
18.1: 了解进化
- Page ID
- 202166

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 描述当今的进化论是如何发展的
- 定义适应
- 解释趋同和发散的演变
- 描述同源结构和残余结构
- 讨论对进化论的误解
自然选择的进化描述了物种如何随着时间的推移而变化的机制。 早在达尔文开始探索这个想法之前,物种变化就已经提出过建议和辩论。 认为物种是静止不变的观点以柏拉图的著作为基础,但也有古希腊人表达了进化思想。 在十八世纪,博物学家乔治·路易·勒克莱尔·孔德·布冯重新引入了关于动物进化的观念,他观察到即使环境相似,各个地理区域也有不同的动植物种群。 人们还承认存在灭绝的物种。
在此期间,苏格兰博物学家詹姆斯·赫顿(James Hutton)提出,地质变化是通过长期以来像今天一样运作的过程产生的微小变化积累而逐渐发生的。 这与占主导地位的观点形成鲜明对比,即地球的地质学是相对较短的过去发生的灾难性事件的结果。 赫顿的观点在十九世纪被成为达尔文朋友的地质学家查尔斯·莱尔所普及。 莱尔的想法对达尔文的思维产生了影响:莱尔关于更长的地球时代的概念为物种的逐步变化提供了更多的时间,而变化的过程为物种的逐渐变化提供了一个类比。 十九世纪初,让-巴蒂斯特·拉马克出版了一本书,详细介绍了进化变化的机制。 这种机制现在被称为后天特征的继承,通过这种遗传,个体因其环境而发生变化,或者在其生命周期内使用或废弃结构,可以由其后代继承,从而带来物种的变化。 尽管这种进化变革机制声名狼藉,但拉马克的思想对进化思想产生了重要影响。
查尔斯·达尔文与自然选择
十九世纪中叶,进化的实际机制是由两位博物学家独立构思和描述的:查尔斯·达尔文和阿尔弗雷德·罗素·华莱士。 重要的是,每位博物学家都花时间在热带探险中探索自然世界。 从 1831 年到 1836 年,达尔文乘坐 H.M.S. Beagle 环游世界,包括在南美、澳大利亚和非洲南端停留。 华莱士于 1848 年至 1852 年前往巴西在亚马逊雨林采集昆虫,1854 年至 1862 年前往马来群岛。 达尔文的旅程,就像华莱士后来前往马来群岛的旅程一样,包括在几个岛屿链上停留,最后一次是厄瓜多尔以西的加拉帕戈斯群岛。 在这些岛屿上,达尔文观察到不同岛屿上存在明显相似但有明显差异的生物种类。 例如,居住在加拉帕戈斯群岛的地下雀由几种具有独特喙形状的物种组成(图\(\PageIndex{1}\))。 岛上的物种有一系列分级的喙大小和形状,最相似的鸟之间的差异很小。 他观察到这些雀科与南美大陆的另一种雀科物种非常相似。 达尔文认为岛屿物种可能是从原始大陆物种之一改造而来的物种。 经过进一步研究,他意识到每只雀的喙各不相同,可以帮助鸟类获得特定类型的食物。 例如,吃种子的雀有更强、更粗的喙可以破碎种子,而吃昆虫的雀有长矛状的喙用来刺伤猎物。
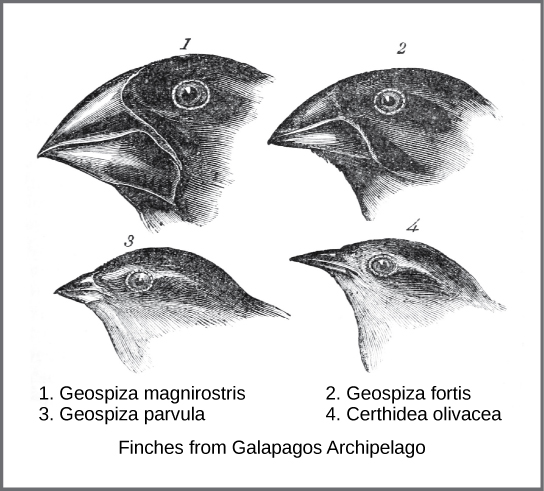
华莱士和达尔文在其他生物体中都观察到了相似的模式,他们对如何以及为什么会发生这种变化独立地做出了相同的解释。 达尔文称这种机制为自然选择。 自然选择,也被称为 “适者生存”,是指具有良好特征的个体因这些特征而在环境变化中幸存下来的更高产繁殖;这会导致进化变化。
