
11.2: تكرار الحمض النووي
- Page ID
- 194674

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)أهداف التعلم
- شرح معنى تكرار الحمض النووي شبه المحافظ
- اشرح لماذا يكون تكرار الحمض النووي ثنائي الاتجاه ويتضمن خيطًا رائدًا ومتخلفًا
- اشرح لماذا تتشكل شظايا أوكازاكي
- وصف عملية تكرار الحمض النووي ووظائف الإنزيمات المعنية
- حدد الاختلافات بين تكرار الحمض النووي في البكتيريا وحقيقيات النواة
- شرح عملية النسخ المتماثل للدائرة الدوارة
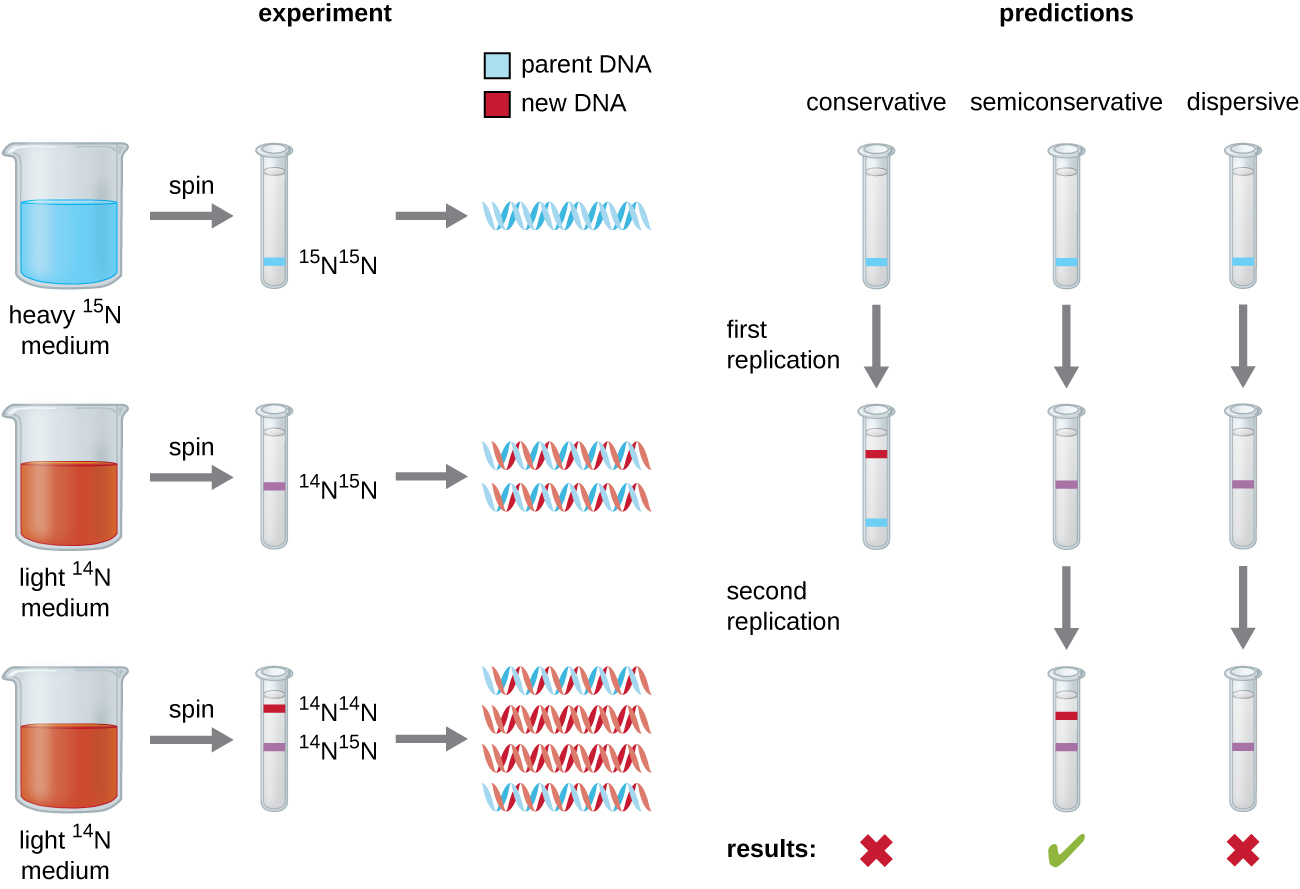
قدم توضيح بنية اللولب المزدوج من قبل جيمس واتسون وفرانسيس كريك في عام 1953 تلميحًا عن كيفية نسخ الحمض النووي أثناء عملية النسخ المتماثل. سيوفر فصل خيوط اللولب المزدوج نموذجين لتوليف خيوط تكميلية جديدة، ولكن بالضبط كيف تم بناء جزيئات الحمض النووي الجديدة لا تزال غير واضحة. في أحد النماذج، وهو النسخ المتماثل شبه المحافظ، ينفصل شريطان اللولب المزدوج أثناء تكرار الحمض النووي، ويعمل كل خيط كقالب يتم نسخ الخيط التكميلي الجديد منه؛ وبعد النسخ المتماثل، يتضمن كل حمض نووي مزدوج الخصلة خيطًا أبويًا أو «قديمًا» وخيطًا «جديدًا» واحدًا. تم اقتراح نموذجين متنافسين أيضًا: نموذج متحفظ ونموذج مشتت، كما هو موضح في الشكل\(\PageIndex{1}\).

