
11.1: 减数分裂的过程
- Page ID
- 202844

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 描述减数分裂期间染色体的行为
- 描述减数分裂期间的细胞事件
- 解释减数分裂和有丝分裂的区别
- 解释减数分裂内部在减数分裂产物之间产生遗传变异的机制
有性生殖需要受精,即来自两个单独生物体的两个细胞的结合。 如果这两个细胞各包含一组染色体,则生成的细胞包含两组染色体。 单倍体细胞包含一组染色体。 含有两组染色体的细胞被称为二倍体。 细胞中染色体组的数量称为其倍体水平。 如果要继续繁殖周期,那么二倍体细胞必须以某种方式减少其染色体组的数量,然后才能再次受精,否则每代染色体组的数量将持续翻一番。 因此,除了受精外,有性生殖还包括减少染色体组数量的核分裂。
大多数动植物是二倍体,包含两组染色体。 在生物体的每个体细胞(除配子或生殖细胞之外的所有多细胞生物体细胞)中,细胞核包含每条染色体的两个副本,称为同源染色体。 体细胞有时被称为 “身体” 细胞。 同源染色体是配对的染色体,其长度上位于相同位置的相同基因。 二倍体生物从每个亲本那里继承每条同源染色体的一份副本;总而言之,它们被视为一整套染色体。 包含每条同源染色体的单个副本的单倍体细胞仅存在于产生配子或孢子的结构中。 孢子是单倍体细胞,可以产生单倍体生物,也可以与另一个孢子融合形成二倍体细胞。 所有动物和大多数植物都会产生卵子和精子或配子。 有些植物和所有真菌都会产生孢子。
形成单倍体细胞的核分裂称为减数分裂,与有丝分裂有关。 如你所知,有丝分裂是细胞繁殖周期的一部分,它会产生相同的子核,这些子核在基因上也与原始亲本核相同。 在有丝分裂中,母核和子核处于相同的倍体水平——大多数动植物都是二倍体。 减数分裂采用许多与有丝分裂相同的机制。 但是,起始核始终是二倍体,减数分裂细胞分裂结束时产生的核是单倍体。 为了减少染色体数量,减数分裂包括一轮染色体复制和两轮核分裂。 由于在每个分裂阶段发生的事件都类似于有丝分裂事件,因此分配了相同的阶段名。 但是,由于分为两轮,主要过程和阶段以 “I” 或 “II” 指定。 因此,减数分裂 I 是减数分裂的第一轮,由前期 I、prometaphase I 等组成。 减数分裂 I I 是第二轮减数分裂发生的,包括前期 II、prometaphase II 等。