例如,达尔文观察到在加拉帕戈斯群岛发现的巨型乌龟的脖子比生活在其他低地干燥岛屿上的乌龟脖子长。 这些乌龟之所以被 “选中”,是因为与脖子短的乌龟相比,它们可以获得更多的树叶和更多的食物。 在干旱时期,可用的叶子会减少,那些能长出更多叶子的叶子比那些无法获得食物来源的叶子有更好的进食和存活机会。 因此,长颈乌龟更有可能在繁殖方面取得成功,并将长颈特质传给它们的后代。 随着时间的推移,种群中只会出现长颈乌龟。
达尔文认为,自然选择是自然界中运作的三项原则的必然结果。 首先,生物的大多数特征都是遗传的,或者是从父母传给后代的。 尽管当时没有人,包括达尔文和华莱士,知道这是怎么发生的,但这是一个普遍的认识。 其次,产生的后代多于存活的能力,因此存活和繁殖的资源有限。 所有生物的繁殖能力都超过了支持其数量的可用资源。 因此,每一代人都在争夺这些资源。 达尔文和华莱士对这一原则的理解都来自阅读经济学家托马斯·马尔萨斯的一篇文章,他讨论了这一原则与人口的关系。 第三,后代的特征各不相同,这些变异是遗传的。 达尔文和华莱士认为,具有遗传特征、使他们能够最好地竞争有限资源的后代将比那些具有竞争能力较差的变体的人存活下来并拥有更多的后代。 由于特征是遗传的,因此这些特征将在下一代中得到更好的体现。 这将导致人口在几代人之间发生变化,达尔文称之为经过修改的血统。 最终,自然选择会使人口更好地适应当地环境;它是唯一已知的适应性进化机制。
1858 年,在伦敦林纳协会之前,一起阅读了达尔文和华莱士(图\(\PageIndex{2}\))介绍自然选择概念的论文。 次年,达尔文出版了《论物种起源》一书。 他的书相当详细地概述了他关于自然选择进化的论点。

通过自然选择演示进化既费时又难以获得。 最好的例子之一就是帮助启发达尔文理论的鸟:加拉帕戈斯雀科。 自1976年以来,彼得和罗斯玛丽·格兰特及其同事每年都对加拉帕戈斯雀科种群进行研究,并提供了自然选择的重要示范。 Grants 发现,加拉帕戈斯岛达芙妮·梅杰岛上有中等地面雀的喙形状的分布从一代到下一代发生了变化。 这些鸟继承了账单形状的变化,有些鸟的账单很大,而另一些鸟的账单更薄。 在厄尔尼诺现象导致降雨量高于正常水平的时期,大嘴鸟吃的大型硬种子数量减少了;但是,小嘴鸟吃了大量的小软种子。 因此,在接下来的几年中,小嘴鸟的存活和繁殖要好得多。 在厄尔尼诺现象之后的几年中,补助金测量了种群中的喙大小,发现平均账单规模较小。 由于账单大小是一种遗传特征,因此账单较小的父母有更多的后代,账单的规模也变小了。 随着1987年条件的改善和更大的种子越来越多,平均账单规模缩小的趋势停止了。
职业联系:野外生物学家
许多人徒步旅行、探索洞穴、水肺潜水或爬山进行娱乐。 人们经常参加这些活动,希望看到野生动物。 体验户外活动可以令人难以置信的愉快和充满活力。 如果你的工作是待在野外呢? 顾名思义,野外生物学家在 “野外” 工作。 在本例中,“字段” 一词是指室外的任何位置,甚至是水下。 野外生物学家通常将研究重点放在特定物种、生物群或单一栖息地上(图\(\PageIndex{3}\))。