ابتكر ماثيو ميسيلسون (1930—) وفرانكلين ستال (1929—) تجربة في عام 1958 لاختبار أي من هذه النماذج يمثل بشكل صحيح تكرار الحمض النووي (الشكل\(\PageIndex{2}\)). لقد قاموا بزراعة E. coli لعدة أجيال في وسط يحتوي على نظير «ثقيل» للنيتروجين (15 نيوتن) تم دمجه في القواعد النيتروجينية، وفي النهاية، في الحمض النووي. هذا يسمى الحمض النووي الأبوي. ثم تم تحويل ثقافة E. coli إلى وسط يحتوي على 14 نيوتن وسمح لها بالنمو لجيل واحد. تم حصاد الخلايا وعزل الحمض النووي. تم فصل الحمض النووي عن طريق الطرد المركزي الفائق، والذي شكل خلاله الحمض النووي نطاقات وفقًا لكثافته. من المتوقع أن يشكل الحمض النووي الذي ينمو في 15 نيوتن نطاقًا في موضع كثافة أعلى من ذلك الذي نما في 14 نيوتن. وأشار ميسيلسون وستال إلى أنه بعد جيل واحد من النمو في 14 نيوتن، كان النطاق الواحد الذي لوحظ متوسطًا في الموضع بين الحمض النووي للخلايا التي نمت حصريًا في 15 N أو 14 N. اقترح هذا إما أسلوب التكرار شبه المحافظ أو التشتت. تم السماح لبعض الخلايا بالنمو لجيل آخر في 14 نيوتن ثم نسجها مرة أخرى. شكل الحمض النووي الذي تم حصاده من الخلايا التي نمت لجيلين في 14 نيوتن نطاقين: كان أحد نطاقات الحمض النووي في الموضع المتوسط بين 15 N و 14 N، والآخر يتوافق مع نطاق 14 N DNA. لا يمكن تفسير هذه النتائج إلا إذا تم تكرار الحمض النووي بطريقة شبه محافظة. لذلك، تم استبعاد النموذجين الآخرين. نتيجة لهذه التجربة، نعلم الآن أنه أثناء تكرار الحمض النووي، يعمل كل من الخطين اللذين يشكلان اللولب المزدوج كقالب يتم نسخ خيوط جديدة منه. سيكون الشريط الجديد مكملاً للحبل الأبوي أو «القديم». تحتوي جزيئات الحمض النووي الناتجة على نفس التسلسل وتنقسم بالتساوي إلى الخليتين البنتين.
التمارين\(\PageIndex{1}\)
ماذا كان يمكن أن تكون نتيجة تجربة ميسيلسون وستال إذا عثروا، بعد الجيل الأول، على شريطين من الحمض النووي؟
تكرار الحمض النووي في البكتيريا
تمت دراسة تكرار الحمض النووي جيدًا في البكتيريا في المقام الأول بسبب صغر حجم الجينوم والطفرات المتوفرة. يحتوي E. coli على 4.6 مليون زوج أساسي (Mbp) في كروموسوم دائري واحد ويتم تكراره جميعًا في حوالي 42 دقيقة، بدءًا من أصل واحد للتكاثر والمضي قدمًا حول الدائرة ثنائية الاتجاه (أي في كلا الاتجاهين). هذا يعني أنه يتم إضافة ما يقرب من 1000 نيوكليوتيدات في الثانية. هذه العملية سريعة جدًا وتحدث مع بعض الأخطاء.
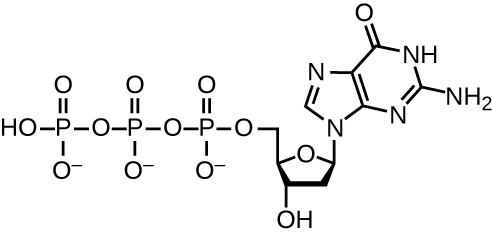
يستخدم تكرار الحمض النووي عددًا كبيرًا من البروتينات والإنزيمات (الجدول\(\PageIndex{1}\)). أحد اللاعبين الرئيسيين هو إنزيم بوليميراز DNA، المعروف أيضًا باسم DNA pol. في البكتيريا، تُعرف ثلاثة أنواع رئيسية من بوليميرات الحمض النووي: DNA pol I و DNA pol II و DNA pol III. من المعروف الآن أن DNA pol III هو الإنزيم المطلوب لتخليق الحمض النووي؛ DNA pol I و DNA pol II مطلوبان بشكل أساسي للإصلاح. يضيف DNA pol III الديوكسي ريبونوكليوتيدات كل منها مكملاً للنيوكليوتيد على شريط القالب، واحدًا تلو الآخر إلى مجموعة 3'-OH من سلسلة الحمض النووي المتنامية. تتطلب إضافة هذه النيوكليوتيدات طاقة. توجد هذه الطاقة في روابط ثلاث مجموعات فوسفات مرتبطة بكل نيوكليوتيد (نيوكليوتيد ثلاثي الفوسفات)، على غرار كيفية تخزين الطاقة في روابط الفوسفات الخاصة بالأدينوزين ثلاثي الفوسفات (ATP) (الشكل\(\PageIndex{3}\)). عندما تنكسر الرابطة بين الفوسفات ويتم إطلاق ثنائي الفوسفات، تسمح الطاقة المنبعثة بتكوين رابطة فوسفوديستر تساهمية عن طريق تخليق الجفاف بين النيوكليوتيد الوارد ومجموعة 3'-OH الحرة على خيط الحمض النووي المتنامي.
