
4.3: دورة حمض الستريك والفسفرة التأكسدية
- Page ID
- 191711

دورة حمض الستريك
في الخلايا حقيقية النواة، يتم نقل جزيئات البيروفات المنتجة في نهاية تحلل السكر إلى الميتوكوندريا، وهي مواقع للتنفس الخلوي. إذا كان الأكسجين متاحًا، فسيمضي التنفس الهوائي إلى الأمام. في الميتوكوندريا، سيتم تحويل البيروفات إلى مجموعة أسيتيل ثنائية الكربون (عن طريق إزالة جزيء من ثاني أكسيد الكربون) سيتم التقاطها بواسطة مركب حامل يسمى الإنزيم المساعد A (CoA)، وهو مصنوع من فيتامين ب 5. يسمى المركب الناتج أسيتيل CoA. (الشكل\(\PageIndex{1}\)). يمكن استخدام Acetyl CoA بعدة طرق من قبل الخلية، ولكن وظيفته الرئيسية هي توصيل مجموعة الأسيتيل المشتقة من البيروفات إلى المسار التالي في تقويض الجلوكوز.
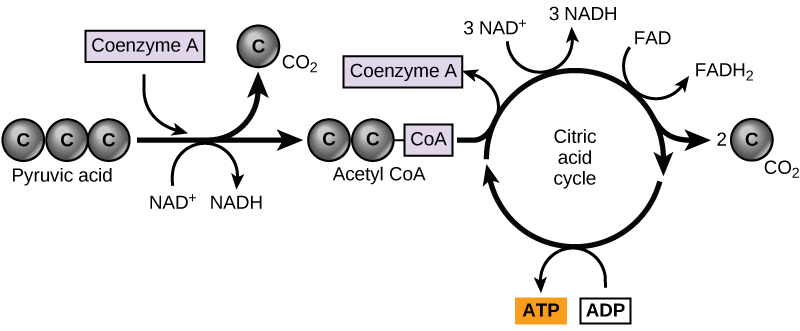
مثل تحويل البيروفات إلى أسيتيل CoA، تحدث دورة حمض الستريك في الخلايا حقيقية النواة في مصفوفة الميتوكوندريا. على عكس تحلل السكر، فإن دورة حمض الستريك عبارة عن حلقة مغلقة: الجزء الأخير من المسار يجدد المركب المستخدم في الخطوة الأولى. الخطوات الثماني للدورة هي سلسلة من التفاعلات الكيميائية التي تنتج جزيئين من ثاني أكسيد الكربون، وجزيء ATP واحد (أو ما يعادله)، وأشكال مخفضة (NADH و FADH 2) من NAD+ و FAD+، وهي إنزيمات مساعدة مهمة في الخلية. يعتبر جزء من هذا مسارًا هوائيًا (يتطلب الأكسجين) لأن NADH و FADH 2 المنتجين يجب أن ينقلوا إلكتروناتهم إلى المسار التالي في النظام، والذي سيستخدم الأكسجين. في حالة عدم وجود الأكسجين، لا يحدث هذا النقل.
تدخل ذرتان من الكربون في دورة حمض الستريك من كل مجموعة أسيتيل. يتم إطلاق جزيئين من ثاني أكسيد الكربون في كل دورة؛ ومع ذلك، لا تحتوي هذه الجزيئات على نفس ذرات الكربون التي تساهم بها مجموعة الأسيتيل في هذا المنعطف من المسار. سيتم إطلاق ذرتي الأسيتيل والكربون في نهاية المطاف في دورات لاحقة من الدورة؛ بهذه الطريقة، سيتم إطلاق جميع ذرات الكربون الست من جزيء الجلوكوز الأصلي في نهاية المطاف على شكل ثاني أكسيد الكربون. يستغرق الأمر دورتين من الدورة لمعالجة ما يعادل جزيء جلوكوز واحد. كل دورة من الدورة تشكل ثلاثة جزيئات NADH عالية الطاقة وجزيء FADH 2 عالي الطاقة. سوف تتصل هذه الناقلات عالية الطاقة بالجزء الأخير من التنفس الهوائي لإنتاج جزيئات ATP. يتم أيضًا إجراء ATP واحد (أو ما يعادله) في كل دورة. يمكن استخدام العديد من المركبات الوسيطة في دورة حمض الستريك في تصنيع الأحماض الأمينية غير الأساسية؛ لذلك، تكون الدورة ابتنائية وتقويضية.
