
16.5: 真核生物转录后基因调控
- Page ID
- 202984

培养技能
- 了解 RNA 拼接并解释其在调节基因表达中的作用
- 描述 RNA 稳定性在基因调控中的重要性
RNA 是经过转录的,但在开始翻译之前必须经过处理成熟的形式。 这种在 RNA 分子被转录之后但在转化为蛋白质之前的处理称为转录后修饰。 与加工的表观遗传学和转录阶段一样,也可以调节这个转录后步骤以控制细胞中的基因表达。 如果不对RNA进行处理、穿梭或翻译,则不会合成任何蛋白质。
RNA 剪接,转录后控制的第一阶段
在真核细胞中,RNA转录本通常包含称为内含子的区域,这些区域在翻译之前被移除。 编码蛋白质的 RNA 区域称为外显子(图\(\PageIndex{1}\))。 在 RNA 分子被转录后,但在它离开待翻译的细胞核之前,会对 RNA 进行处理并通过拼接去除内含子。
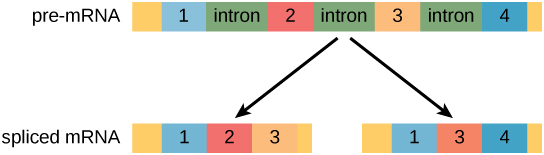
进化连接:替代 RNA 拼接
在20世纪70年代,首次观察到表现出替代性RNA拼接的基因。 替代性RNA拼接是一种机制,当从转录本中移除不同的内含子(有时是外显子)组合时,允许从一个基因中产生不同的蛋白质产物(图\(\PageIndex{2}\))。 这种替代拼接可能是随意的,但更常见的是它受到控制并起到基因调控机制的作用,不同的拼接替代方案的频率由细胞控制,以此来控制不同细胞中或不同阶段不同蛋白质产物的产生开发。 现在,人们认为替代拼接是真核生物中常见的基因调控机制;根据一项估计,人类70%的基因是通过替代拼接以多种蛋白质表达的。
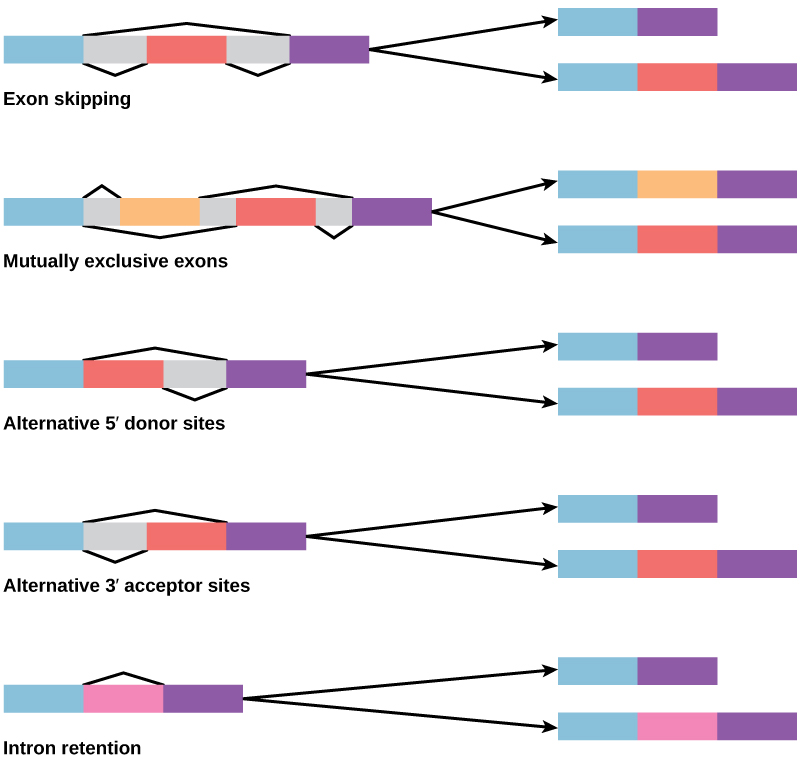
替代拼接会如何演变? 内含子有开头和结尾的识别序列;很容易想象拼接机制无法识别内含子的末端,而是找到下一个内含子的结尾,从而移除两个内含子和中间的外显子。 实际上,有防止这种内含子跳过的机制,但是突变很可能导致其失效。 这种 “错误” 很可能会产生不起作用的蛋白质。 事实上,许多遗传疾病的起因是替代性拼接而不是序列中的突变。 但是,替代拼接将在不损失原始蛋白质的情况下产生蛋白质变体,从而为新变体适应新功能开辟可能性。 基因复制以类似的方式在新功能的演变中发挥了重要作用,它提供了可以在不消灭原始功能蛋白的情况下进化的基因。
链接到学习
观看此视频中的实际过程,可视化 mRNA 拼接是如何发生的。 NDSU 虚拟细胞动画项目动画 “mRNA 拼接”。
控制 RNA 的稳定性
在 mRNA 离开细胞核之前,它会被赋予两个保护性的 “帽子”,以防止链末端在旅途中降解。 5'帽位于mRNA的5'末端,通常由甲基化的三磷酸鸟苷分子(GTP)组成。 附着在 3' 末端的 Poly-a 尾巴通常由一系列腺嘌呤核苷酸组成。 一旦RNA被运送到细胞质,RNA在那里停留的时间长度就可以得到控制。 每个 RNA 分子都有明确的寿命并以特定的速率衰变。 这种衰变速度会影响细胞中蛋白质的含量。 如果衰变率增加,RNA在细胞质中就不会存在那么长时间,从而缩短了翻译发生的时间。 相反,如果衰变率降低,RNA分子在细胞质中的停留时间将更长,并且可以翻译更多的蛋白质。 这种衰变速率被称为 RNA 稳定性。 如果 RNA 稳定,则将在细胞质中检测到更长时间。
