
11.2: 有性生殖
- Page ID
- 202843

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 解释减数分裂和有性生殖是进化的特征
- 将后代之间的变异确定为有性生殖的潜在进化优势
- 描述性多细胞生物的三种不同的生命周期类型及其共同点
有性生殖是真核细胞出现后的早期进化创新。 它似乎非常成功,因为大多数真核生物都能够通过性繁殖,而在许多动物中,它是唯一的繁殖方式。 然而,科学家们认识到了有性生殖的一些真正缺点。 从表面上看,创造作为父母遗传克隆的后代似乎是一个更好的系统。 如果亲本生物成功占领栖息地,则具有相同特征的后代也将同样成功。 在无性萌芽、碎片化或无性卵的有利环境下,生物体也具有明显的好处,它可以产生后代。 这些繁殖方法不需要另一种异性生物。 事实上,一些过着孤独生活方式的生物保留了无性繁殖的能力。 此外,在无性人群中,每个人都有繁殖能力。 在性人群中,雄性不是自己产生后代,因此从理论上讲,无性种群的增长速度可能是原来的两倍。
但是,完全依赖无性繁殖的多细胞生物极为罕见。 为什么性(和减数分裂)如此普遍? 这是生物学中悬而未决的重要问题之一,也是二十世纪下半叶开始的许多研究的重点。 有几种可能的解释,其中之一是有性生殖在后代中产生的变异对于种群的生存和繁殖非常重要。 因此,平均而言,有性生殖的人群将比原本相似的无性繁殖人口留下的后代更多。 无性生物变异的唯一来源是突变。 这是性生物变异的最终来源,但除此之外,当不同的亲本将他们独特的基因组组合在一起,并且这些基因在第一阶段通过交叉和随机混合成不同的组合时,这些不同的突变会不断地从一代转移到下一代中间阶段的分类 I
进化连接:红皇后假说