许多野外生物学家的目标之一是发现从未记录过的新物种。 这些发现不仅扩大了我们对自然界的理解,而且还带来了医学和农业等领域的重要创新。 特别是植物和微生物物种,可以揭示新的药物和营养知识。 其他生物可以在生态系统中发挥关键作用,或者被认为是稀有的,需要保护。 一旦被发现,这些重要物种可以用作环境法规和法律的证据。
进化的过程和模式
只有在人群中的个体之间存在变异或差异时,才能进行自然选择。 重要的是,这些差异必须有一定的遗传基础;否则,这种选择不会导致下一代的改变。 这很关键,因为个体之间的变异可能是由非遗传原因引起的,例如个体因为营养好而不是基因不同而变高。
人群的遗传多样性来自两个主要机制:突变和有性生殖。 突变是DNA的改变,是任何群体中新等位基因或新遗传变异的最终来源。 突变引起的遗传变化可能是表型的三种结果之一。 突变会影响生物体的表型,使其健康状况降低,存活的可能性降低或后代减少。 突变可能会产生对健康有益影响的表型。 而且,许多突变也不会影响表型的适应性;这些突变被称为中性突变。 突变还可能对以其表型表达它们的生物的适应性产生各种效应大小,从小效应到大效应不等。 有性生殖还会导致遗传多样性:当两个父母繁殖时,独特的等位基因组合会聚在一起,产生独特的基因型,从而在每个后代中产生表型。
一种有助于生物在当前环境中存活和繁殖的可遗传特征称为适应。 科学家描述了当遗传变异范围随着时间的推移发生变化,从而增加或维持种群与环境的 “适应性” 时,生物群体就会适应其环境。 鸭嘴兽的网状脚适合游泳。 雪豹厚实的皮毛适合在寒冷中生活。 猎豹的速度很快,适合捕捉猎物。
某一特质是否有利取决于当时的环境条件。 并非总是选择相同的特征,因为环境条件可能会发生变化。 例如,假设一种植物在潮湿的气候中生长,不需要节约用水。 之所以选择大叶子,是因为它们使植物能够从太阳中获得更多能量。 大叶子比小叶子需要更多的水来维持,潮湿的环境为支撑大叶提供了有利的条件。 几千年后,气候发生了变化,该地区不再有过多的水。 自然选择的方向发生了变化,因此选择了叶子较小的植物,因为这些种群能够节约用水以在新的环境条件下生存。
物种的进化导致了形态和功能的巨大差异。 有时,进化会产生彼此截然不同的生物群。 当两个物种从一个共同点朝不同的方向进化时,它被称为发散进化。 这种不同的进化可以从具有相同基本解剖结构的开花植物的生殖器官的形式中看出;但是,由于在不同的物理环境中进行选择以及对不同种类的传粉媒介的适应,它们的外观可能会大不相同(图\(\PageIndex{4}\))。

在其他情况下,相似的表型在远距离相关的物种中独立进化。 例如,蝙蝠和昆虫都有飞行进化,它们都有我们称之为翅膀的结构,可以适应飞行。 但是,蝙蝠和昆虫的翅膀是从截然不同的原始结构进化而来的。 这种现象被称为融合进化,即相似的特征在最近没有共同血统的物种中独立进化。 这两个物种起到了相同的作用,飞行,但彼此分开飞行。
这些物理变化发生在很长的一段时间内,有助于解释进化是如何发生的。 自然选择作用于单个生物,而个体生物反过来又可以塑造整个物种。 尽管自然选择可能在一代人之内对个体起作用,但整个物种的基因型可能需要数千甚至数百万年的时间才能进化。 正是在这么长的时间跨度内,地球上的生命发生了变化并且还在继续变化。