蛋白质与 RNA 的结合会影响其稳定性。 称为 RNA结合蛋白或RBP的蛋白质可以与蛋白质编码区域的上游或下游的RNA区域结合。 RNA 中这些未转化为蛋白质的区域称为未翻译区域或 UTR。 它们不是内含子(它们已在原子核中被移除)。 相反,这些区域是调节 mRNA 定位、稳定性和蛋白质翻译的区域。 蛋白质编码区域之前的区域被称为 5' UTR,而编码区域之后的区域称为 3' UTR(图\(\PageIndex{3}\))。 RBP 与这些区域的结合会增加或降低 RNA 分子的稳定性,具体取决于结合的特定 RBP。
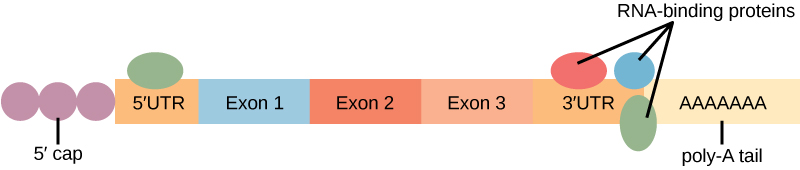
RNA 稳定性和 microRNA
除了结合和控制(增加或降低)RNA 稳定性的 RBP 外,其他称为 microRNA 的元素也可以与 RNA 分子结合。 这些 m icroRN A 或 miRNA 是短的 RNA 分子,长度只有 21—24 个核苷酸。 miRNA 是在细胞核中作为更长的 miRNA 之前生成的。 这些 pre-miRNA 被一种叫做 dicer 的蛋白质切成成熟的 miRNA。 像转录因子和RBP一样,成熟的miRNA可以识别特定的序列并与RNA结合;但是,miRNA也与一种称为R NA诱导沉默复合物(RISC)的核糖核蛋白复合物有关。 RISC 与 miRNA 结合以降解靶向 mRNA。 miRNA 和 RISC 复合物共同迅速摧毁 RNA 分子。
摘要
转录后控制可以在转录后的任何阶段发生,包括 RNA 剪接、核穿梭和 RNA 稳定性。 一旦转录了RNA,就必须对其进行处理以生成可以翻译的成熟RNA。 这涉及去除不编码蛋白质的内含子。 Spliceosomes 绑定到标记外显子/内含子边界的信号以移除内含子并将外显子连接在一起。 一旦发生这种情况,RNA就会成熟并且可以翻译。 RNA 是在细胞核中产生和拼接的,但需要输送到细胞质进行翻译。 RNA 通过核孔复合体输送到细胞质。 一旦 RNA 进入细胞质,它在降解之前存在的时间长度(称为 RNA 稳定性)也可以改变,以控制合成的蛋白质总量。 RNA稳定性可以增加,从而延长在细胞质中的停留时间,也可以缩短,从而缩短时间并减少蛋白质合成。 RNA 的稳定性由 RNA 结合蛋白 (rpB) 和 microRNA (miRNA) 控制。 这些 rpb 和 miRNA 与 RNA 的 5' UTR 或 3' UTR 结合,以增加或降低 RNA 的稳定性。 根据 RBP,稳定性可以显著增加或降低;但是,miRNA 总是会降低稳定性并促进衰变。
词汇表
- 3' UTR
- 3' 未翻译区域;未翻译的 RNA 分子中蛋白质编码区的下游区域
- 5' 帽子
- 一种甲基化的三磷酸鸟苷 (GTP) 分子,附着在信使 RNA 的 5' 末端以保护末端免受降解
- 5' UTR
- 5' 未翻译区域;未翻译的 RNA 分子中蛋白质编码区的上游区域
- 切块机
- 将前 miRNA 切成成成熟形式的 miRNA 的酶
- microRNA (miRNA)
- 与 RNA 分子结合以降解的小型 RNA 分子(长度约为 21 个核苷酸)
- poly-a 尾巴
- 一系列附着在 mRNA 3' 末端的腺嘌呤核苷酸,以保护末端免受降解
- RNA 结合蛋白 (RBP)
- 与 3' 或 5' UTR 结合以增加或降低 RNA 稳定性的蛋白质
- RNA 稳定性
- RNA 分子将在细胞质中保持完整多长时间
- 未翻译的区域
- 未转化为蛋白质的 RNA 分子片段。 这些区域位于蛋白质编码区之前(上游或 5')和之后(下游或 3')
- RISC
- 与 miRNA 结合到 RNA 以降解的蛋白质复合物