一个原始种群中形成两个新种群,并且它们的进化方式必须使来自两个新种群的个体无法杂交。 生物学家提出了可能发生这种情况的机制,分为两大类。 All@@ opatric 物种形成(allo-= “其他”;-patric = “家园”)涉及种群与亲本物种的地理分离以及随后的进化。 Sy@@ mpatric 物种形成(sym-= “相同”;-patric = “家园”)涉及留在一个地点的亲本物种内部发生的物种形成。
生物学家认为物种形成事件是将一个祖先物种分裂成两个后代物种。 没有理由为什么一次形成的物种不超过两个,只是这种可能性较小,而且可以将多个事件概念化为单次分裂在近时间内发生。
同种异体形态形成
地理上连续的人群拥有相对同质的基因库。 基因流,即等位基因在物种范围内的移动,相对自由,因为个体可以移动,然后在新的位置与个体交配。 因此,分布一端的等位基因的频率将与另一端等位基因的频率相似。 当种群在地理上变得不连续时,等位基因的自由流动就会受到阻碍。 当这种分离持续一段时间时,这两个种群就能够沿着不同的轨迹进化。 因此,随着每个群体中的突变独立产生新的等位基因,它们在众多遗传基因座上的等位基因频率逐渐变得越来越不同。 通常,两个种群的气候、资源、捕食者和竞争对手等环境条件会有所不同,导致自然选择倾向于每个群体的不同适应方式。
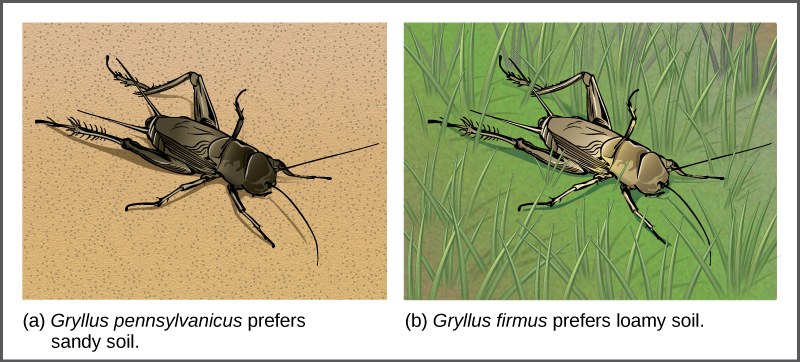
分离导致同种异体物种形成的种群可以通过多种方式发生:河流形成新的分支,侵蚀形成新的山谷,一群无法返回的新地点的生物,或者种子漂浮在海洋上空到岛屿。 隔离种群所必需的地理分离的性质完全取决于生物体的生物学及其传播潜力。 如果两个飞行昆虫种群居住在附近的不同山谷中,则每个种群中的个体很可能会来回飞行,继续进行基因流动。 但是,如果两个啮齿动物种群因新湖的形成而分开,则不太可能持续进行基因流动;因此,物种形成的可能性更大。
生物学家将同种异体过程分为两类:传播和替代过程。 传播是指物种的少数成员迁移到新的地理区域,而替代是指出现物理分裂生物的自然情况。
科学家记录了许多同种异体物种形成的案例。 例如,在美国西海岸,存在两个单独的斑点猫头鹰亚种。 北方斑点猫头鹰与其近亲有遗传和表型差异:生活在南方的墨西哥斑点猫头鹰(图\(\PageIndex{4}\))。
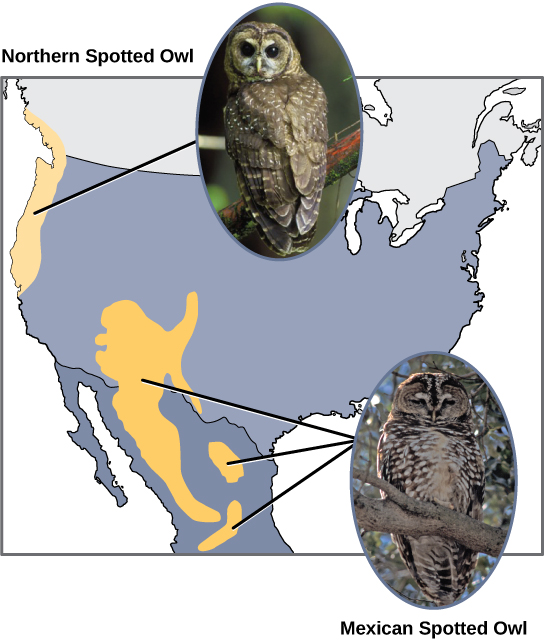
此外,科学家们发现,曾经是同一个物种的两个群体之间的距离越远,物种形成的可能性就越大。 这似乎是合乎逻辑的,因为随着距离的增加,各种环境因素的共同点可能不如近距离的地点那么共同点。 以两只猫头鹰为例:在北方,气候比南方凉爽;每个生态系统中的生物类型不同,它们的行为和习惯也不同;此外,南方猫头鹰的狩猎习惯和猎物选择与北方猫头鹰不同。 这些差异可能导致猫头鹰的进化差异,并且很可能会发生物种形成。