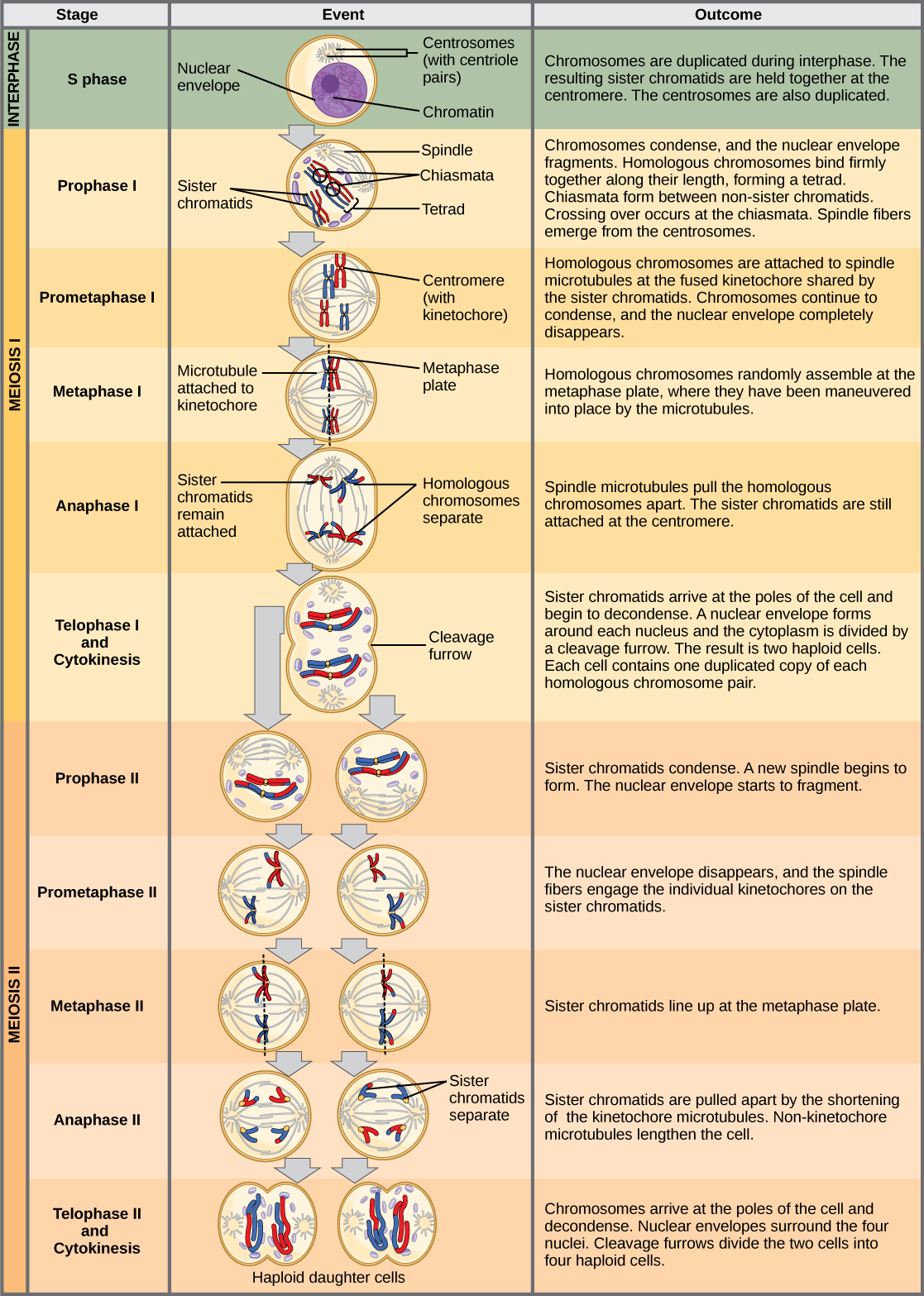
减数分裂 I
减数分裂之前是由 G 1、S 和 G 2 相组成的间期,它们与有丝分裂之前的阶段几乎相同。 G 1 阶段,也称为第一间隙阶段,是中间阶段的第一阶段,侧重于细胞生长。 S 期是中间阶段的第二阶段,在此期间,染色体的 DNA 被复制。 最后,G 2 阶段,也称为第二间隙阶段,是中间阶段的第三个也是最后一个阶段;在这个阶段,细胞为减数分裂做最后的准备。
在 S 期的 DNA 复制过程中,每条染色体都会被复制以产生两个相同的拷贝,称为姐妹染色体,它们由粘合素蛋白在着丝粒处聚集在一起。 Cohesin 将染色体保持在一起直到 anaphase II。 中心体是组织减数分裂主轴微管的结构,也可以复制。 这使细胞为进入第一阶段,即第一个减数分裂阶段做好准备。
Prophase I
在第一阶段的早期,在显微镜下可以清楚地看到染色体之前,同源染色体的尖端会被蛋白质附着在核包膜上。 随着核包膜开始分解,与同源染色体相关的蛋白质使这对染色体彼此靠近。 回想一下,在有丝分裂中,同源染色体不会配对在一起。 在有丝分裂中,同源染色体端到端排列,因此当它们分裂时,每个子细胞都会从同源对的两个成员那里获得一个姊妹染色体。 突触复合物是同源染色体之间的蛋白质格子,首先在特定位置形成,然后扩散到覆盖染色体的整个长度。 同源染色体的紧密配对称为突触。 在突触中,同源染色体染色体染色体上的基因彼此精确对齐。 synaptonemal 复合物支持非姐妹同源染色体之间染色体片段的交换,这种过程称为交叉。 交换后可以直观地观察到交叉为 chiasmata(单数 = chiasma)(图\(\PageIndex{1}\))。
在人类等物种中,尽管 X 和 Y 性染色体不是同源的(它们的大多数基因不同),但它们的同源区域很小,允许 X 和 Y 染色体在 I 前期配对。部分突触复合体仅在同源区域之间生长。
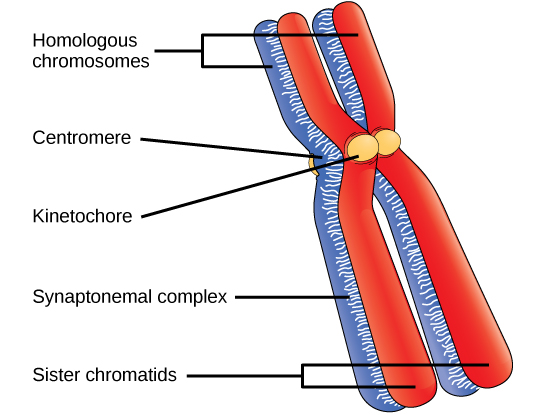
