
5.1: 组件和结构
- Page ID
- 202499

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 了解细胞膜的流体镶嵌模型
- 描述膜中磷脂、蛋白质和碳水化合物的功能
- 讨论膜流动性
细胞的质膜定义细胞,勾勒出其边界,并确定其与环境相互作用的性质(摘要见表\(\PageIndex{1}\))。 细胞排出某些物质,吸收其他物质,然后排出其他物质,全部以受控的数量排出。 质膜必须非常灵活,以允许某些细胞(例如红细胞和白细胞)在穿过狭窄的毛细血管时改变形状。 这些是质膜更明显的功能。 此外,质膜表面携带标志物,使细胞能够相互识别,这对于早期发育期间的组织和器官形成至关重要,后来在免疫反应的 “自我” 与 “非自我” 区分中起着作用。
质膜最复杂的功能之一是能够通过称为受体的复杂整体蛋白质传递信号。 这些蛋白质既是细胞外输入的接收者,又是细胞内过程的激活剂。 这些膜受体为激素和生长因子等效应因子提供细胞外附着位点,当它们的效应因子结合时,它们会激活细胞内反应级联。 有时,受体会被病毒(例如HIV,人类免疫缺陷病毒)劫持,这些病毒利用它们进入细胞,有时,编码受体的基因会发生突变,导致信号转导过程出现故障,造成灾难性后果。
流体马赛克模型
质膜的存在是在 19 世纪 90 年代确定的,其化学成分在 1915 年被发现。 当时鉴定的主要成分是脂质和蛋白质。 休·戴夫森和詹姆斯·丹尼利在1935年提出了第一个被广泛接受的质膜结构模型;该模型基于早期电子显微照片中质膜的 “铁路轨道” 外观。 他们认为,质膜的结构类似于三明治,蛋白质类似于面包,脂质类似于馅料。 20 世纪 50 年代,显微镜的进步,尤其是透射电子显微镜(TEM)的进步,使研究人员看到质膜的核心由双层而不是单层组成。 1972 年,S.J. Singer 和 Garth L. Nicolson 提出了一个能够更好地解释微观观测结果和质膜功能的新模型。
辛格和尼科尔森提出的解释被称为流体马赛克模型。 随着时间的推移,该模型有所发展,但仍能最好地解释我们现在所理解的质膜的结构和功能。 流体镶嵌模型将质膜的结构描述为成分的马赛克,包括磷脂、胆固醇、蛋白质和碳水化合物,赋予膜流体特征。 等离子膜的厚度介于 5 到 10 nm 之间。 相比之下,通过光学显微镜可见的人体红细胞宽度约为 8 µm,大约比质膜宽 1000 倍。 膜看起来确实有点像三明治(图\(\PageIndex{1}\))。
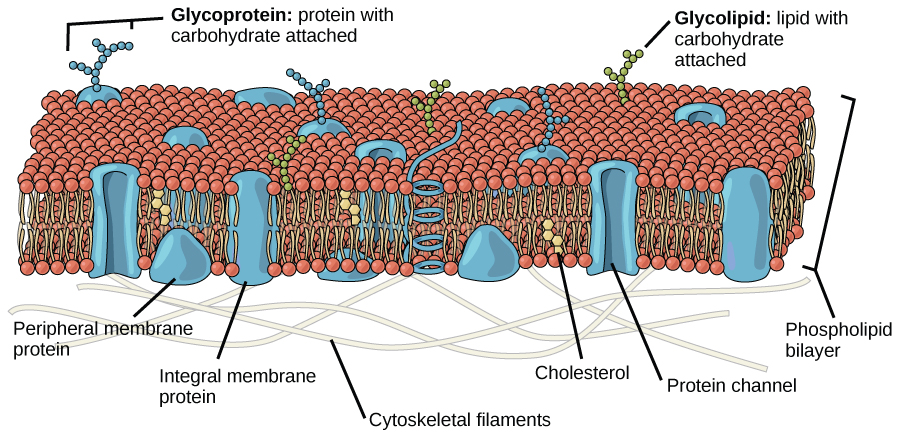