المبادرة
يحدث بدء النسخ المتماثل عند تسلسل نوكليوتيد محدد يسمى أصل النسخ المتماثل، حيث ترتبط البروتينات المختلفة لبدء عملية النسخ المتماثل. يحتوي E. coli على أصل وحيد للتكاثر (كما هو الحال مع معظم بدائيات النواة)، يسمى ORIC، على كروموسوم واحد. يبلغ أصل النسخ المتماثل ما يقرب من 245 زوجًا أساسيًا وهو غني بتسلسلات الأدينين-الثيامين (AT).
تعتبر بعض البروتينات التي ترتبط بأصل النسخ المتماثل مهمة في جعل المناطق أحادية الشريط من الحمض النووي متاحة للتكرار. عادةً ما يتم لف الحمض النووي الكروموسومي حول الأحجار الهستونية (في حقيقيات النواة والأركيا) أو البروتينات الشبيهة بالهيستون (في البكتيريا)، ويتم لفه بشكل فائق أو لفه على نطاق واسع ولفه على نفسه. هذه العبوة تجعل المعلومات الموجودة في جزيء الحمض النووي غير قابلة للوصول. ومع ذلك، فإن الإنزيمات التي تسمى توبويزوميراز تغير شكل الكروموسوم ولفه الفائق. لبدء تكرار الحمض النووي البكتيري، يتم تخفيف الكروموسوم فائق اللف بواسطة توبويزوميراز II، المعروف أيضًا باسم DNA gyrase. ثم يفصل إنزيم يسمى الهليكوز خيوط الحمض النووي عن طريق كسر الروابط الهيدروجينية بين أزواج القاعدة النيتروجينية. تذكر أن تسلسلات AT تحتوي على روابط هيدروجينية أقل، وبالتالي فهي تحتوي على تفاعلات أضعف من تسلسلات الغوانين-السيتوزين (GC). تتطلب هذه الإنزيمات التحلل المائي ATP. عندما ينفتح الحمض النووي، تتشكل هياكل على شكل حرف Y تسمى شوكات النسخ المتماثل. يتم تشكيل شوكتين للتكرار في أصل النسخ المتماثل، مما يسمح بالتكرار ثنائي الاتجاه وتشكيل بنية تشبه الفقاعة عند عرضها باستخدام المجهر الإلكتروني للإرسال؛ ونتيجة لذلك، تسمى هذه البنية فقاعة النسخ المتماثل. يتم طلاء الحمض النووي بالقرب من كل شوكة نسخ ببروتينات ربط أحادية الشريط لمنع الحمض النووي أحادي الجديلة من الارتداد إلى حلزون مزدوج.
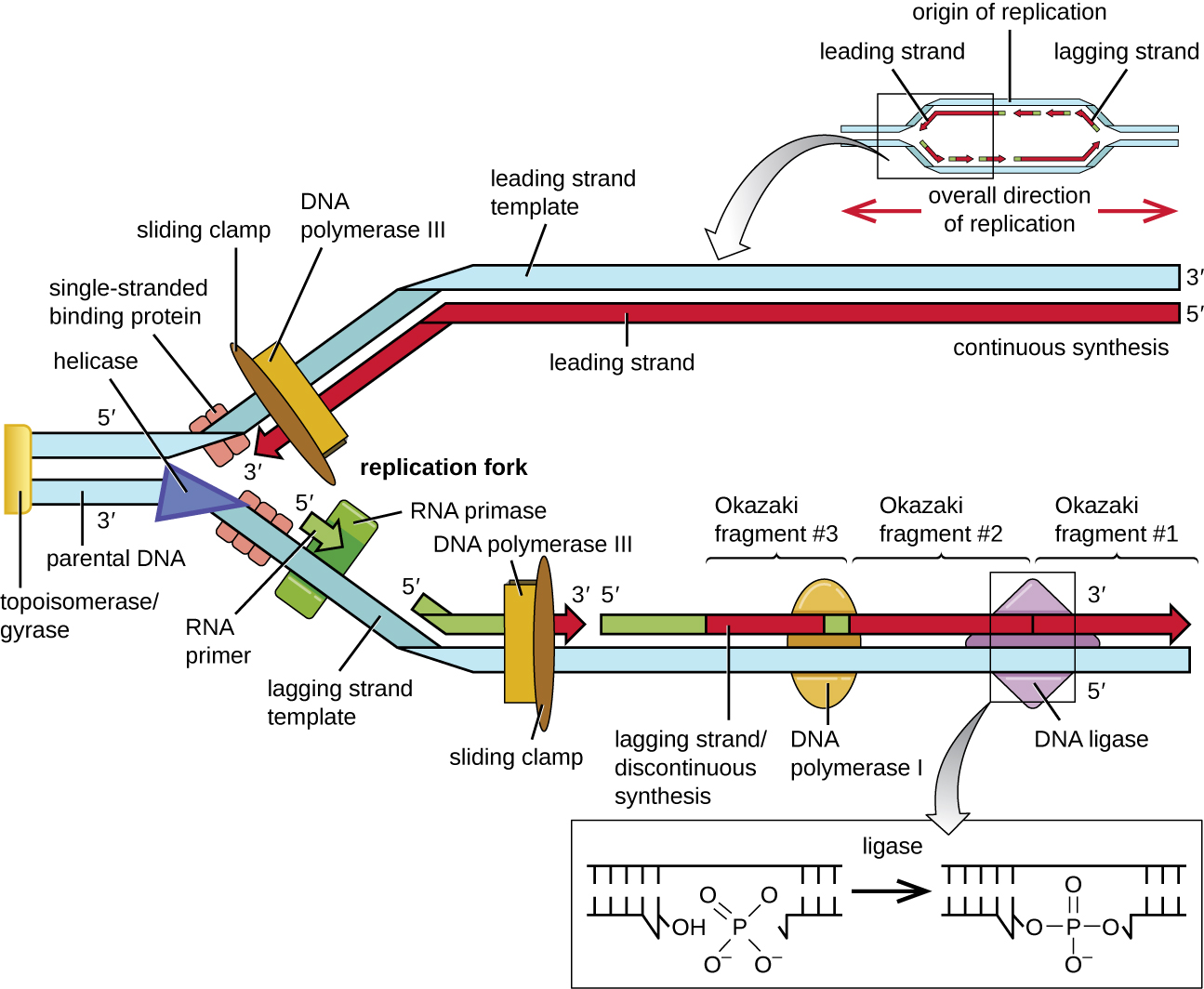
بمجرد الوصول إلى الحمض النووي أحادي الجديلة في أصل النسخ المتماثل، يمكن أن يبدأ تكرار الحمض النووي. ومع ذلك، فإن DNA pol III قادر على إضافة النيوكليوتيدات فقط في اتجاه 5 إلى 3 بوصات (يمكن تمديد خيط DNA جديد فقط في هذا الاتجاه). هذا لأن بوليميراز الحمض النووي يتطلب مجموعة 3'-OH مجانية يمكنه إضافة النيوكليوتيدات إليها عن طريق تكوين رابطة فوسفوديستر تساهمية بين نهاية 3'-OH وفوسفات 5' من النيوكليوتيد التالي. هذا يعني أيضًا