沿着突触复合体间隔分布着称为重组结节的大型蛋白质组件。 这些组合标志着后来的奇亚斯玛塔的要点,并介导了非姐妹染色体之间交叉或基因重组的多步过程。 在每个染色体上的重组结节附近,双链 DNA 被分解,切口被修改,非姐妹染色体之间建立了新的连接。 随着第一阶段的进展,突触复合体开始分解,染色体开始凝结。 当突触复合物消失后,同源染色体在着丝粒和 chiasmata 处仍然相互附着。 chiasmata 会一直持续到 anaphase I。chiasmata 的数量因种类和染色体的长度而异。 在减数分裂 I 期间,每条染色体必须至少有一个 chiasma 才能正确分离同源染色体,但可能多达 25 个。 交叉后,synaptonemal 复合物会分解,同源对之间的粘合连接也被移除。 在第一阶段结束时,两对仅在 chiasmata 处聚集在一起(图\(\PageIndex{2}\)),之所以被称为 t etrads,是因为每对同源染色体的四个姐妹染色体现在都可见。
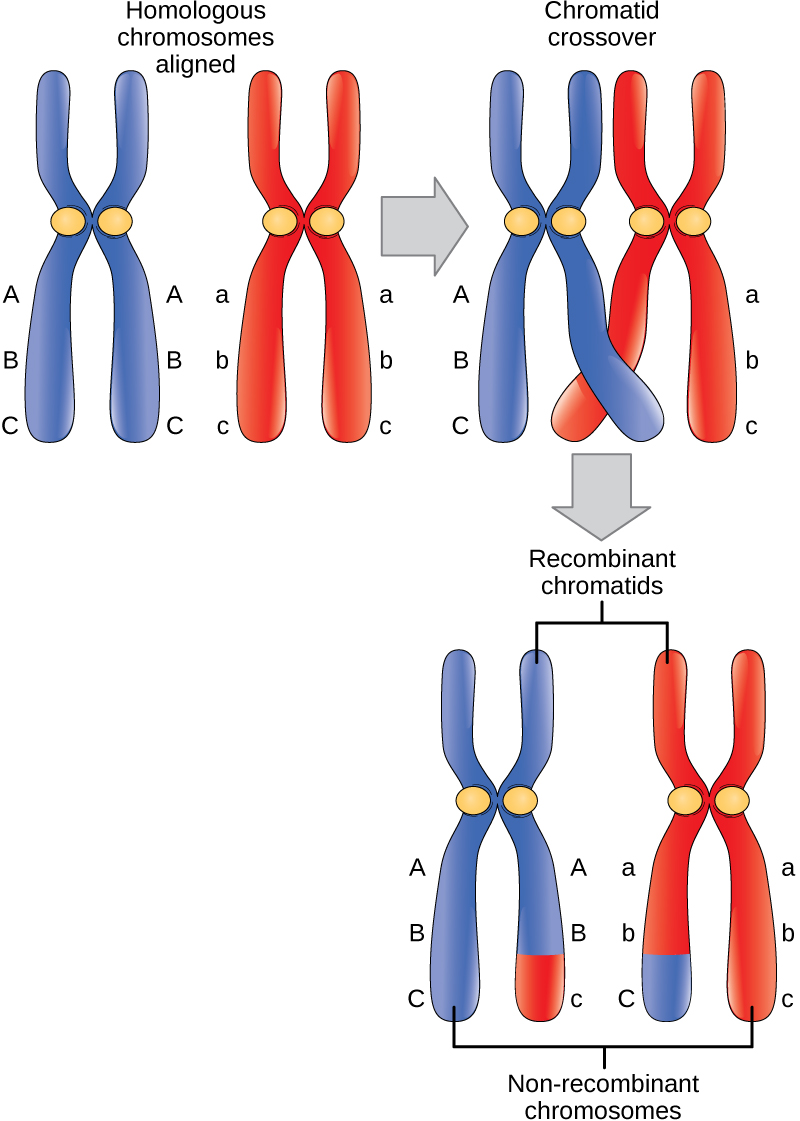
交叉事件是减数分裂产生的核中遗传变异的第一个来源。 同源非姐妹染色体之间的单次交叉事件会导致母体染色体和父系染色体之间等效DNA的相互交换。 现在,当那个姐妹染色体被转移到配子细胞中时,它将携带来自个体父母一方的一些DNA和来自另一个亲本的一些DNA。 姊妹重组染色体具有母系和父系基因的组合,这些基因在交叉之前并不存在。 染色体一臂中的多个交叉具有相同的效果,即交换 DNA 片段以创建重组染色体。
Prometaphase I
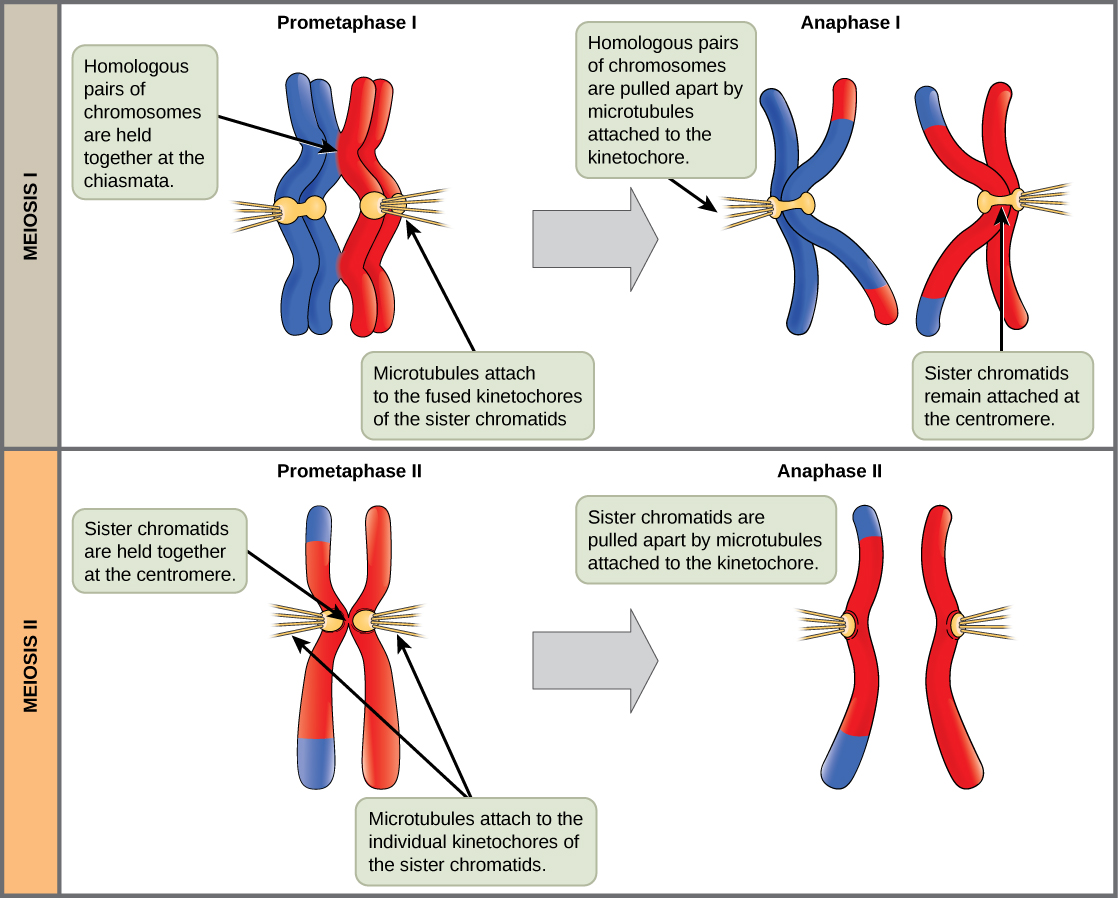
prometaphase I 中的关键事件是纺锤纤维微管附着在 centromeres 的 kinetochore 蛋白上。 Kinetochore 蛋白是将染色体的中心点与有丝分裂纺锤的微管结合的多蛋白复合物。 微管从放置在细胞两极的中心体中生长。 微管向细胞中间移动,附着在两条融合的同源染色体中的一条上。 微管附着在每条染色体的动力学上。 当同源对中的每个成员都附着在细胞的两极上时,在下一阶段,微管可以将同源对分开。 附着在 kinetochore 上的纺锤纤维被称为 kinetochore 微管。 在 prometaphase I 的末尾,每个 tetrad 都附着在两极的微管上,每个极点都有一条同源染色体。 在 chiasmata,同源染色体仍然保持在一起。 此外,核膜已经完全分解。
Metachase I
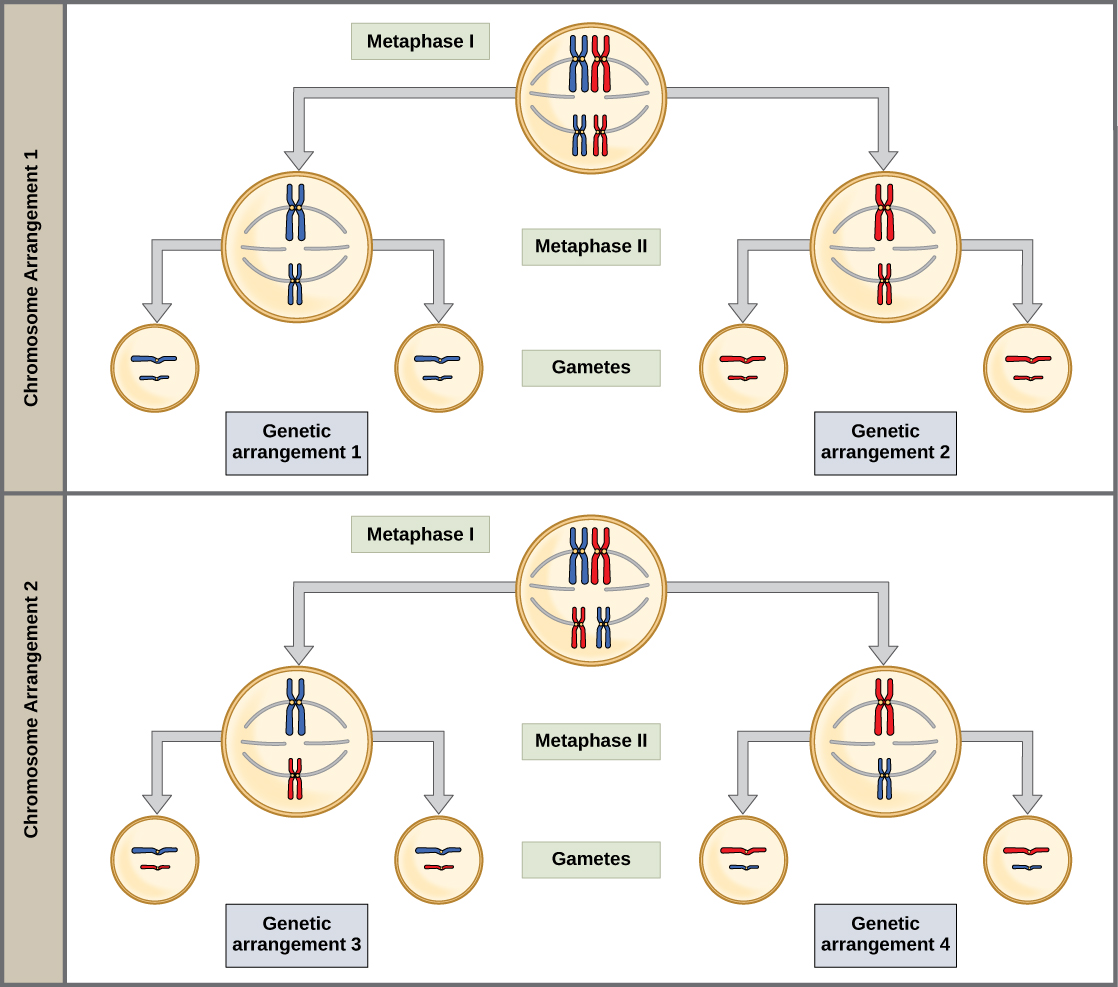