质膜的主要成分是脂质(磷脂和胆固醇)、蛋白质和附着在某些脂质和某些蛋白质上的碳水化合物。 磷脂是由甘油、两种脂肪酸和一个与磷酸盐连接的头组组成的分子。 胆固醇是另一种由四个熔融碳环组成的脂质,与膜核心中的磷脂一起存在。 质膜中蛋白质、脂质和碳水化合物的比例因细胞类型而异,但对于典型的人体细胞,蛋白质按质量计约占成分的50%,脂质(所有类型)按质量计约占成分的40%,其余的10%按质量分列的成分是碳水化合物。 但是,蛋白质和脂质的浓度因细胞膜的不同而不同。 例如,髓磷脂是隔离周围神经轴突的特殊细胞膜的产物,仅含有 18% 的蛋白质和 76% 的脂质。 线粒体内膜含有 76% 的蛋白质和 24% 的脂质。 人类红细胞的质膜为30%的脂质。 碳水化合物仅存在于质膜的外表面,附着在蛋白质上,形成糖蛋白,或附着在脂质上,形成糖脂。
磷脂
膜的主要结构由两亲性磷脂分子组成。 这些分子的亲水或 “爱水” 区域(看起来像艺术家演示的模型中的一组球)(图\(\PageIndex{1}\))与细胞内外的水液接触。 疏水分子或讨厌水分子往往是非极性的。 它们在化学反应中与其他非极性分子相互作用,但通常不与极性分子相互作用。 当放入水中时,疏水分子往往会形成球或团簇。 磷脂的亲水区域往往在细胞的外部和内部与水和其他极性分子形成氢键。 因此,面向细胞内部和外部的膜表面是亲水性的。 相比之下,细胞膜内部具有疏水性,不会与水相互作用。 因此,磷脂形成极好的双层细胞膜,可将细胞内的液体与细胞外的液体分开。
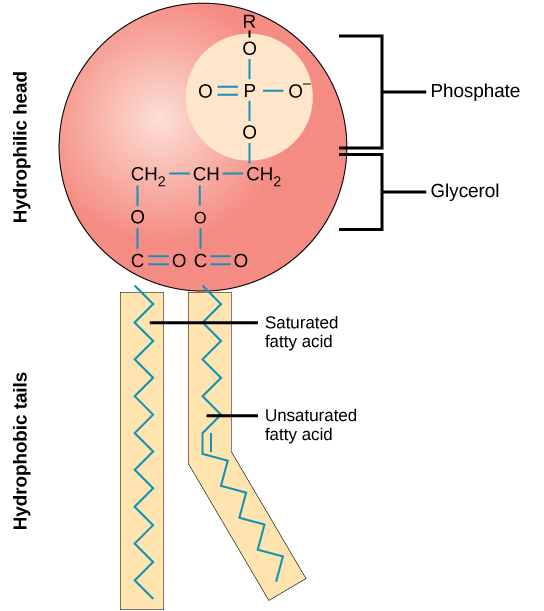
磷脂分子(图\(\PageIndex{2}\))由一个三碳甘油主链组成,其中两个脂肪酸分子附着在碳 1 和 2 上,一个含有磷酸盐的基团附着在第三个碳上。 这种排列使整个分子有一个被描述为其头部的区域(含磷酸盐基团),具有极性特征或负电荷,还有一个称为尾部(脂肪酸)的区域,没有电荷。 头部可以形成氢键,但尾巴不能。 具有正电荷或负电荷区域以及无电荷或非极性区域排列的分子被称为两亲性或 “双重爱好”。
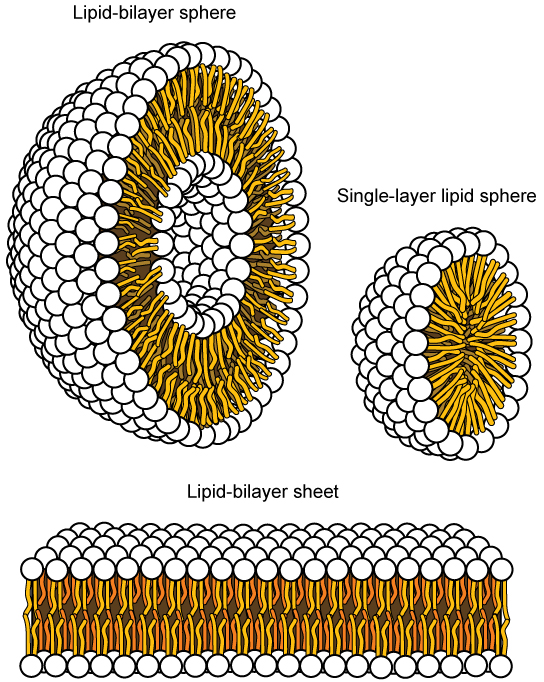
这种特征对质膜的结构至关重要,因为在水中,磷脂往往排列成排列,其疏水性尾巴彼此面对,亲水性头朝外。 通过这种方式,它们形成脂质双层——一种由双层磷脂组成的屏障,将屏障一侧的水和其他物质与另一侧的水和其他物质分开。 实际上,在水溶液中加热的磷脂往往会自发形成小球体或液滴(称为胶束或脂质体),其亲水头形成外部,疏水尾部在内部(图\(\PageIndex{3}\))。
蛋白质
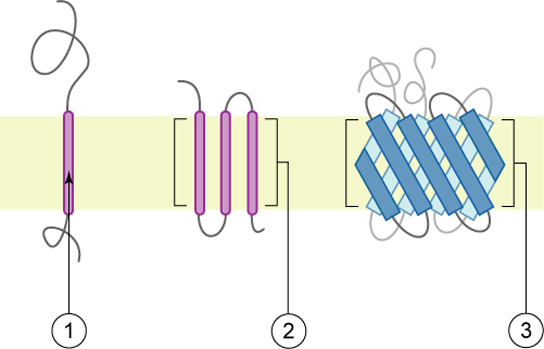