进化的证据
进化的证据令人信服且广泛。 从生命系统的各个组织层面来看,生物学家看到了过去和现在进化的标志。 达尔文在他的《论物种起源》一书中用了很大一部分时间来识别自然界中与进化相一致的模式,自达尔文以来,我们的理解变得更加清晰和广泛。
化石
化石提供了确凿的证据,表明过去的生物与今天发现的生物不同,化石显示出进化的进展。 科学家确定化石的年龄,并将其从世界各地进行分类,以确定这些生物何时相对于彼此生活。 由此产生的化石记录讲述了过去的故事,并显示了数百万年来形态的演变(图\(\PageIndex{5}\))。 例如,科学家已经恢复了显示人类和马匹进化的非常详细的记录(图\(\PageIndex{5}\))。 鲸鳍的形态与鸟类和哺乳动物的附属物类似(图\(\PageIndex{6}\)),表明这些物种有着共同的祖先。

解剖学和胚胎学
进化的另一种证据是生物体中存在具有相同基本形态的结构。 例如,人类、狗、鸟和鲸的附属物中的骨头都具有相同的整体结构(图\(\PageIndex{6}\)),因为它们起源于共同祖先的附属物。 随着时间的推移,进化导致不同物种中这些骨骼的形状和大小发生变化,但它们的总体布局保持不变。 科学家称这些同义部分为同源结构。
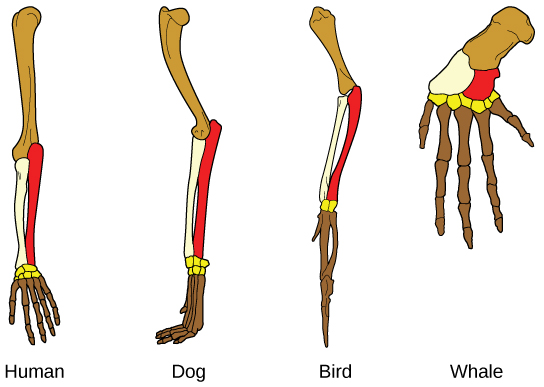
有些结构存在于完全没有明显功能的生物体中,似乎是过去共同祖先的残留部分。 这些没有功能的未使用结构称为残余结构。 其他残留结构的例子包括不会飞的鸟的翅膀、一些仙人掌上的树叶和鲸鱼的后腿骨。
链接到学习
访问这个互动网站,猜测哪些骨骼结构是同源的,哪些是相似的,并查看进化适应的例子来说明这些概念。
进化的另一个证据是共享相似环境的生物的形态趋同。 例如,生活在北极地区的无关动物物种,例如北极狐和雷鸟,被选为冬季的季节性白色表型,以便与冰雪融为一体(图\(\PageIndex{7}\))。 这些相似之处不是因为共同的血统,而是因为相似的选择压力——不被捕食者看见的好处。

胚胎学是研究生物体解剖结构发展到成体形态的研究,它也为现在差异很大的生物群体之间存在关联性提供了证据。 胚胎的突变调整可能会对成年人产生如此严重的后果,以至于胚胎的形成往往是保守的。 因此,某些群体中不存在的结构通常以其胚胎形式出现,并在达到成体或幼体形态时消失。 例如,所有脊椎动物胚胎,包括人类,在早期发育的某个阶段都表现出鳃缝和尾巴。 它们在陆生群体的成年人中消失,但在鱼类和一些两栖动物等成虫形式的水生群体中得以维持。 大猿胚胎,包括人类,在发育过程中具有尾巴结构,在出生时会消失。
生物地理学
地球上生物的地理分布遵循的模式最能通过演化以及地质时期构造板块的运动来解释。 在超大陆Pangea解体(大约2亿年前)解体之前进化的广泛群体分布在世界各地。 自解体以来进化的群体在地球各地区表现得独一无二,例如由超大陆劳拉西亚形成的北方大陆的独特动植物群和由冈瓦纳超大陆形成的南方大陆的独特动植物。 Proteaceae 植物家族成员在澳大利亚、南部非洲和南美的存在,最好用它们在南部超大陆冈瓦纳解体之前的存在来解释。