在第一中期,同源染色体排列在细胞中心,动力学朝向相反的两极。 同源对将自己随机定向在赤道。 例如,如果 1 号染色体的两个同源成员被标记为 a 和 b,则染色体可以排列 a-b 或 b-a。 这对于确定配子携带的基因很重要,因为每个配子只能获得两条同源染色体中的一条。 回想一下,同源染色体并不相同。 它们的遗传信息略有不同,导致每个配子都有独特的基因组成。
这种随机性是产生第二种形式的后代遗传变异的物理基础。 考虑一下,有性繁殖生物体的同源染色体最初是作为两组独立的染色体遗传的,每个亲本各有一组。 以人类为例,母亲捐赠的卵中存在一组23条染色体。 父亲在精子中提供另一组23条染色体,使卵子受精。 多细胞后代的每个细胞都有原始两组同源染色体的副本。 在减数分裂的前期第一阶段,同源染色体形成四边体。 在中期 I 中,这些对在细胞两极之间的中间点排成一列,形成中期板。 由于微管纤维遇到母系或父系遗传的染色体的可能性相同,因此四边形在中期板上的排列是随机的。 任何母系遗传的染色体都可能面对任一极点。 任何父系遗传的染色体也可能面对任一极点。 每个四边形的方向与其他 22 个四边形的方向无关。
这个事件——中间板块上随机(或独立)分类的同源染色体——是引入配子或孢子变异的第二种机制。 在每个经历减数分裂的细胞中,四边体的排列是不同的。 变异的数量取决于组成一个集合的染色体的数量。 在中期板上定向有两种可能性;因此,可能的对齐次数等于 2 n,其中 n 是每组染色体的数量。 人类有23对染色体,这就产生了超过八百万(2 23)个可能具有遗传差异的配子。 这个数字不包括先前通过交叉在姊妹染色体中产生的变异性。 鉴于这两种机制,减数分裂产生的任何两个单倍体细胞极不可能具有相同的遗传成分(图\(\PageIndex{3}\))。
总结减数分裂 I 的遗传后果,母系和父系基因通过第一阶段前期每对同源物之间发生的交叉事件进行重组。此外,中期板上随机分类的四边形会产生母系和父系染色体的独特组合这将进入配子。
Anaphase I
在 anaphase I 中,微管将连接的染色体分开。 姐妹染色体在着丝粒处仍然紧密结合在一起。 chiasmata 在 anaphase I 中被破坏,因为附着在融合动力学上的微管将同源染色体分开(图\(\PageIndex{4}\))。
Telophase I 和细胞分裂