蛋白质构成血浆膜的第二个主要成分。 顾名思义,整体蛋白(一些特殊类型被称为整合素)完全整合到膜结构中,它们的疏水膜跨越区域与磷脂双层的疏水区域相互作用(图\(\PageIndex{1}\))。 单程整合膜蛋白通常具有由20—25个氨基酸组成的疏水性跨膜片段。 有些只跨过膜的一部分(与单层相关),而另一些则从膜的一侧延伸到另一侧,并且两侧都暴露在外。 一些复杂的蛋白质由单个蛋白质的多达12个片段组成,这些片段被广泛折叠并嵌入膜中(图\(\PageIndex{4}\))。 这种类型的蛋白质具有一个或多个亲水区域,以及一个或多个轻度疏水区域。 蛋白质区域的这种排列倾向于使蛋白质与磷脂并列,蛋白质的疏水区域与磷脂的尾部相邻,蛋白质的亲水区域从膜中伸出来并与细胞溶胶接触;或细胞外液体。
外周蛋白存在于膜的外表面和内表面,附着在整体蛋白质或磷脂上。 外周蛋白和整体蛋白可以用作酶,作为细胞骨架纤维的结构附着物,也可以作为细胞识别位点的一部分。 这些有时被称为 “细胞特异性” 蛋白。 人体识别自己的蛋白质并攻击与侵入性病原体相关的外来蛋白。
碳水化
碳水化合物是血浆膜的第三个主要成分。 它们总是存在于细胞的外表面,与蛋白质(形成糖蛋白)或脂质(形成糖脂)结合(图\(\PageIndex{1}\))。 这些碳水化合物链可能由2—60个单糖单位组成,可以是直链的,也可以是分支的。 碳水化合物与外周蛋白一起在细胞表面形成特殊位点,使细胞能够相互识别。 这些部位具有独特的图案,可以识别细胞,就像每个人独特的面部特征允许他或她被识别一样。 这种识别功能对细胞非常重要,因为它允许免疫系统区分人体细胞(称为 “自我”)和外来细胞或组织(称为 “非自我”)。 在病毒表面发现了类似类型的糖蛋白和糖脂,它们可能经常变化,从而阻止免疫细胞识别和攻击它们。
细胞外表面的这些碳水化合物——糖蛋白和糖脂的碳水化合物成分——统称为 glycocalyx(意思是 “糖衣”)。 glycocalyx 具有很高的亲水性,可以将大量的水吸引到细胞表面。 这有助于细胞与其水环境的相互作用,也有助于细胞获得溶解在水中的物质的能力。 如上所述,glycocalyx 对于细胞鉴定、自我/非自主决定和胚胎发育也很重要,用于细胞附着以形成组织。
进化连接:病毒如何感染特定器官
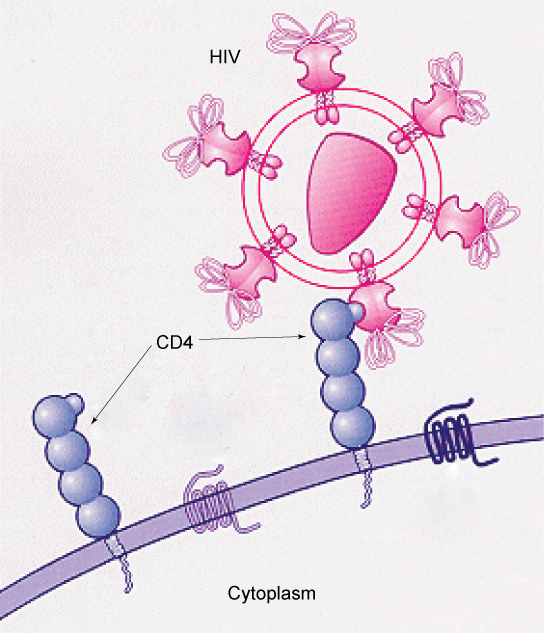
细胞表面的糖蛋白和糖脂模式为许多病毒提供了感染的机会。 HIV和肝炎病毒仅感染人体中的特定器官或细胞。 HIV能够穿透一种称为T辅助细胞的亚型淋巴细胞的质膜,以及一些单核细胞和中枢神经系统细胞。 肝炎病毒攻击肝细胞。