الفسفرة التأكسدية
لقد قرأت للتو عن مسارين في هدم الجلوكوز - تحلل السكر ودورة حمض الستريك - اللذان يولدان ATP. ومع ذلك، فإن معظم ATP المتولد أثناء الهدم الهوائي للجلوكوز لا يتولد مباشرة من هذه المسارات. بل إنها تنبع من عملية تبدأ بتمرير الإلكترونات عبر سلسلة من التفاعلات الكيميائية إلى مستقبل الإلكترون النهائي، الأكسجين. تحدث هذه التفاعلات في مجمعات بروتينية متخصصة تقع في الغشاء الداخلي للميتوكوندريا للكائنات حقيقية النواة وعلى الجزء الداخلي من غشاء الخلية للكائنات بدائية النواة. يتم حصاد طاقة الإلكترونات واستخدامها لتوليد تدرج كهروكيميائي عبر غشاء الميتوكوندريا الداخلي. يتم استخدام الطاقة الكامنة لهذا التدرج لتوليد ATP. وتسمى هذه العملية برمتها بالفسفرة التأكسدية.
سلسلة نقل الإلكترون (الشكل\(\PageIndex{2}\) أ) هي المكون الأخير للتنفس الهوائي وهي الجزء الوحيد من عملية التمثيل الغذائي الذي يستخدم الأكسجين الجوي. ينتشر الأكسجين باستمرار في النباتات لهذا الغرض. في الحيوانات، يدخل الأكسجين الجسم من خلال الجهاز التنفسي. نقل الإلكترون هو سلسلة من التفاعلات الكيميائية التي تشبه لواء الجرافة حيث يتم تمرير الإلكترونات بسرعة من مكون إلى آخر، إلى نقطة نهاية السلسلة حيث يكون الأكسجين هو متقبل الإلكترون النهائي ويتم إنتاج الماء. هناك أربعة مجمعات تتكون من البروتينات، المصنفة من الأول إلى الرابع في الشكل\(\PageIndex{2}\) ج، ويسمى تجميع هذه المجمعات الأربعة، جنبًا إلى جنب مع حاملات الإلكترون المتنقلة والملحقات المرتبطة بها، بسلسلة نقل الإلكترون. توجد سلسلة نقل الإلكترون في نسخ متعددة في غشاء الميتوكوندريا الداخلي لحقيقيات النوى وفي الغشاء البلازمي لبدائيات النواة. في كل عملية نقل للإلكترون عبر سلسلة نقل الإلكترون، يفقد الإلكترون الطاقة، ولكن مع بعض عمليات النقل، يتم تخزين الطاقة كطاقة محتملة باستخدامها لضخ أيونات الهيدروجين عبر غشاء الميتوكوندريا الداخلي إلى الفضاء بين الأغشية، مما يؤدي إلى حدوث تدرج كهروكيميائي.