أنه لا يمكن إضافة النيوكليوتيدات في حالة عدم توفر مجموعة 3'-OH المجانية، وهذا هو الحال بالنسبة لخيط واحد من الحمض النووي. تم حل المشكلة بمساعدة تسلسل RNA الذي يوفر نهاية 3'-OH المجانية. نظرًا لأن هذا التسلسل يسمح ببدء تخليق الحمض النووي، فإنه يُطلق عليه بشكل مناسب اسم البرايمر. يتكون البرايمر من خمسة إلى 10 نيوكليوتيدات وهو مكمل للحمض النووي الأبوي أو النموذجي. يتم تصنيعه بواسطة RNA primase، وهو بوليميراز RNA. على عكس بوليميرات الحمض النووي، لا تحتاج بوليميرات الحمض النووي الريبي إلى مجموعة 3'-OH مجانية لتجميع جزيء RNA. الآن بعد أن يوفر البرايمر مجموعة 3'-OH المجانية، يمكن الآن لبوليميراز الحمض النووي الثالث توسيع نطاق هذا الحمض النووي التمهيدي، بإضافة نيوكليوتيدات الحمض النووي واحدة تلو الأخرى التي تكمل خيط القالب (الشكل\(\PageIndex{1}\)).
الاستطالة
أثناء الاستطالة في تكرار الحمض النووي، تحدث إضافة النيوكليوتيدات بمعدل أقصى يبلغ حوالي 1000 نيوكليوتيدات في الثانية. يمكن أن يمتد بوليميراز الحمض النووي III فقط في اتجاه 5 إلى 3 بوصات، مما يشكل مشكلة في شوكة النسخ المتماثل. الحلزون المزدوج للحمض النووي مضاد للتوازي؛ أي أن أحد الخطين موجه في الاتجاه من 5 إلى 3 بوصات والآخر موجه في اتجاه 3 إلى 5 بوصات (انظر بنية ووظيفة الحمض النووي). أثناء النسخ المتماثل، يتم تصنيع خيط واحد، وهو مكمل لخيط الحمض النووي الأبوي من 3 إلى 5 بوصات، بشكل مستمر باتجاه شوكة النسخ المتماثل لأن البوليميراز يمكن أن يضيف النيوكليوتيدات في هذا الاتجاه. يُعرف هذا الشريط المركب باستمرار باسم الخصلة الرائدة. أما الخيط الآخر، المكمل للحمض النووي الأبوي الذي يتراوح طوله بين 5 بوصات و3 بوصات، فينمو بعيدًا عن شوكة النسخ المتماثل، لذلك يجب أن يتحرك البوليميراز مرة أخرى نحو شوكة النسخ المتماثل لبدء إضافة قواعد إلى برايمر جديد، مرة أخرى في الاتجاه بعيدًا عن شوكة النسخ المتماثل. يقوم بذلك حتى يصطدم بالحبل الذي تم تصنيعه مسبقًا ثم يتحرك مرة أخرى (الشكل\(\PageIndex{4}\)). تنتج هذه الخطوات أجزاء صغيرة من تسلسل الحمض النووي تُعرف باسم أجزاء Okazaki، يتم فصل كل منها بواسطة RNA primer. تمت تسمية أجزاء أوكازاكي على اسم فريق البحث الياباني والزوجان ريجي وتسونيكو أوكازاكي، الذين اكتشفوها لأول مرة في عام 1966. يُعرف الخيط الذي يحتوي على شظايا أوكازاكي باسم الخصلة المتخلفة، ويقال إن تركيبه متقطع.