这些病毒能够侵入这些细胞,因为这些细胞的表面有特定于某些病毒并与之相容的结合位点(图\(\PageIndex{5}\))。 病毒表面的其他识别位点与人体免疫系统相互作用,促使人体产生抗体。 抗体是针对与侵袭性病原体相关的抗原或蛋白质而产生的,或者是对外来细胞(例如器官移植可能发生的外来细胞)的反应。 这些相同的位点是抗体附着的地方,可以破坏或抑制病毒的活性。 不幸的是,由于突变,HIV上的这些识别位点变化很快,这使得随着病毒的进化和适应,生产针对该病毒的有效疫苗变得非常困难。 感染HIV的人将很快发展出不同的病毒群体或变异体,这些群体或变异体以这些识别位点的差异为特征。 表面标志物的这种快速变化降低了人体免疫系统攻击病毒的有效性,因为抗体无法识别表面模式的新变异。 就HIV而言,该病毒特异性感染和破坏参与免疫反应的细胞,使宿主进一步丧失能力,这一事实使问题变得更加复杂。
膜流动性
流体马赛克模型中描述的膜的马赛克特性有助于说明其性质。 不可分割的蛋白质和脂质作为单独但附着松散的分子存在于膜中。 它们类似于马赛克图片中单独的彩色瓷砖,它们漂浮,彼此之间有些移动。 但是,膜不像可以膨胀和收缩的气球;相反,它相当坚硬,如果被穿透或者细胞吸收过多的水,可能会破裂。 但是,由于其镶嵌性质,非常细的针头可以很容易地穿透质膜而不会导致其破裂,并且在提取针头时膜会流动并自密封。
膜的马赛克特性解释了其流动性的部分但不是全部。 还有其他两个因素有助于保持这种流体特性。 一个因素是磷脂本身的性质。 在饱和形式中,磷脂尾中的脂肪酸被结合的氢原子饱和。 相邻的碳原子之间没有双键。 这会导致尾巴相对较直。 相比之下,不饱和脂肪酸不含最大数量的氢原子,但它们在相邻的碳原子之间确实含有一些双键;双键会导致碳串弯曲约30度(图\(\PageIndex{2}\))。
因此,如果饱和脂肪酸及其直尾部因温度下降而被压缩,它们就会相互压缩,形成致密且相当坚硬的膜。 如果不饱和脂肪酸被压缩,它们尾巴上的 “扭结” 会使相邻的磷脂分子消失,从而在磷脂分子之间保持一定的空间。 这种 “肘部空间” 有助于保持膜的流动性,在此温度下,磷脂中含有饱和脂肪酸尾部的膜会 “冻结” 或固化。 在寒冷的环境中,膜的相对流动性尤其重要。 寒冷的环境往往会压缩主要由饱和脂肪酸组成的膜,使它们的液体减少并且更容易破裂。 许多生物(例如鱼)能够通过改变其膜中不饱和脂肪酸的比例来适应寒冷的环境,以应对温度的降低。
链接到学习
访问此网站,观看膜的流动性和镶嵌质量的动画。
动物有额外的膜成分,有助于保持流动性。 胆固醇与膜中的磷脂并存,往往会抑制温度对膜的影响。 因此,这种脂质起到缓冲作用,防止较低的温度抑制流动性,并防止温度升高导致流动性过度增加。 因此,胆固醇在两个方向上延伸了膜具有适当流体从而起作用的温度范围。 胆固醇还具有其他功能,例如将跨膜蛋白簇组织成脂质筏。
| 组件 | 地点 |
|---|---|
| 磷脂 | 薄膜的主要面料 |
| 胆固醇 | 附着在磷脂之间和两个磷脂层之间 |
| 整体蛋白(例如,整合素) | 嵌入在磷脂层内。 可能会也可能不会穿透两层 |