自适应辐射
在某些情况下,一个物种的种群分散在整个区域,每个物种都会找到不同的利基或孤立的栖息地。 随着时间的推移,他们新生活方式的不同需求导致了源于单一物种的多种物种形成事件。 这被称为自适应辐射,因为许多适应是从单一起源点演变而来的;因此,导致物种辐射成几个新的物种。 像夏威夷群岛这样的岛屿群岛为适应性辐射事件提供了理想的环境,因为水环绕着每个岛屿,这导致许多生物在地理上处于隔离状态。 夏威夷蜜爬虫举例说明了自适应辐射的一个例子。 从一个称为创始物种的单一物种中进化出许多物种,包括图中所示的六个物种\(\PageIndex{5}\)。
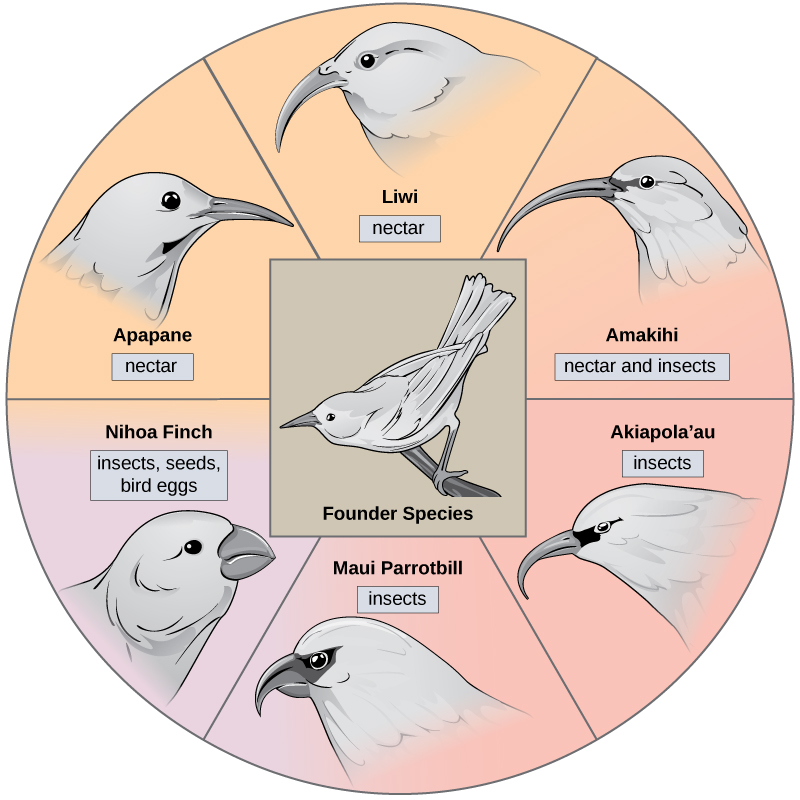
注意图中物种的喙的差异\(\PageIndex{5}\)。 在每个新栖息地中,基于特定食物来源的自然选择而进化,导致了适合特定食物来源的不同喙的进化。 吃种子的鸟的喙更厚、更强壮,适合打破坚硬的坚果。 吃花蜜的鸟有长嘴可以浸入花中才能到达花蜜。 吃昆虫的鸟有像剑一样的嘴,适合刺伤和刺穿昆虫。 达尔文的雀科是群岛中适应性辐射的又一个例子。
链接到学习
点击这个互动网站,查看从500万年前到今天,岛鸟是如何以进化增量进化的。
Sympatric 形态形成
如果没有物理屏障来隔离继续在同一栖息地生活和繁殖的个体,会发生分歧吗? 答案是肯定的。 同一个空间内的物种形成过程称为同情物种形成;前缀 “sym” 的含义相同,因此 “sympatric” 的意思是 “同一个家园”,而 “allopatric” 的意思是 “其他家园”。 已经提出并研究了许多同情物种形成的机制。
一种形式的同情物种形成可能始于细胞分裂过程中的严重染色体错误。 在正常的细胞分裂事件中,染色体复制、配对,然后分离,这样每个新细胞都有相同数量的染色体。 但是,有时两对会分开,终末细胞产物的单个染色体过多或过少,这种情况称为非整倍体(图\(\PageIndex{6}\))。
艺术连接
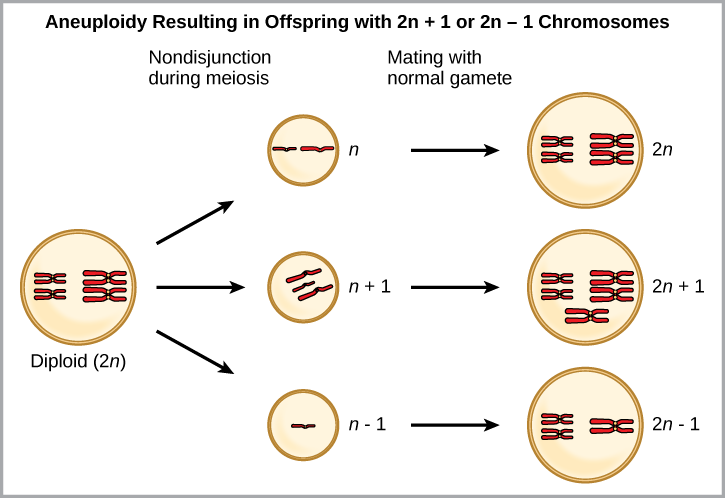
哪个最有可能存活,是有 2 n +1 染色体的后代还是拥有 2 n -1 染色体的后代?
多倍体是一种细胞或生物体拥有一组或多组额外染色体的情况。 科学家们已经确定了两种主要的多倍体类型,它们可能导致处于多倍体状态的个体的生殖隔离。 生殖隔离是指无法杂交。 在某些情况下,多倍体个体将拥有来自自身物种的两组或更多完整的染色体,这种情况称为自多倍体(图\(\PageIndex{7}\))。 前缀 “auto-” 表示 “自我”,因此该术语表示来自自己物种的多条染色体。 多倍体是由减数分裂中的一个错误造成的,在该错误中,所有染色体都移入一个细胞,而不是分离。