在末期,分离的染色体到达相反的两极。 其余的典型末期事件可能会发生,也可能不会发生,具体取决于物种。 在某些生物体中,染色体解密,在末期 I 的染色体周围形成核包膜。在其他生物体中,细胞分裂(将细胞质成分物理分离成两个子细胞)发生在没有细胞核重构的情况下。 在几乎所有种类的动物和某些真菌中,细胞分裂作用通过分裂沟(肌动蛋白环的收缩导致细胞质分裂)分离细胞内容物。 在植物中,细胞分裂过程中由高尔基囊泡融合在中叶板上形成细胞板。 这种细胞板最终将导致形成分隔两个子细胞的细胞壁。
两个单倍体细胞是第一个减数分裂的最终结果。 这些细胞是单倍体,因为在每个极点,每对同源染色体中只有一对。 因此,只有一整套染色体存在。 这就是为什么细胞被视为单倍体的原因——尽管每个同系物仍然由两个姐妹染色体组成,但只有一组染色体。 回想一下,姐妹染色体只是两条同源染色体中的一条的重复体(交叉过程中发生的变化除外)。 在减数分裂 II 中,这两个姐妹染色体将分离,产生四个单倍体子细胞。

减数分裂 II
在某些物种中,细胞在进入减数分裂 II 之前会进入短暂的中间阶段或相互作用。 Interkinesis 缺少 S 期,因此染色体不会重复。 减数分裂 I 中产生的两个细胞同步经历减数分裂 II 的事件。 在减数分裂 II 期间,两个子细胞内的姊妹染色体分离,形成四个新的单倍体配子。 减数分裂 II 的机制与有丝分裂相似,不同的是每个分裂细胞只有一组同源染色体。 因此,作为经历有丝分裂的二倍体细胞,每个细胞有待分离出的姊妹染色体数量的一半。
前期二
如果染色体在末期第一阶段解密,它们会再次凝结。 如果形成了核包膜,它们就会分裂成囊泡。 运动间复制的中心体相互移动,向相反的极点移动,然后形成新的主轴。
Prometapase II
核包络被完全分解,主轴完全成型。 每个姐妹 chromatid 都会形成一个单独的 kinetochore,它从相反的极点附着在微管上。
中期 II
姐妹染色体在细胞赤道处最大限度地凝结并对齐。
Anaphase II
姊妹染色体被 kinetochore 微管拉开,然后向相反的极点移动。 非 Kinetochore 微管会拉长细胞。
Telophase II 和细胞分裂
染色体到达相反的两极并开始解密。 核包膜在染色体周围形成。 细胞分裂作用将两个细胞分成四个独特的单倍体细胞。 此时,新形成的原子核都是单倍体。 所产生的细胞在基因上是独一无二的,因为父系和母系同系物是随机分类的,也因为在交叉过程中染色体的母系和父系片段(及其基因组)会重组。 减数分裂的整个过程如图所示\(\PageIndex{5}\)。