| 外周蛋白 | 在磷脂双层的内表面或外表面;未嵌入磷脂中 |
| 碳水化合物(糖蛋白和糖脂的成分) | 通常附着在外膜层的蛋白质上 |
职业联系:免疫学家
影响细胞识别位点的外周蛋白和碳水化合物的变异是免疫学中最感兴趣的。 在疫苗开发中考虑到了这些变化。 疫苗的使用战胜了许多传染病,例如天花、脊髓灰质炎、白喉和破伤风。
免疫学家是研究和开发疫苗以及治疗和研究过敏或其他免疫问题的医生和科学家。 一些免疫学家研究和治疗自身免疫问题(一个人的免疫系统攻击他或她自己的细胞或组织的疾病,例如狼疮)和免疫缺陷,无论是获得性免疫缺陷(例如获得性免疫缺陷综合征或艾滋病)还是遗传性的(例如严重的联合免疫缺陷或 SCID))。 免疫学家被要求帮助治疗器官移植患者,这些患者的免疫系统必须受到抑制,这样他们的身体就不会排斥移植的器官。 一些免疫学家努力了解自然免疫以及人的环境对自然免疫的影响。 其他人则在研究有关免疫系统如何影响癌症等疾病的问题。 过去,人们根本不了解拥有健康的免疫系统在预防癌症方面的重要性。
要成为免疫学家,需要博士或医学博士。 此外,免疫学家在认可的课程中接受至少2-3年的培训,并且必须通过美国过敏和免疫学委员会的考试。 免疫学家必须了解与免疫以外问题有关的人体功能,以及药理学和医疗技术(例如药物、疗法、测试材料和外科手术)的知识。
摘要
现代人对质膜的理解被称为流体镶嵌模型。 质膜由双层磷脂组成,它们的疏水性脂肪酸尾部相互接触。 膜的景观布满了蛋白质,其中一些蛋白质横跨膜。 其中一些蛋白质用于将物质运入或运出细胞。 碳水化合物附着在膜朝外表面的某些蛋白质和脂质上,形成复合物,可以将细胞识别给其他细胞。 膜的流体性质归因于温度、脂肪酸尾部的构造(有些被双键扭结)、膜中嵌入胆固醇的存在以及蛋白质和蛋白质-碳水化合物组合的镶嵌性质,这些组合物没有牢固地固定在适当的位置。 质膜封闭并定义了细胞的边界,但它们不是一个静态的袋子,而是动态的,不断变化。
词汇表
- 两亲性的
- 具有极性或带电区域以及能够与亲水和疏水环境相互作用的非极性或无电荷区域的分子
- 流体马赛克模型
- 将质膜的结构描述为包括磷脂、胆固醇、蛋白质、糖蛋白和糖脂(分别附着在蛋白质或脂质上的糖链)等成分的马赛克,从而形成流体特征(流动性)
- 糖脂
- 碳水化合物和脂质的组合
- 糖蛋白
- 碳水化合物和蛋白质的组合
- 亲水性的
- 具有与水结合能力的分子;“爱水”
- 恐水的
- 没有能力与水结合的分子;“讨厌水”
- 整体蛋白
- 蛋白质整合到膜结构中,与膜脂的碳氢化合物链广泛相互作用,通常横跨膜;这些蛋白质只有通过洗涤剂破坏膜才能去除
- 外周蛋白
- 蛋白质存在于质膜表面,无论是外侧还是内侧;这些蛋白质可以通过高盐洗涤去除(从膜上冲走)