毫无疑问,有性生殖为利用这种机制产生后代的生物提供了进化优势。 但是,即使面对相当稳定的条件,为什么有性生殖在个体生物更加困难和成本更高的情况下仍能持续下去? 变异是有性生殖的结果,但为什么需要持续的变异呢? 进入 Leigh Van Valen 于 1973 年首次提出的红皇后假说。 1 这个概念是根据刘易斯·卡罗尔的著作《透过窥镜》中的红皇后种族命名的。
所有物种都与其他生物共同进化;例如,捕食者与猎物一起进化,寄生虫与宿主一起进化。 通过有利变异获得的每一个微小优势都会使物种比紧密的竞争对手、捕食者、寄生虫甚至猎物更具优势。 让共同进化的物种保持自身资源份额的唯一方法是不断提高其适应性。 当一个物种获得优势时,这会增加对其他物种的选择;它们还必须发展优势,否则它们将被击败。 没有哪个物种能向前发展太远,因为有性生殖后代之间的遗传变异为所有物种提供了快速改善的机制。 无法跟上步伐的物种就会灭绝。 红皇后的口号是:“你需要尽一切努力才能待在同一个地方。” 这是对竞争物种之间共同进化的恰当描述。
性繁殖生物的生命周期
受精和减数分裂在性生命周期中交替出现。 这两个事件之间会发生什么取决于生物体。 减数分裂过程将染色体数量减少一半。 受精,即两个单倍体配子的结合,可以恢复二倍体状态。 多细胞生物的生命周期主要分为三类:二倍体占主导地位,其中多细胞二倍体阶段是最明显的生命阶段,例如包括人类在内的大多数动物;单倍体占主导地位,其中多细胞单倍体阶段最多显而易见的生命阶段,例如所有真菌和一些藻类;以及世代交替,这两个阶段因群体的不同而在不同程度上显而易见,就像植物和某些藻类一样。
二倍体占主导地位的生命周期
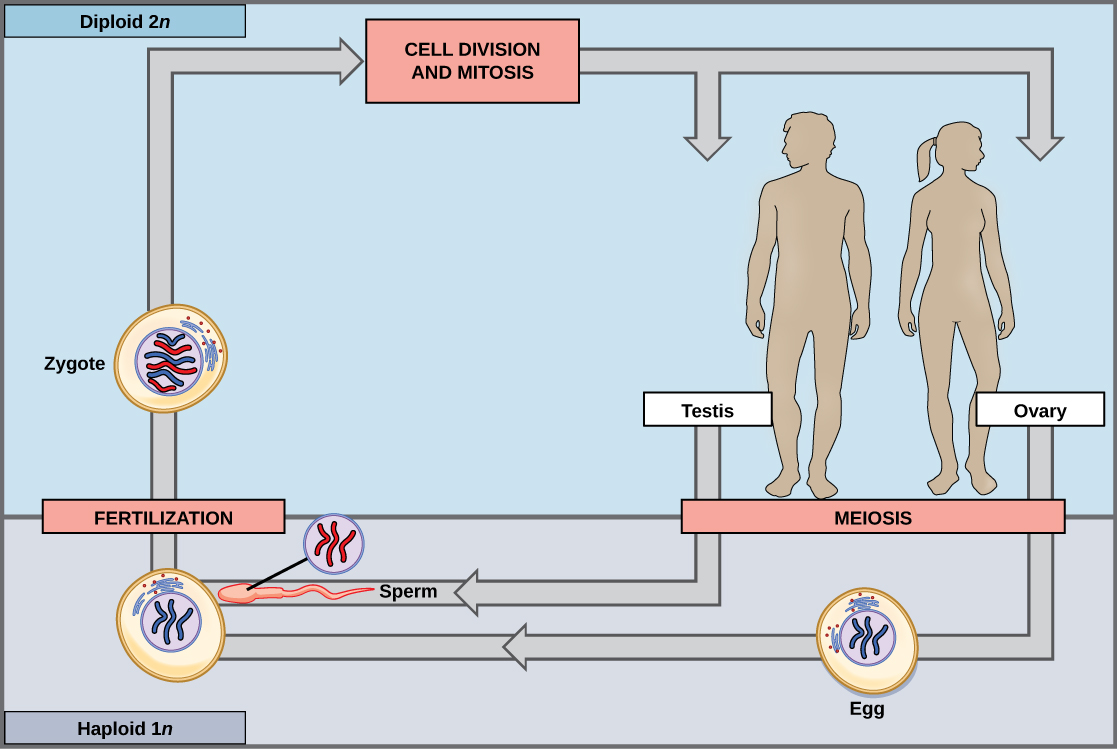
几乎所有动物都采用二倍体占主导地位的生命周期策略,其中生物体产生的唯一单倍体细胞是配子。 在胚胎发育的早期,专门的二倍体细胞(称为生殖细胞)会在性腺内产生,例如睾丸和卵巢。 生殖细胞能够进行有丝分裂以延续细胞系,减数分裂产生配子。 一旦单倍体配子形成,它们就会失去再次分裂的能力。 没有多细胞单倍体的生命阶段。 受精是通过两个配子(通常来自不同个体)融合而发生的,从而恢复二倍体状态(图\(\PageIndex{1}\))。