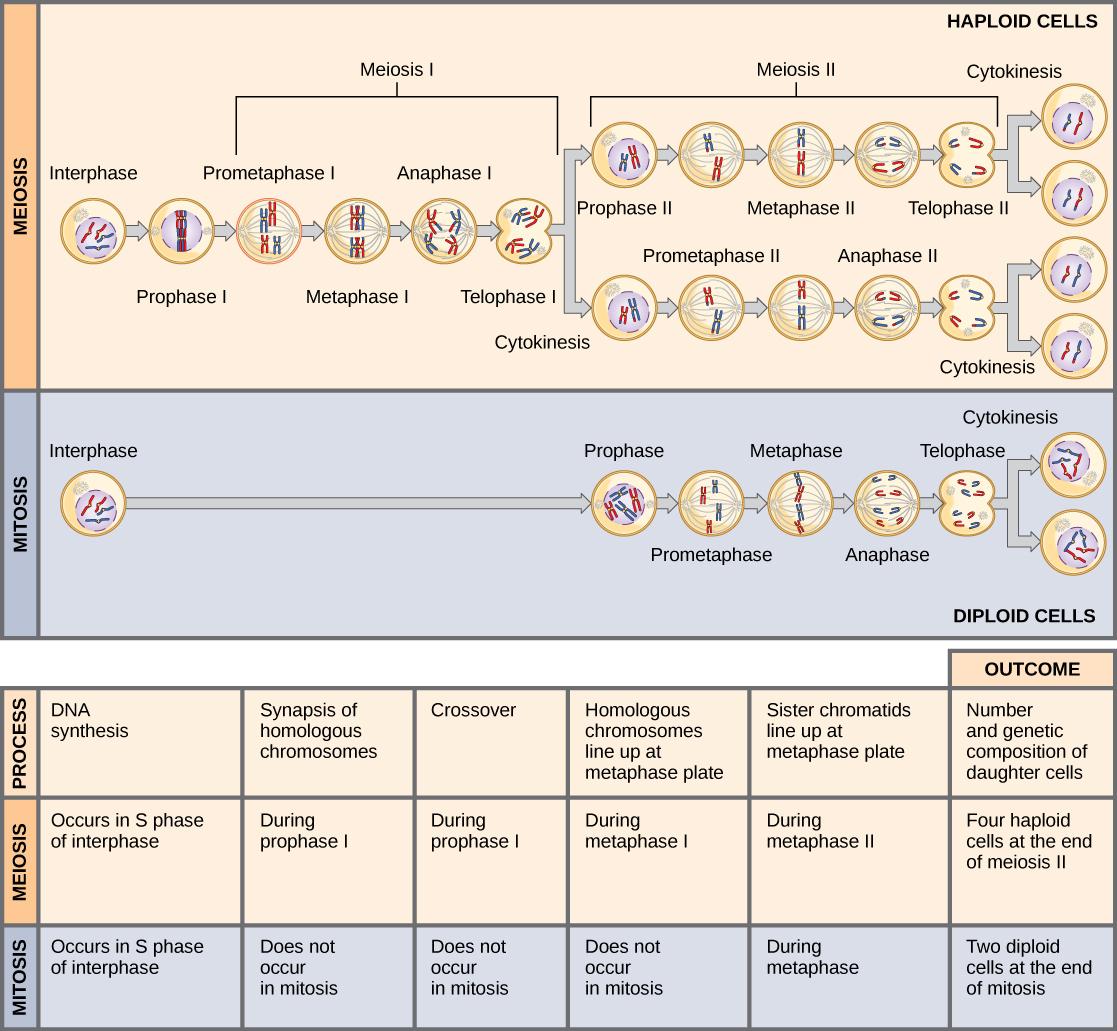
比较减数分裂和有丝分裂
有丝分裂和减数分裂都是真核细胞中细胞核分裂的形式。 它们有一些相似之处,但也表现出明显的差异,导致结果截然不同(图\(\PageIndex{6}\))。 有丝分裂是一种单核分裂,它会产生两个核,通常分成两个新细胞。 有丝分裂分裂产生的细胞核在基因上与原始细胞核相同。 它们具有相同数量的染色体集,单倍体细胞为一组,二倍体细胞为两组。 在大多数植物和所有动物物种中,通常是二倍体细胞经历有丝分裂形成新的二倍体细胞。 相比之下,减数分裂由两个核分裂组成,形成四个核,通常分成四个新细胞。 减数分裂产生的细胞核在基因上并不相同,它们只包含一组染色体。 这是原始细胞(二倍体)中染色体组数的一半。
有丝分裂和减数分裂的主要区别在于减数分裂 I,减数分裂与有丝分裂是截然不同的核分裂。 在 meiosis I 中,同源染色体对相互关联,与突触复合体结合在一起,产生 chiasmata 并在姊妹染色体之间进行交叉,然后沿着四边形的中期板排成四边形,来自相反主轴极的 kinetochore 纤维附着四边形中同系物的每种动力学。 所有这些事件只发生在减数分裂 I 中。
当 chiasmata 消失并且四边形被分解,同系物移至一个极点或另一个极点时,倍体水平(未来每个原子核中的染色体组数量)已从两组减少到一组。 出于这个原因,减数分裂 I 被称为减数分裂。 在有丝分裂期间,倍体水平没有这样的降低。
减数分裂 II 更类似于有丝分裂分裂。 在这种情况下,重复的染色体(只有一组)在中期板上排成一列,分开的动力学从相反的极点附着在kinetochore纤维上。 在 anaphase II 期间,就像在有丝分裂后期一样,动力学分裂,一个姐妹染色体(现在称为染色体)被拉到一极,而另一个姐妹染色体被拉到另一极。 如果不是因为存在交叉,那么每个减数分裂 II 分裂的两个产物将是相同的(比如在有丝分裂中)。 相反,它们是不同的,因为每条染色体一直至少有一个交叉点。 减数分裂 II 不是还原分裂,因为尽管生成的细胞中的基因组拷贝较少,但仍然有一组染色体,就像减数分裂 I 末期一样
进化连接:减数分裂进化的奥秘