يمكن تمديد الخصلة الرائدة من برايمر واحد فقط، بينما تحتاج الخصلة المتخلفة إلى برايمر جديد لكل قطعة من أجزاء أوكازاكي القصيرة. سيكون الاتجاه العام للحبل المتخلف من 3 إلى 5 بوصات، والاتجاه العام للشريط الرائد من 5 إلى 3 بوصات. يقوم بروتين يسمى المشبك المنزلق بتثبيت بوليميراز الحمض النووي في مكانه حيث يستمر في إضافة النيوكليوتيدات. المشبك المنزلق عبارة عن بروتين على شكل حلقة يرتبط بالحمض النووي ويحافظ على البوليميراز في مكانه. بالإضافة إلى دوره في البدء، يمنع التوبويزوميراز أيضًا لف اللولب المزدوج للحمض النووي قبل شوكة النسخ المتماثل أثناء فتح الحمض النووي؛ وهو يفعل ذلك عن طريق إحداث شقوق مؤقتة في حلزون الحمض النووي ثم إعادة غلقه. مع تقدم عملية التوليف، يتم استبدال بادئات الحمض النووي الريبي بالحمض النووي. تتم إزالة المواد الأولية من خلال نشاط إكسونوكلياز لبوليميراز الحمض النووي I، ويتم ملء الفجوات. يتم ختم النكات المتبقية بين الحمض النووي المركب حديثًا (الذي حل محل الحمض النووي الريبي التمهيدي) والحمض النووي المركب مسبقًا بواسطة إنزيم ليغاز الحمض النووي الذي يحفز تكوين ارتباط الفسفوديستر التساهمي بين نهاية 3'-OH لجزء واحد من الحمض النووي ونهاية الفوسفات 5' للجزء الآخر، تثبيت العمود الفقري لفوسفات السكر لجزيء الحمض النووي.
الإنهاء
بمجرد تكرار الكروموسوم الكامل، يجب إنهاء تكرار الحمض النووي. على الرغم من معرفة الكثير عن بدء النسخ المتماثل، إلا أنه لا يُعرف الكثير عن عملية الإنهاء. بعد النسخ المتماثل، يتم ربط الجينومات الدائرية الكاملة الناتجة عن بدائيات النواة، مما يعني أن كروموسومات الحمض النووي الدائرية متشابكة ويجب فصلها عن بعضها البعض. يتم تحقيق ذلك من خلال نشاط توبويزوميراز IV البكتيري، الذي يدخل فواصل مزدوجة في جزيئات الحمض النووي، مما يسمح لها بالانفصال عن بعضها البعض؛ ثم يعيد الإنزيم إغلاق الكروموسومات الدائرية. يعد حل المحاريات مشكلة فريدة لتكرار الحمض النووي الأولي بسبب الكروموسومات الدائرية. نظرًا لاختلاف كل من الحمض النووي الجرثومي gyrase و topoisomerase IV عن نظرائهما حقيقية النواة، تعمل هذه الإنزيمات كأهداف لفئة من الأدوية المضادة للميكروبات تسمى الكينولونات.
| إنزيم أو عامل | وظيفة |
|---|---|
| عينة الحمض النووي الأولى | يزيل نشاط إكسونوكلياز برايمر RNA ويستبدله بالحمض النووي المركب حديثًا |
| مجموعة الحمض النووي 3 | الإنزيم الرئيسي الذي يضيف النيوكليوتيدات في اتجاه 5 إلى 3 |
| هيليكاس | يفتح حلزون الحمض النووي عن طريق كسر الروابط الهيدروجينية بين القواعد النيتروجينية |
| الليغاس | يسد الفجوات بين شظايا أوكازاكي على الخيط المتخلف لإنشاء خيط واحد مستمر من الحمض النووي |
| زهرة الربيع | يقوم بتجميع بادئات الحمض النووي الريبي اللازمة لبدء النسخ المتماثل |
| بروتينات ربط أحادية الجديلة | اربط بالحمض النووي أحادي الجديلة لمنع الترابط الهيدروجيني بين خيوط الحمض النووي، وإصلاح الحمض النووي المزدوج |
| مشبك منزلق | يساعد على تثبيت الحمض النووي POL III في مكانه عند إضافة النيوكليوتيدات |
| توبويزوميراز الثاني (جيراز الحمض النووي) | يريح الكروموسوم فائق اللف لجعل الحمض النووي أكثر سهولة لبدء النسخ؛ يساعد على تخفيف الضغط على الحمض النووي عند الفك، عن طريق التسبب في حدوث فواصل ثم إعادة إغلاق الحمض النووي |
| توبويزوميراز الرابع | يقوم بإدخال كسر أحادي الجديلة إلى الكروموسومات المتسلسلة لإطلاقها من بعضها البعض، ثم يعيد ختم الحمض النووي |
التمارين\(\PageIndex{2}\)
- ما الإنزيم الذي يكسر الروابط الهيدروجينية التي تربط شريطي الحمض النووي معًا بحيث يمكن أن يحدث التكرار؟
- هل هو الخيط المتخلف أو الخيط الرئيسي الذي يتم تصنيعه في الاتجاه نحو فتحة شوكة النسخ المتماثل؟
- ما الإنزيم المسؤول عن إزالة بادئات الحمض النووي الريبي في الحمض النووي البكتيري المنسوخ حديثًا؟
تكرار الحمض النووي في حقيقيات النوى
تعتبر الجينومات حقيقية النواة أكثر تعقيدًا وأكبر من الجينومات بدائية النواة وتتكون عادةً من كروموسومات خطية متعددة (الجدول\(\PageIndex{2}\)). يحتوي الجينوم البشري، على سبيل المثال، على 3 مليارات زوج أساسي لكل مجموعة أحادية الصبغيات من الكروموسومات، ويتم إدخال 6 مليارات زوج أساسي أثناء النسخ المتماثل. هناك أصول متعددة للتكرار على كل كروموسوم حقيقي النواة (الشكل\(\PageIndex{5}\))؛ يحتوي الجينوم البشري على 30.000 إلى 50,000 أصل من النسخ المتماثل. يبلغ معدل النسخ المتماثل حوالي 100 نيوكليوتيدات في الثانية - أبطأ بعشر مرات من النسخ المبكر النواة.