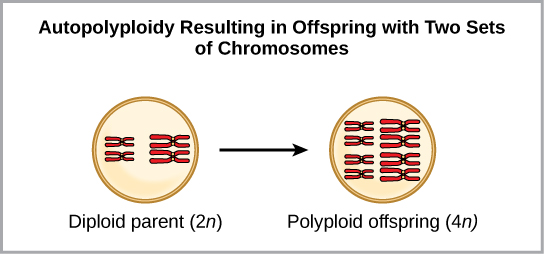
例如,如果 2 n = 6 的植物物种产生的自多倍体配子也是二倍体(2 n = 6,当它们应该为 n = 3 时),则配子现在的染色体数量是应有的两倍。 这些新配子将与该植物物种产生的普通配子不相容。 但是,它们既可以自授粉,也可以与其他配子具有相同二倍体数的自多倍体植物一起繁殖。 这样,通过形成 4 n 的后代,称为四倍体,可以快速形成同情物种形态。 这些个体只能用这种新物种而不是祖先物种的个体立即繁殖。
另一种形式的多倍体发生在两个不同物种的个体繁殖形成称为同种异体多倍体的可存活后代时。 前缀 “allo-” 的意思是 “其他”(从同种异体中召回):因此,当来自两个不同物种的配子结合时,就会出现同种异体多倍体。 该图\(\PageIndex{8}\)说明了同种多倍体形成的一种可能方式。 请注意,需要两代人或两代人的繁殖行为才能产生可行的肥沃杂种。
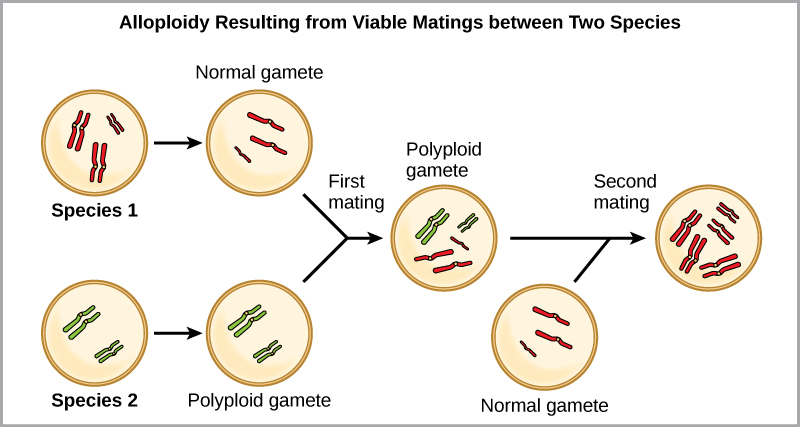
小麦、棉花和烟草植物的栽培形式都是同种异体类。 尽管多倍体偶尔发生在动物身上,但它最常发生在植物中。 (具有此处描述的任何类型的染色体畸变的动物都不太可能存活并产生正常的后代。) 科学家们发现,在所有研究的植物物种中,有一半以上与通过多倍体进化的物种有关。 由于植物中的多倍体率如此之高,一些科学家假设这种机制更多地是一种适应而不是错误。
生殖隔离
如果有足够的时间,种群之间的遗传和表型差异将影响繁殖的特征:如果将两个种群中的个体聚集在一起,交配的可能性就会降低,但如果发生交配,后代将无法存活或不育。 许多类型的不同角色可能会影响两个种群的生殖隔离,即杂交能力。
生殖隔离可以通过多种方式进行。 科学家将它们分为两组:prezygotic 屏障和合子后屏障。 回想一下,合子是受精卵:生物体发育的第一个性繁殖细胞。 因此,prezygotic 屏障是一种阻断繁殖的机制;这包括在生物尝试繁殖时阻止受精的屏障。 合子形成后会出现合子后屏障;这包括无法在胚胎阶段存活的生物和出生时不育的生物。
某些类型的 prezygotic 屏障完全阻碍了繁殖。 许多生物只在一年中的特定时间繁殖,通常每年才繁殖。 育种时间表的差异,称为时间隔离,可以作为生殖隔离的一种形式。 例如,有两种青蛙栖息在同一区域,但一种在1月至3月繁殖,而另一种在三月至五月繁殖(图\(\PageIndex{9}\))。