生物的某些特征是如此广泛和基本,以至于有时很难记住它们像其他更简单的特征一样进化。 减数分裂是一系列极其复杂的细胞事件,以至于生物学家很难假设和测试减数分裂可能是如何演变的。 尽管减数分裂与有性生殖及其优缺点密不可分,但将减数分裂的演变和性别的进化问题区分开来很重要,因为早期减数分裂的优势可能与现在不同。 跳出框框思考,想象减数分裂的早期好处可能是什么,这是揭示减数分裂可能是如何演变的一种方法。
减数分裂和有丝分裂具有明显的细胞过程,减数分裂是由有丝分裂演变而来的,这是有道理的。 困难在于减数分裂 I 和有丝分裂之间的明显区别。 亚当·威尔金斯和罗宾·霍利迪 1 总结了减数分裂从有丝分裂演变为需要发生的独特事件。 这些步骤是同源染色体配对、交叉交换、在后期阶段保持姐妹染色体附着以及抑制相间DNA复制。 他们认为第一步是最艰难和最重要的,了解它是如何演变的将使进化过程更加清晰。 他们建议进行基因实验,这些实验可能会揭示突触的演变。
还有其他方法可以理解减数分裂的演变正在进行中。 单细胞原生生物中存在不同形式的减数分裂。 有些似乎是更简单或更 “原始” 的减数分裂形式。 比较不同原生生物的减数分裂可能揭示减数分裂的演变。 Marilee Ramesh 及其同事 2 比较了原生生物减数分裂所涉及的基因,以了解减数分裂可能在何时何地进化。 尽管研究仍在进行中,但最近对原生生物减数分裂的奖学金表明,减数分裂的某些方面可能比其他方面发展得晚。 这种遗传比较可以告诉我们减数分裂的哪些方面是最古老的,以及它们可能从早期的细胞中借鉴了哪些细胞过程。
摘要
有性生殖要求二倍体生物产生单倍体细胞,这些细胞可以在受精过程中融合形成二倍体后代。 与有丝分裂一样,DNA复制发生在细胞周期的S阶段减数分裂之前。 减数分裂是一系列将染色体和染色体排列成子细胞并将其分离成子细胞的事件。 在减数分裂的中间阶段,每条染色体都是重复的。 在减数分裂中,有两轮核分裂产生四个核,通常是四个子细胞,每个子细胞的染色体数量是母细胞的一半。 第一种分离同系物,第二种(如有丝分裂)将染色体分离成单个染色体。 在减数分裂期间,子核的变异是由于第一阶段的交叉和中期 I 时四分体的随机对齐而引入的。减数分裂产生的细胞在基因上是独一无二的。
