5.2: 定义 Homo 属
- Page ID
- 204752

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)在本节结束时,您将能够:
将 Homo 置于情境中
在了解构成 H omo 属的人类物种之前,熟悉一下与 H omo 相关的关键考古时期会很有帮助。 以下各节将更详细地探讨下文提到的物种和文化发展。
定义 Homo 属的挑战
上一章介绍了南方古猿,它们的身体特征各不相同(柔软而健壮),下巴和牙齿大,大脑大小小。 南方古猿和人属的一个关键特征是两足动物。 向两足动物的过渡与各种解剖学变化有关,包括腿部更长、脊柱曲率变化以及足弓的发育以节省能量并增加行走时的平衡。
除了两足动物之外,还有什么标准可以用来将物种归类为 Homo 属? 许多人类学家试图建立特定的标准,用于确定 Homo的分类。 古人类学家玛丽·利基、路易斯·利基和约翰·纳皮尔以及灵长类动物学家菲利普·托比亚斯是最早广泛研究哈比利斯人化石的人之一,哈比利斯被认为是人类属中最早的物种之一。 H.habil is的大脑大小约为661—700 cc,比南方古猿大,由于骨骼结构的变化和拇指的重新定位,双手能够灵活地制作工具,这使得握力更好。
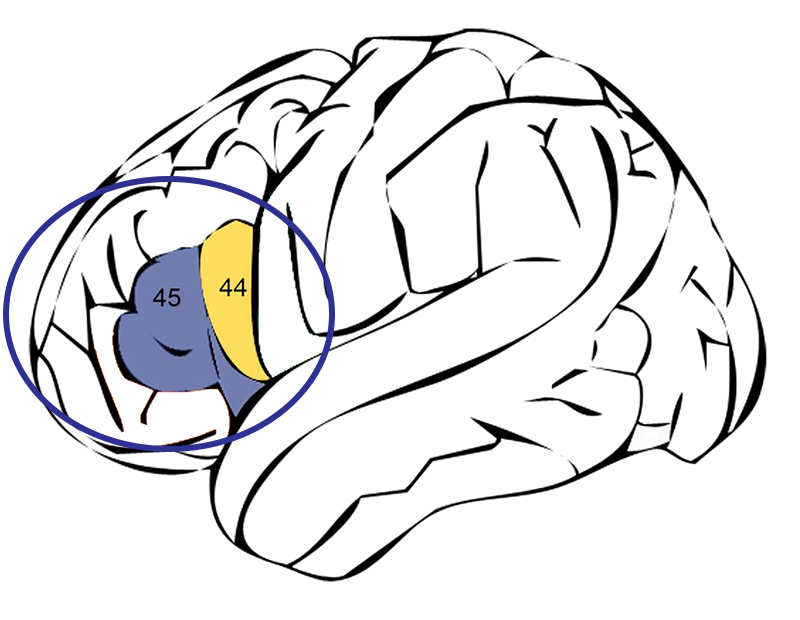
H.habilis 的 OH 7 型标本可追溯到 2 至 1.7 MYA 之间,由乔纳森和玛丽·利基于 1960 年在奥尔杜瓦伊峡谷发现。 它是路易斯·利基在 1964 年描述的。 类型标本是指作为该物种分类单元或分类组标准的标本。 OH 7 是这个特定标本的识别号或入藏号,代表 “Olduvai Hominid #7”。 标本由部分幼体头骨、手骨和脚骨组成。 它的牙齿比任何南方古猿都要小得多,并且可能与强大的南方古猿(Paranthropus)共存。 根据颅内石膏(脑壳内部的印记),确定哈比利斯可能在大脑中拥有所谓的布罗卡区域。 布罗卡的区域包括两个布罗德曼区域(称为44和45),位于大脑左大脑皮层的中间,对言语发育尤其重要(图5.2)。 一些科学家提出,哈比利斯开始开发人类言语所必需的神经网络,而另一些科学家则认为哈比利斯可能已经有言语了。
Homo habil is的颅后特征(除头骨以外的身体骨骼结构)并不像许多其他早期的人类化石那样得到充分证实。 这可能是有问题的,因为许多人参物种与重叠的特征共存。 同样,有颅后物质而不是颅骨或头骨也可能有问题。 当颅后材料不能提供足够的证据或提供令人困惑的证据时,头骨通常用作诊断工具。
根据他们对 H. habilis 的研究,Mary Leakey、Louis Leakey 和 John Napier 提出了以下对 Homo 进行分类的标准:大脑大小超过 600 cc;圆形球状头骨;工具使用;减少前后倾力(下巴突出)以及下巴和下颌骨变小;像人一样颅后特征;以及完全适合行走的双脚(Leakey、Tobias 和 Napier 1964)。 尽管这份清单确立了具体且相当全面的指导,但特征的多样性及其重叠方式并不总是符合标准。