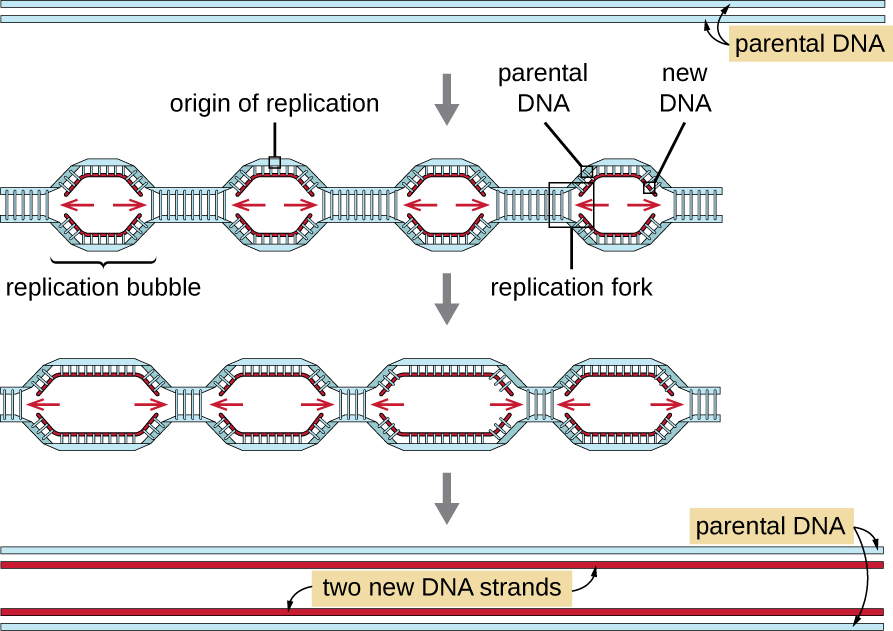
الخطوات الأساسية للتكرار في حقيقيات النوى هي نفسها كما في بدائيات النواة. قبل أن يبدأ النسخ المتماثل، يجب إتاحة الحمض النووي كنموذج. يتم تغليف الحمض النووي حقيقي النواة وتعبئته بشكل كبير، وهو ما تسهله العديد من البروتينات، بما في ذلك الهستونات (انظر بنية ووظيفة الجينومات الخلوية). في بداية النسخ المتماثل، يقوم مركب التكرار المسبق المكون من عدة بروتينات، بما في ذلك الهيليكاس، بتكوين وتوظيف إنزيمات أخرى تشارك في بدء التكاثر، بما في ذلك توبويزوميراز لتخفيف الالتواء الفائق وبروتين الربط أحادي الجديلة وبريماز الحمض النووي الريبي وبلمرة الحمض النووي الريبي. بعد بدء النسخ المتماثل، في عملية مماثلة لتلك الموجودة في بدائيات النواة، يتم تسهيل الاستطالة بواسطة بوليميرات الحمض النووي حقيقية النواة. يتم تصنيع الخيط الرئيسي باستمرار بواسطة إنزيم البوليميراز حقيقي النواة pol، بينما يتم تصنيع الخيط المتخلف بواسطة pol. يحافظ بروتين المشبك المنزلق على بوليميراز الحمض النووي في مكانه حتى لا يسقط من الحمض النووي. يقوم إنزيم الريبونوكلياز H (RNase H)، بدلاً من بوليميراز الحمض النووي كما هو الحال في البكتيريا، بإزالة مادة RNA الأولية، والتي يتم استبدالها بعد ذلك بنيوكليوتيدات الحمض النووي. يتم سد الفجوات المتبقية بواسطة ليغاز الحمض النووي.
نظرًا لأن الكروموسومات حقيقية النواة خطية، فقد يتوقع المرء أن يكون تكرارها أكثر وضوحًا. كما هو الحال في بدائيات النواة، يمكن لبوليميراز الحمض النووي حقيقي النواة إضافة النيوكليوتيدات فقط في اتجاه 5 إلى 3 بوصات. في الخيط الرئيسي، يستمر التخليق حتى يصل إما إلى نهاية الكروموسوم أو شوكة نسخ أخرى تتقدم في الاتجاه المعاكس. على الخيط المتخلف، يتم تصنيع الحمض النووي في فترات قصيرة، يتم بدء كل منها بواسطة برايمر منفصل. عندما تصل شوكة النسخ المتماثل إلى نهاية الكروموسوم الخطي، لا يوجد مكان لعمل دليل تمهيدي لشظية الحمض النووي ليتم نسخها في نهاية الكروموسوم. وبالتالي تظل هذه الأطراف غير مزدوجة، ومع مرور الوقت، قد تصبح أقصر تدريجيًا مع استمرار انقسام الخلايا.