澳大利亚有袋动物的巨大多样化以及其他哺乳动物的缺乏反映了澳大利亚的长期隔离。 澳大利亚有大量的特有物种——其他任何地方都找不到的物种——这是岛屿的典型特征,这些岛屿被大片水域隔离,使物种无法迁移。 随着时间的推移,这些物种在进化上分化为新的物种,这些物种看起来与可能存在于大陆的祖先截然不同。 澳大利亚的有袋动物、加拉帕戈斯群岛上的雀科以及夏威夷群岛上的许多物种都因其单一起源点而独一无二,但它们与大陆的祖先物种表现出遥远的关系。
分子生物学
与解剖结构一样,生命分子的结构通过修改反映了血统。 DNA作为遗传物质的普遍性以及遗传密码和DNA复制和表达机制的近乎普遍性反映了所有生命都有共同祖先的证据。 这三个领域之间生命的基本分歧反映在原本保守的结构(例如核糖体成分和膜结构)的重大结构差异上。 总的来说,生物群体的关联性反映在它们的 DNA 序列的相似性上,这正是共同祖先的血统和多样化所期望的模式。
DNA序列也揭示了一些进化机制。 例如,很明显,蛋白质新功能的演变通常发生在基因复制事件之后,这些事件允许通过突变、选择或漂移(偶然导致人群基因库的变化)自由修改一个拷贝,而第二个拷贝继续产生功能性的蛋白质。
对进化的误解
尽管进化论在最初提出时引起了一些争议,但在《论物种起源》出版后的20年内,它几乎被生物学家,尤其是年轻的生物学家普遍接受。 尽管如此,进化论是一个困难的概念,对它如何运作的误解比比皆是
链接到学习
该网站解决了与进化论有关的一些主要误解。
进化只是一种理论
对进化论的批评者故意将 “理论” 一词的日常用法与科学家使用该词的方式混为一谈,从而否认了其重要性。 在科学中,“理论” 被理解为对自然世界的一系列观测结果进行全面测试和验证的解释。 科学家们有原子理论、引力理论和相对论,每种理论都描述了关于世界的可理解事实。 同样,进化论描述了有关生活世界的事实。 因此,科学理论在科学家为抹黑其信誉而付出的巨大努力中幸存下来。 相比之下,普通白话中的 “理论” 是一个意思是猜测或建议解释的词;这个含义更类似于 “假设” 的科学概念。 当进化论的批评者说进化论 “只是一个理论” 时,他们暗示几乎没有证据支持进化,它仍在接受严格的考验。 这是一种错误的描述。
个人进化
进化是指具有某些等位基因的个体差异繁殖导致的种群遗传组成随着时间的推移,特别是世代相传的变化。 显然,个体在一生中确实会发生变化,但这被称为发育,它涉及由个人在出生时获得的与个体环境协调后获得的基因组编程的变化。 在考虑特征的演变时,最好考虑总体中该特征的平均值随着时间的推移而发生的变化。 例如,当自然选择导致加拉帕戈斯群岛中地雀的账单大小变化时,这并不意味着雀科的单个账单正在发生变化。 如果一次测量人口中所有个人的平均账单规模,然后在几年后测量人口的平均账单规模,那么这个平均值将随着演变而有所不同。 尽管有些人可以从第一次存活到第二次存活,但他们的账单金额仍将保持不变;但是,会有许多新人为平均账单规模的变化做出贡献。
进化解释了生命的起源
人们普遍误解进化包括对生命起源的解释。 相反,该理论的一些批评者认为它无法解释生命的起源。 该理论并不试图解释生命的起源。 进化论解释了种群如何随着时间的推移而变化,以及生命如何使物种起源多样化。 它没有揭示生命的起源,包括第一个细胞的起源,这就是生命的定义。 地球生命起源的机制是一个特别困难的问题,因为它发生在很久以前,而且可能只发生过一次。 重要的是,生物学家认为,地球上生命的存在排除了导致地球生命的事件重演的可能性,因为中间阶段将立即成为现有生物的食物。