哈比利斯一直是关于其分类学地位以及与其他早期古人类物种关系的几场辩论的焦点。 例如,哈比利斯最初被认为是直立人和现代人类血统中的直接人类祖先。 这种观点现在正在争论中,导致了支持哈比利斯的人和那些认为另一种人类物种 H. rudolfensis 是直立杆菌祖先的人之间存在科学分歧。 H. rudolfensis 是一种古老的 Homo,其历史可追溯到大约 2 MYA,在此期间与其他人类物种共存。 1972 年,当地肯尼亚人伯纳德·恩吉内奥在肯尼亚的图尔卡纳湖沿岸发现了颅骨。 古人类学家理查德·利基后来描述了这个标本。 关于这个物种还有很多未知的地方;科学家们缺少颅后材料,到目前为止还没有发现任何工具。 有人假设 H. rudolfensis 可能是哈比利斯雄性,其颅骨比雌性 H. habilis 的颅骨大。 其他人则认为它是一个完全不同的物种。 另一个争议集中在工具的使用上。 尽管哈比利斯人长期以来一直被认为是最早使用石器的人类,但根据切割痕迹的证据,已经确定至少有一个南方古猿(A. garhi)在哈比利斯之前使用石器,大约为 2.6 MYA(Semaw等人,1997年)。

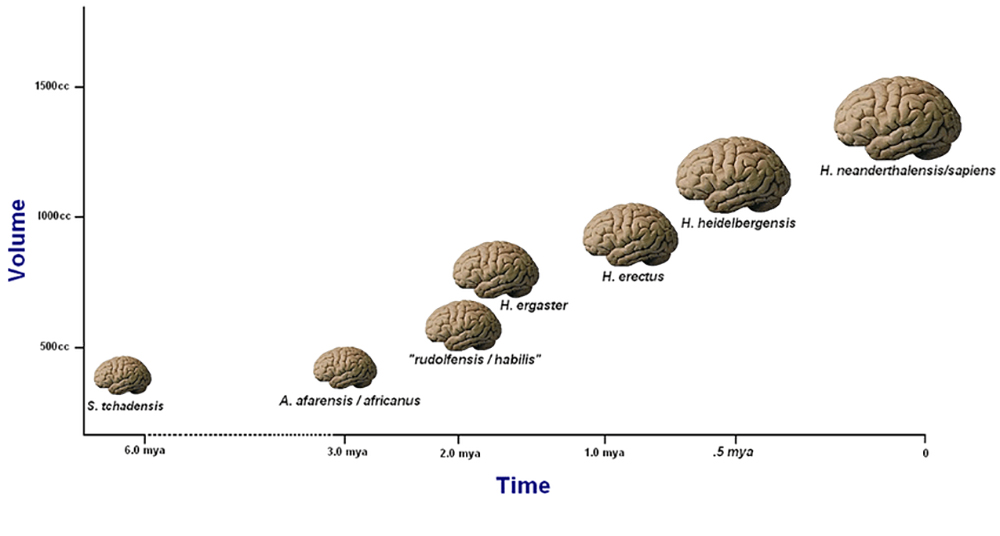
将 H. habilis 归类为人而不是南方古猿的主要考虑因素之一是其颅骨容量,这是一种表明大脑大小的测量值。 除了一些例外,颅骨容量可以作为人参进化树中人参化石可能属于哪里的指标。 脑化是指随着时间的推移,大脑大小逐渐增加。 在人类进化过程中,我们可以观察到脑化从 H omo habilis 开始,然后通过直立杆菌发展得更快。 脑化与行为、认知和文化复杂性的增加相关。 认知发展与我们构造和形成想法的能力相对应,包括通过象征性和抽象语言(例如讲故事、仪式和艺术中使用的语言)进行思考和沟通的能力。 但是,总会有例外,例如居住在岛上、脑子矮小的 H. floresiensis,他将在本章后面介绍。 尽管大脑很小,但 H. floresiensis 还是制造和使用了工具并制造了火灾。 这一发现挑战了我们认为我们对人类进化中大脑大小与认知发育之间相关性的认识。
脑化商(EQ)可以作为H omo 属内分类的良好指标(某些例外情况除外)。 脑化商数是通过比较实际大脑大小(通过质量或体积计算确定)与预期大脑大小之间的比率得出的计算结果。 体型是这些测量中的一个因素,因为预期的大脑大小反映了给定分类组的大脑和体型之间的关系(Jerison 1973)。 相对于总体体重,大脑的重量越大,大脑被用于更复杂的认知任务的可能性就越大。 哈里·杰里森(1973 年)是第一个开发均衡器测量的人。 他用于计算鸟类和哺乳动物情商的公式是脑质量/0.12×(体重)0.66。 还提出了其他公式,例如情商=大脑质量(11.22×体重0.76)(Martin 1981)。 尽管情商是研究早期人参素大脑大小的有力工具,但在处理碎片化石时总会有潜在的误差余地,并且正在提出越来越多的替代测量形式。 一项研究提出,不应再使用情商作为计算灵长类动物和其他脊椎动物大脑大小的工具,其前提是认知能力不取决于体型,因此不应将体型包括在公式中(Schaik 等人,2021)。 其他理论认为,在考虑认知能力时,皮质神经元和神经连接的数量最为重要(Roth and Dicke 2012)。 根据这种方法,皮层的密度更多地与智力有关,而不是与大脑的大小有关。 这些替代方法也许可以更好地解释化石记录中的例外情况,例如 H. floresiensis。 其他有趣的研究是研究与工具复杂程度相关的潜在认知和记忆水平(Read and van der Leeuw 2008)。
尽管有这些批评,但许多人认为情商测量提供了相当一致的结果。 现代人类(智人)的情商约为6.0—7.0(这意味着他们的大脑质量是人们在相同体型的可比哺乳动物中预期的六到七倍)。 直立人的情商为4.0,而南方古猿的情商约为2.5至3.0(Fuente 2012,227)。 图 5.4 显示了不同物种的平均大脑大小随着时间的推移而增加。