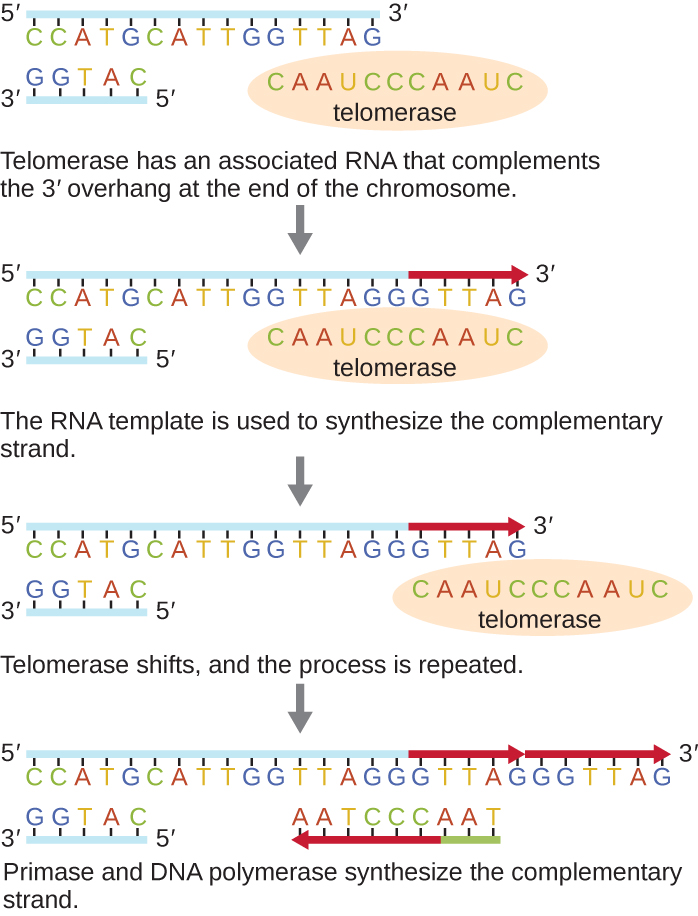
تُعرف نهايات الكروموسومات الخطية باسم التيلوميرات وتتكون من تسلسلات متكررة غير مشفرة. تحمي التيلوميرات تسلسلات الترميز من الضياع مع استمرار انقسام الخلايا. في البشر، يتم تكرار سلسلة مكونة من ستة أزواج أساسية، TTAGGG، من 100 إلى 1000 مرة لتشكيل التيلومير. أوضح اكتشاف إنزيم التيلوميراز (الشكل\(\PageIndex{6}\)) فهمنا لكيفية الحفاظ على نهايات الكروموسومات. يحتوي التيلوميراز على جزء تحفيزي وقالب RNA مدمج. يتم توصيله بنهاية الكروموسوم، وتتم إضافة القواعد التكميلية لقالب الحمض النووي الريبي على الطرف الثالث من خيط الحمض النووي. بمجرد أن يكون الطرف الثالث من قالب الخيط المتخلف ممدودًا بدرجة كافية، يمكن لبوليميراز الحمض النووي إضافة النيوكليوتيدات المكملة لنهايات الكروموسومات. بهذه الطريقة، يتم تكرار نهايات الكروموسومات. في البشر، ينشط التيلوميراز عادةً في الخلايا الجرثومية والخلايا الجذعية البالغة؛ وهو غير نشط في الخلايا الجسدية البالغة وقد يرتبط بشيخوخة هذه الخلايا. كما تنتج الميكروبات حقيقية النواة بما في ذلك الفطريات والبروتوزوان التيلوميراز للحفاظ على سلامة الكروموسومات. لاكتشافها التيلوميراز وعمله، حصلت إليزابيث بلاكبيرن (1948—) على جائزة نوبل للطب أو علم وظائف الأعضاء في عام 2009.
| الملكية | بكتيريا | حقيقيات النوى |
|---|---|---|
| بنية الجينوم | كروموسوم دائري واحد | كروموسومات خطية متعددة |
| عدد الأصول لكل كروموسوم | مفرد | متعددة |
| معدل النسخ | 1000 نيوكليوتيدات في الثانية | 100 نيوكليوتيدات في الثانية |
| تيلوميراز | غير موجود | الحاضر |
| إزالة الحمض النووي الريبي | عينة الحمض النووي الأولى | رانيس إتش |
| استطالة ستراند | مجموعة الحمض النووي 3 | بول، بول |
التمارين\(\PageIndex{3}\)
- كيف يختلف أصل النسخ المتماثل بين حقيقيات النواة وبدائيات النواة؟
- ما هي إنزيمات البوليميراز المسؤولة عن تخليق الحمض النووي أثناء التكاثر حقيقي النواة؟
- ما الذي يوجد في نهايات الكروموسومات في حقيقيات النوى ولماذا؟
تكرار الحمض النووي للعناصر خارج الكروموسومات: البلازميدات والفيروسات
لنسخ الأحماض النووية، تستخدم البلازميدات والفيروسات بشكل متكرر اختلافات في نمط تكرار الحمض النووي الموصوف لجينومات بدائيات النواة. لمزيد من المعلومات حول مجموعة واسعة من استراتيجيات تكرار الفيروس، راجع دورة الحياة الفيروسية.