在某些情况下,一个物种的种群会移动或被转移到新的栖息地,并在不再与同一物种的其他种群重叠的地方居住。 这种情况称为栖息地隔离。 与亲本物种的繁殖停止,出现了一个新的群体,该群体现在在繁殖和遗传上是独立的。 例如,洪水后分裂的板球种群无法再相互作用。 随着时间的推移,自然选择、突变和遗传漂移的力量可能会导致两组的分歧(图\(\PageIndex{10}\))。
当特定行为的存在或不存在阻碍繁殖时,就会发生行为@@ 隔离。 例如,雄性萤火虫使用特定的光线模式来吸引雌性萤火虫。 不同种类的萤火虫显示灯光的方式不同。 如果一个物种的雄性试图吸引另一个物种的雌性,她将无法识别光线模式,也不会与雄性交配。
当配子细胞(卵子和精子)的差异阻止受精时,其他 prezygot ic 屏障就会起作用;这被称为配子屏障。 同样,在某些情况下,密切相关的生物会尝试交配,但它们的繁殖结构根本无法融合在一起。 例如,不同物种的豆娘雄性的生殖器官形状不同。 如果一个物种试图与另一个物种的雌性交配,它们的身体部位根本无法组合在一起(图\(\PageIndex{11}\))。

在植物中,某些旨在同时吸引一种传粉媒介的结构会阻止另一种传粉媒介进入花粉。 动物必须经过的隧道在长度和直径上差异很大,这可以防止植物与其他物种进行异花授粉(图\(\PageIndex{12}\))。
当受精发生并形成合子时,合子后屏障可以阻止繁殖。 在许多情况下,杂交个体无法在子宫内正常形成,根本无法存活到胚胎阶段。 这被称为杂交隐身,因为杂交生物根本无法生存。 在另一种合子后情况中,繁殖会导致不育且无法繁殖自己的后代的杂种的诞生和生长;这就是所谓的杂交不育。
栖息地对物种形成的影响
Sympatric 物种形成也可能以多倍体以外的其他方式发生。 例如,以生活在湖中的鱼类为例。 随着人口的增长,对食物的竞争也在加剧。 在寻找食物的压力下,假设其中一组鱼具有遗传灵活性,可以发现并以其他鱼类未使用的另一种资源为食。 如果在湖的不同深度发现了这种新的食物来源会怎样? 随着时间的推移,以第二种食物来源为食的人之间的相互作用比其他鱼类的互动更多;因此,它们也会一起繁殖。 这些鱼的后代很可能会像他们的父母一样行事:在同一个区域喂食和生活,与原始种群分开。 如果这组鱼继续与第一个种群分开,那么随着它们之间积累更多的遗传差异,最终可能会出现同情形态的形成。
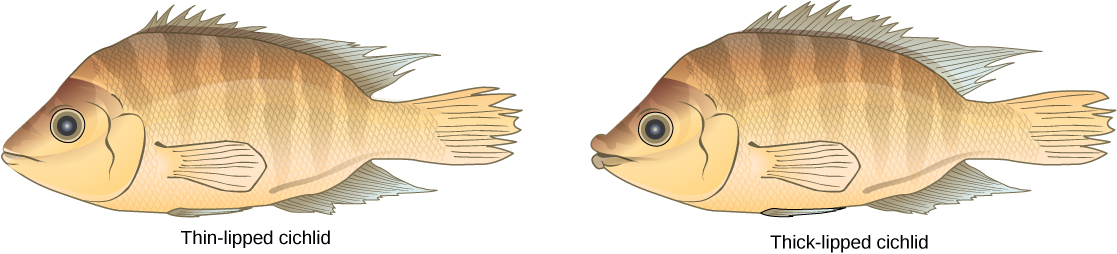