单倍体占主导地位的生命周期
大多数真菌和藻类采用生命周期类型,其中生物的 “身体”(生命周期中重要的生态部分)是单倍体。 构成显性多细胞阶段组织的单倍体细胞是由有丝分裂形成的。 在有性生殖过程中,来自两个个体的特殊单倍体细胞(指定为(+)和(−)交配类型,结合形成二倍体合子。 合子立即经历减数分裂,形成四个称为孢子的单倍体细胞。 尽管单倍体像 “父母” 一样,但这些孢子含有来自两个父母的新遗传组合。 孢子可以在不同的时间段内保持休眠状态。 最终,当条件有利时,孢子会通过多轮有丝分裂形成多细胞单倍体结构(图\(\PageIndex{2}\))。
艺术连接
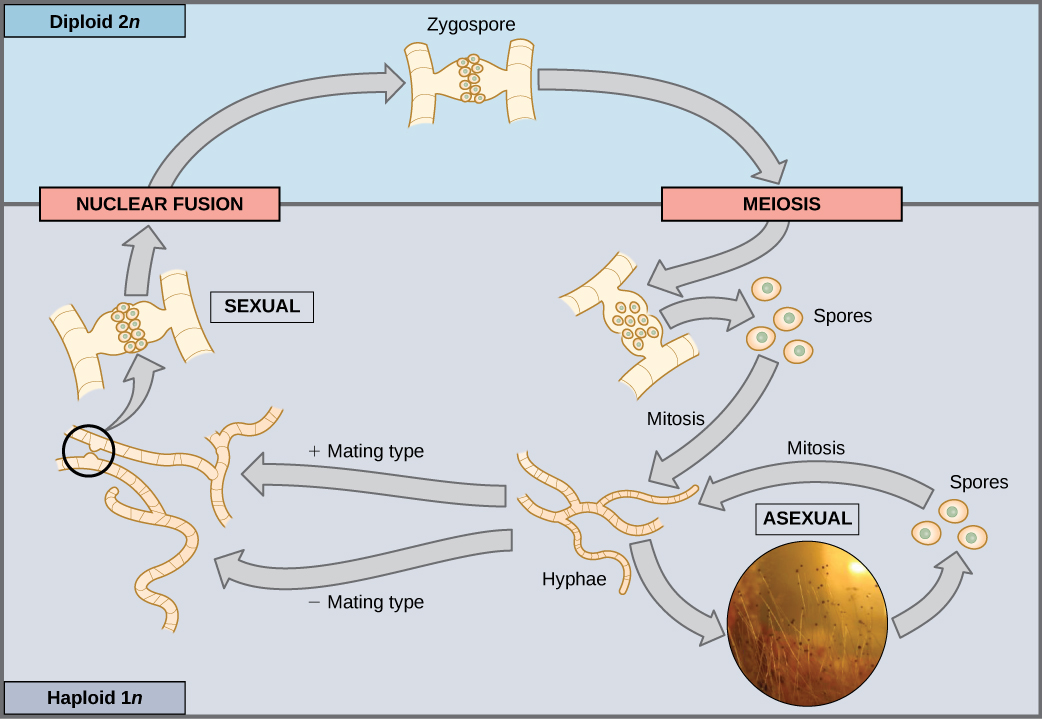
如果发生突变导致真菌无法再产生负交配型,它还能繁殖吗?
世代交替
第三种生命周期类型被一些藻类和所有植物所采用,是单倍体占主导地位和二倍体占主导地位的极端情况的混合体。 世代交替的物种在其生命周期中既有单倍体又有二倍体多细胞生物。 单倍体多细胞植物被称为配子体,因为它们从特殊细胞中产生配子。 在这种情况下,减数分裂并不直接参与配子的产生,因为产生配子的生物已经是单倍体了。 配子之间的受精形成二倍体合子。 合子将经历多轮有丝分裂,并产生一种叫做孢子体的二倍体多细胞植物。 孢子体的特殊细胞将经历减数分裂并产生单倍体孢子。 孢子随后会发育成配子体(图\(\PageIndex{3}\))。
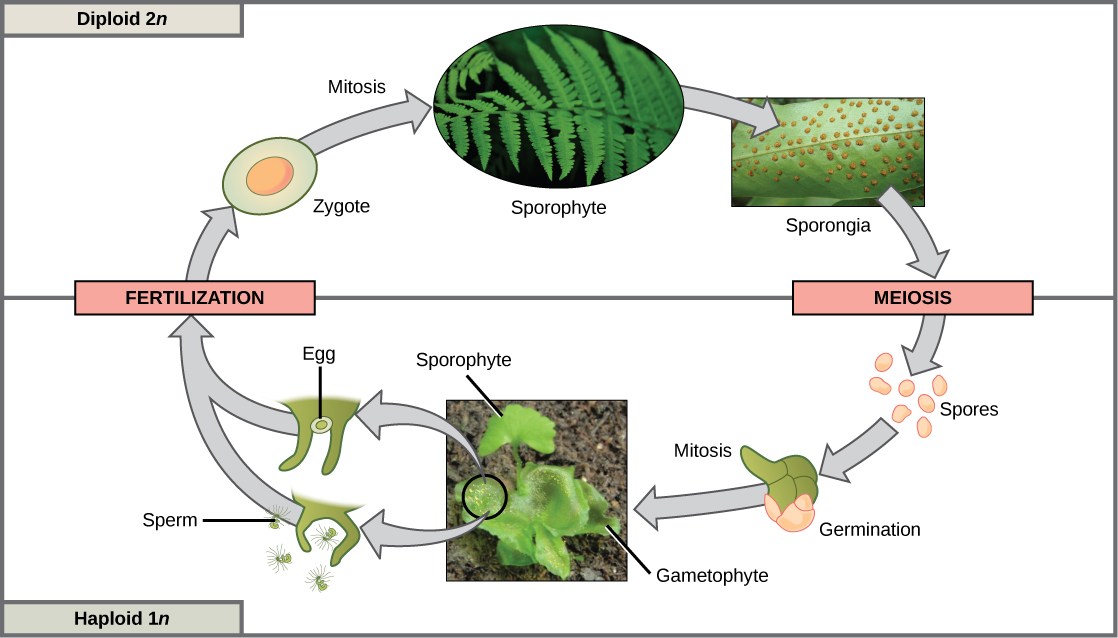
尽管所有植物都使用某种版本的世代交替,但孢子体和配子体的相对大小以及它们之间的关系差异很大。 在苔藓等植物中,配子体生物是自由生活的植物,而孢子体在物理上依赖配子体。 在其他植物中,例如蕨类植物,配子体和孢子体植物都是自由生活的;但是,孢子体要大得多。 在种子植物中,例如木兰树和雏菊,配子体仅由几个细胞组成,就雌配子体而言,配子体完全保留在孢子体中。