النسخ المتماثل للدائرة الدوارة
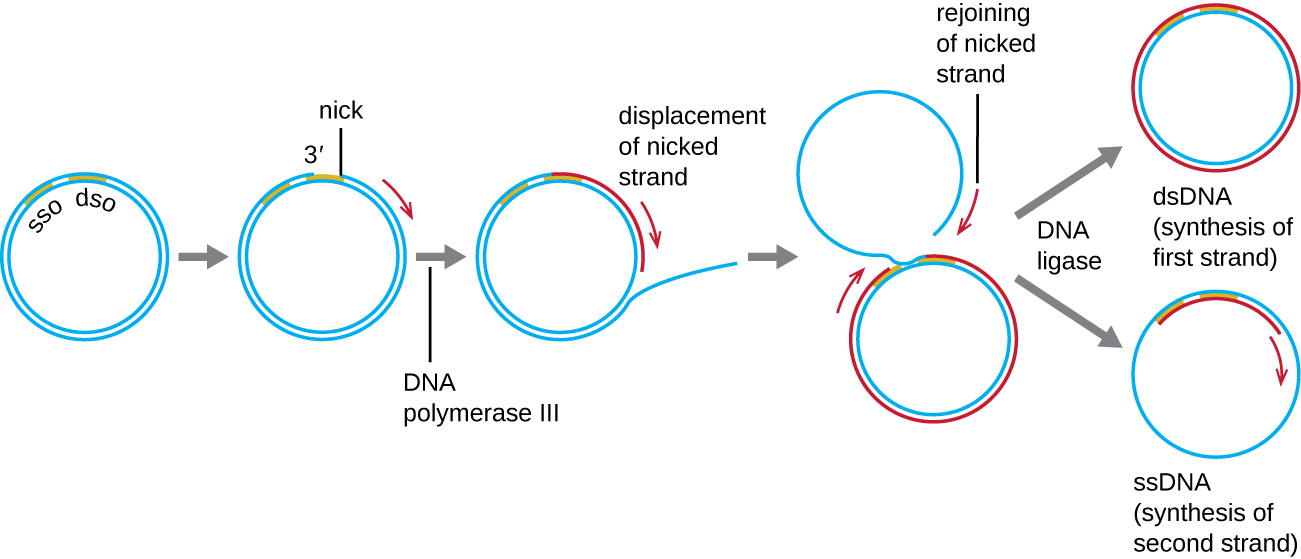
في حين أن العديد من البلازميدات البكتيرية (انظر الخصائص الفريدة للخلايا بدائية النواة) تتكاثر من خلال عملية مشابهة لتلك المستخدمة لنسخ الكروموسوم البكتيري، فإن البلازميدات الأخرى والعديد من البكتيريا وبعض فيروسات حقيقيات النوى تستخدم النسخ المتماثل الدائري (الشكل\(\PageIndex{7}\)). إن الطبيعة الدائرية للبلازميدات وتدوير بعض الجينومات الفيروسية عند العدوى تجعل هذا ممكنًا. يبدأ تكرار الدائرة الدوارة بالربط الأنزيمي لخيط واحد من الجزيء الدائري المزدوج في موقع الأصل المزدوج (dso). في البكتيريا، يرتبط بوليميراز الحمض النووي III بمجموعة 3'-OH من الخصلة المشدودة ويبدأ في تكرار الحمض النووي في اتجاه واحد باستخدام الخصلة غير المحززة كقالب، مما يؤدي إلى إزاحة الخصلة المشدودة أثناء قيامها بذلك. يؤدي الانتهاء من تكرار الحمض النووي في موقع الشق الأصلي إلى الإزاحة الكاملة للحبل المشقوق، والذي قد يتم إعادة تدويره بعد ذلك إلى جزيء DNA أحادي الجديلة. يقوم RNA primase بعد ذلك بتجميع مادة أولية لبدء تكرار الحمض النووي في موقع الأصل أحادي الجديلة (sso) لجزيء الحمض النووي أحادي الجديلة (ssDNA)، مما ينتج عنه جزيء DNA مزدوج السلاسل (dSDDNA) مطابق لجزيء الحمض النووي الدائري الآخر.
التمارين\(\PageIndex{4}\)
هل هناك خيط متخلف في تكرار الدائرة الدوارة؟ لماذا أو لماذا لا؟
المفاهيم الأساسية والملخص
- عملية تكرار الحمض النووي هي عملية شبه محافظة، مما ينتج عنه جزيئان من الحمض النووي، يحتوي كل منهما على خيط أبوي واحد من الحمض النووي وخيوط واحدة مركبة حديثًا.
- في البكتيريا، يحدث بدء التكاثر عند أصل التكاثر، حيث يتم فك الحمض النووي فائق اللفائف بواسطة جيراز الحمض النووي، ويتم تصنيعه بمفرده بواسطة الهليكاز، ويتم ربطه ببروتين رابط أحادي الجديلة للحفاظ على تركيبته أحادية الجديلة دولة تقطعت بها السبل. يقوم Primase بتجميع كتاب تمهيدي قصير للحمض النووي الريبي، مما يوفر مجموعة 3'-OH مجانية يمكن أن يضيف إليها بوليميراز الحمض النووي III نيوكليوتيدات الحمض النووي.
- أثناء الاستطالة، يتم تصنيع الخيط الرئيسي من الحمض النووي بشكل مستمر من برايمر واحد. يتم تصنيع الخيط المتخلف بشكل متقطع في أجزاء أوكازاكي القصيرة، كل منها يتطلب برايمر خاص به. تتم إزالة بادئات الحمض النووي الريبي واستبدالها بنيوكليوتيدات الحمض النووي بواسطة بوليميراز الحمض النووي البكتيري I، ويقوم ليغاز الحمض النووي بسد الفجوات بين هذه الشظايا.
- يتضمن إنهاء التكاثر في البكتيريا تحليل سلاسل الحمض النووي الدائرية بواسطة topoisomerase IV لإطلاق نسختين من الكروموسوم الدائري.
- عادةً ما تحتوي حقيقيات النوى على كروموسومات خطية متعددة، ولكل منها أصول متعددة للتكرار. وبشكل عام، فإن النسخ المتماثل في حقيقيات النوى يشبه النسخ المتماثل في بدائيات النواة.
- تتطلب الطبيعة الخطية للكروموسومات حقيقية النواة التيلوميرات لحماية الجينات بالقرب من نهاية الكروموسومات. يعمل التيلوميراز على توسيع التيلوميرات، مما يمنع تدهورها، في بعض أنواع الخلايا.
- تكرار الدائرة الدوارة هو نوع من تخليق الحمض النووي السريع أحادي الاتجاه لجزيء الحمض النووي الدائري المستخدم لتكرار بعض البلازميدات.