但是,一旦在细胞内或细胞前以像DNA这样的分子形式建立了遗传机制,这些实体将受自然选择原则的约束。 更有效的繁殖者会增加频率,而牺牲了效率低下的繁殖者。 因此,尽管进化无法解释生命的起源,但对于活前实体获得某些属性后运行的某些过程,它可能有话要说。
生物是故意进化的
诸如 “生物因环境变化而进化” 之类的说法很常见,但这样的说法可能导致两种类型的误解。 首先,不能将该陈述理解为意味着个体生物在进化。 这句话是 “人口为应对不断变化的环境而演变” 的简写。 但是,如果将该陈述解释为意味着演变在某种程度上是故意的,则可能会产生第二个误解。 环境的变化会导致人群中的一些个体,即具有特定表型的人,受益于其他表型,因此产生的后代成比例地比其他表型多。 如果特征由基因决定,则会导致种群发生变化。
同样重要的是要明白,自然选择起作用的变异已经存在于人群中,不是因环境变化而产生的。 例如,随着时间的推移,对一群细菌使用抗生素将选择对抗生素具有耐药性的细菌群体。 耐药性是由基因引起的,不是因为抗生素的应用而由突变引起的。 抗药基因已经存在于细菌的基因库中,频率可能很低。 这种抗生素可以杀死没有耐药基因的细菌细胞,它强烈选择具有耐药性的个体,因为这些人将是唯一存活和分裂的个体。 实验表明,抗生素耐药性突变不是由抗生素引起的。
从更大的意义上讲,进化不是以目标为导向的。 物种不会随着时间的推移变得 “更好”;它们只是通过适应来跟踪不断变化的环境,从而在特定时间最大限度地提高其在特定环境中的繁殖。 尽管这种语言在流行话语中很常见,但进化的目标不是创造更快、更大、更复杂甚至更聪明的物种。 物种中哪些特征的演变是当前变异和环境的函数,两者都在以非方向性的方式不断变化。 在将来的某个时候,哪种特质适合一个环境很可能是致命的。 这对昆虫和人类物种同样适用。
摘要
进化是通过突变进行适应的过程,它允许将更理想的特征传给下一代。 随着时间的推移,生物会进化出更多有利于其生存的特征。 为了使活生物体适应和改变环境压力,必须存在遗传变异。 随着遗传变异,个体在形式和功能上存在差异,这使一些人能够比其他人更好地存活某些条件。 这些生物将其有利的特征传给了后代。 最终,环境会发生变化,曾经理想的有利特征可能会变成不良特征,生物可能会进一步进化。 进化可能是趋同的,相似的特征在多个物种中进化,或者在来自共同祖先的多个物种中进化出不同的特征。 进化证据可以通过DNA密码和化石记录来观察,也可以通过同源和残留结构的存在来观察。
词汇表
- 适应
- 生物体中有助于其在当前环境中存活和繁殖的可遗传特征或行为
- 趋同进化
- 生物群独立进化为相似形态的过程
- 发散进化
- 生物群从共同点向不同方向进化的过程
- 同源结构
- 具有共同祖先的不同生物中的平行结构
- 自然选择
- 具有良好遗传特征的个体的繁殖,这些个体由于这些特征而在环境变化中幸存下来,从而导致进化变化
- 变体
- 人群中个体之间的遗传差异
- 残余结构
- 物理结构存在于生物体中,但没有明显的功能,似乎来自遥远祖先的功能结构