有性生殖在多细胞生物中有多种形式。 但是,在每种生命周期的某个时刻,减数分裂会产生单倍体细胞,这些细胞将与另一种生物的单倍体细胞融合。 变异机制——交叉、同源染色体的随机分类和随机受精——存在于所有版本的有性生殖中。 地球上几乎所有多细胞生物都使用有性生殖这一事实有力地证明了产生具有独特基因组合的后代的好处,尽管还有其他可能的好处。
摘要
几乎所有的真核生物都经过有性繁殖。 减数分裂引入生殖细胞的变异似乎是有性生殖的优势之一,它使之如此成功。 减数分裂和受精在性生命周期中交替出现。 减数分裂过程产生称为配子的独特生殖细胞,其染色体数量是母细胞的一半。 受精,即来自两个个体的单倍体配子的融合,可以恢复二倍体状态。 因此,有性繁殖的生物在单倍体和二倍体阶段之间交替出现。 但是,生殖细胞的产生方式以及减数分裂和受精之间的时间差异很大。 生命周期主要分为三类:二倍体占主导地位,由大多数动物证实;单倍体占主导地位,由所有真菌和某些藻类证明;以及世代交替,由植物和某些藻类证明。
艺术联系
图\(\PageIndex{2}\):如果发生突变导致真菌无法再产生负交配型,它还能繁殖吗?
- 回答
-
是的,它将能够无性繁殖。
脚注
- 1 Leigh Van Valen,《新进化定律》,《进化论 1》(1973):1—30
词汇表
- 世代相传
- 二倍体和单倍体阶段交替的生命周期类型
- 二倍体占主导地位
- 多细胞二倍体阶段普遍存在的生命周期类型
- 单倍体占主导地位
- 多细胞单倍体阶段普遍存在的生命周期类型
- 配子体
- 产生配子的多细胞单倍体生命周期阶段
- 生殖细胞
- 产生配子的特殊细胞系,例如卵子或精子
- 生命周期
- 生物体发育过程中的事件顺序和产生后代的细胞的产生
- 孢子体
- 多细胞二倍体生命周期阶段,通过减数分裂产生单倍体孢子