减数分裂和有丝分裂有相似之处,但结果却不同。 有丝分裂分裂是单核分裂,它产生的子核在基因上相同,染色体组数与原始细胞相同。 减数分裂包括两个核分裂,它们产生四个遗传上不同的子核,并且在母细胞中有一组染色体,而不是两组染色体。 这些过程之间的主要区别在于减数分裂的第一分裂,在该分裂中,同源染色体成对并交换非姐妹染色体片段。 在减数分裂 I 期间,同源染色体分离成不同的核,导致第一分区的倍体水平降低。 减数分裂的第二个分裂更类似于有丝分裂分裂,不同之处在于子细胞由于交叉而不包含相同的基因组。
脚注
- 1 亚当·威尔金斯和罗宾·霍利迪,“减数分裂从有丝分裂演变而来”,《遗传学 181》(2009):3—12。
- 2 Marilee A. Ramesh、Shehre-Banoo Malik 和 John M. Logsdon,Jr.,“减数分裂基因的系统发育清单:贾第鞭毛虫性行为的证据和减数分裂的早期真核生物起源”,Current Biology 15(2005): 185—91。
词汇表
- chiasmata
- (单数,chiasma)遗传物质交换后在交叉点形成的结构
- cohesin
- 蛋白质形成复合体,将姐妹染色体密封在中心处,直到减数分裂进入第二阶段
- 交叉
- 在非姐妹染色体之间交换遗传物质导致染色体包含来自生物体父母双方的基因
- 施肥
- 来自两个独立生物的两个单倍体细胞的结合
- 运动互动
- (也是 Interphase II)在减数分裂 I 和减数分裂 II 之间短暂休息一段时间
- 减数分裂
- 产生四个单倍体细胞的核分裂过程
- 减数分裂 I
- 第一轮减数分裂细胞分裂;之所以称为还原分裂,是因为倍体等级从二倍体降低到单倍体
- 减数分裂 II
- 减数分裂 I 之后的第二轮减数分裂细胞分裂;姊妹染色体被分离成单个染色体,结果是四个独特的单倍体细胞
- 重组结节
- 在 synaptonemal 复合物上形成的蛋白质组装体标记交叉事件点并介导非姐妹染色体之间遗传重组的多步过程
- 还原部
- 产生子核的核分裂,每个子核的染色体组数是亲核的一半;减数分裂 I 是一个还原分裂
- 体细胞
- 多细胞生物的所有细胞,配子或生殖细胞除外
- 孢子
- 可以产生单倍体多细胞生物或可以与另一个孢子融合形成二倍体细胞的单倍体细胞
- 突触
- 在第一期前期,同源染色体之间形成密切关联
- 突触复合体
- 在第一期前期在同源染色体之间形成的蛋白质晶格,支持交叉
- tetrad
- 两条重复的同源染色体(四条染色体)在第一阶段被 chiasmata 结合在一起