这种情况确实在自然界中发挥作用,其他导致生殖隔离的情况也是如此。 其中一个地方就是非洲的维多利亚湖,它以慈鲷鱼的同情形态而闻名。 研究人员在这些鱼类中发现了数百个同情物种形成事件,这些事件不仅大量发生,而且发生在很短的时间内。 该图\(\PageIndex{13}\)显示了尼加拉瓜慈鲷鱼种群中的这种物种形态。 在这个地方,两种类型的慈鲷生活在相同的地理位置,但它们的形态却不同,使它们能够食用不同的食物来源。
摘要
物种形成沿两个主要途径发生:地理分离(同种异体物种形成)和通过共享栖息地内发生的机制(同情物种形成)。 这两种途径都以某种形式进行生殖隔离。 生殖隔离机制是密切相关的物种之间的屏障,使它们能够分化并作为遗传独立物种存在。 Prezygotic 屏障在合子形成之前会阻断繁殖,而合子后屏障在受精发生后会阻断繁殖。 要培育出一个新物种,一定有某种东西会导致生殖屏障的突破。 Sympatric 物种形成可能是由于减数分裂中的错误而发生的,减数分裂会形成带有额外染色体(多倍体)的配子。 Autopolyploidy 发生在单个物种中,而同种多倍体发生在密切相关的物种之间。
艺术联系
图\(\PageIndex{6}\):哪个最有可能存活,后代有 2 n +1 条染色体,还是后代有 2 n -1 条染色体?
- 回答
-
遗传物质的流失几乎总是致命的,因此拥有 2 n +1 条染色体的后代更有可能存活。
词汇表
- 自适应辐射
- 当一个物种辐射出来形成其他几个物种时的物种形成
- 同种异体形态形成
- 通过地理分离产生的形态形成
- 同种异体多倍体
- 在两个相关但独立的物种之间形成多倍体
- 非整倍体
- 细胞有多余的染色体或缺少其物种的染色体的状况
- 自多倍体
- 在单个物种中形成多倍体
- 行为隔离
- 一种生殖隔离,发生在特定的行为或缺乏行为阻碍生殖发生时
- 扩散的
- 同种异体物种形成发生在物种的少数成员迁移到新的地理区域时
- 游戏屏障
- prezygotic barrier 发生在不同物种密切相关的个体交配时,但他们的配子细胞(卵子和精子)的差异阻碍了受精的发生
- 栖息地隔离
- 当一个物种的种群移动或迁移到新的栖息地,居住在不再与同一物种的其他种群重叠的地方时,就会导致生殖隔离
- 混合动力
- 两个密切相关的个体的后代,不是同一个物种
- 合子后屏障
- 合子形成后发生的生殖隔离机制
- prezygotic 屏障
- 合子形成之前发生的生殖隔离机制
- 生殖隔离
- 当一个物种在生殖上独立于其他物种时发生的情况;这可能是由行为、位置或繁殖障碍造成的
- 形态
- 新物种的形成
- 物种
- 一群杂交并产生可育后代的种群
- 同情形态形成
- 出现在同一个地理空间中的物种形态
- 时间隔离
- 育种时间表的差异可能成为一种预合屏障,导致生殖隔离
- 代教
- 当环境中的某物将相同物种的生物分成不同的组时,就会发生同种异体物种形态的形成