آرت كونيكشن
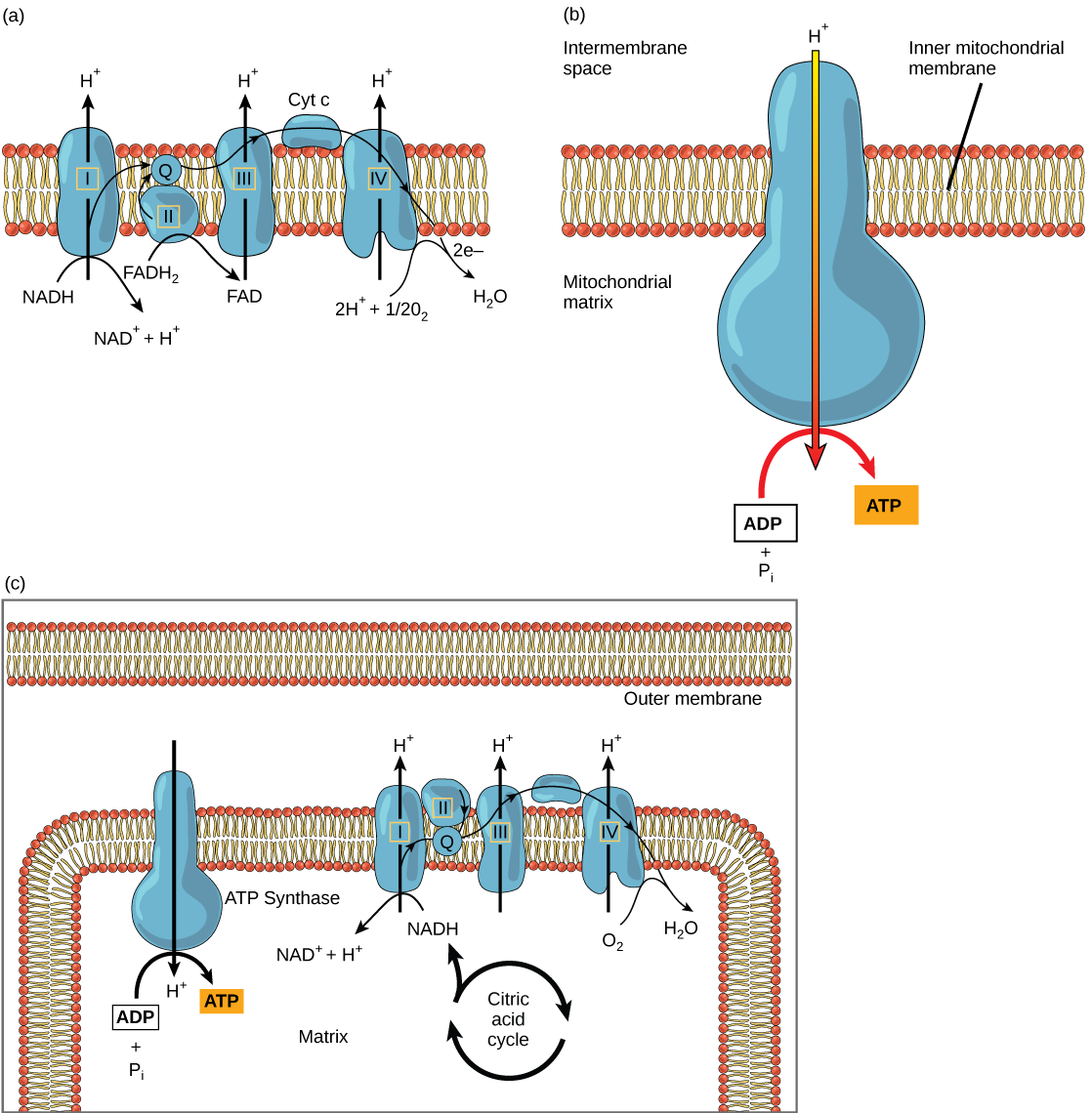
يمنع السيانيد أوكسيديز السيتوكروم سي، وهو أحد مكونات سلسلة نقل الإلكترون. في حالة حدوث تسمم بالسيانيد، هل تتوقع زيادة أو نقصان درجة الحموضة في الفضاء بين الأغشية؟ ما تأثير السيانيد على تخليق ATP؟
يتم تمرير الإلكترونات من NADH و FADH 2 إلى مجمعات البروتين في سلسلة نقل الإلكترون. عند انتقالها من مركب إلى آخر (هناك ما مجموعه أربعة)، تفقد الإلكترونات الطاقة، وتستخدم بعض هذه الطاقة لضخ أيونات الهيدروجين من مصفوفة الميتوكوندريا إلى الفضاء بين الأغشية. في مجمع البروتين الرابع، يتم قبول الإلكترونات بواسطة الأكسجين، وهو المستقبل النهائي. ثم يتحد الأكسجين مع إلكتروناته الإضافية مع اثنين من أيونات الهيدروجين، مما يعزز التدرج الكهروكيميائي، لتكوين الماء. إذا لم يكن هناك أكسجين موجود في الميتوكوندريا، فلا يمكن إزالة الإلكترونات من النظام، وستعود سلسلة نقل الإلكترون بأكملها وتتوقف. لن تتمكن الميتوكوندريا من توليد ATP جديد بهذه الطريقة، وستموت الخلية في النهاية بسبب نقص الطاقة. هذا هو السبب في أننا يجب أن نتنفس لسحب الأكسجين الجديد.
في سلسلة نقل الإلكترون، تُستخدم الطاقة الحرة من سلسلة التفاعلات الموصوفة للتو لضخ أيونات الهيدروجين عبر الغشاء. يؤدي التوزيع غير المتكافئ لأيونات H + عبر الغشاء إلى حدوث تدرج كهروكيميائي، بسبب الشحنة الموجبة لأيونات H + وتركيزها العالي على جانب واحد من الغشاء.
تنتشر أيونات الهيدروجين عبر الغشاء الداخلي من خلال بروتين غشائي متكامل يسمى ATP synthase (الشكل\(\PageIndex{2}\) ب). يعمل هذا البروتين المعقد كمولد صغير، يتم تشغيله بواسطة قوة أيونات الهيدروجين المنتشرة عبره، وصولاً إلى التدرج الكهروكيميائي من الفضاء بين الأغشية، حيث يوجد العديد من أيونات الهيدروجين التي تطرد بشكل متبادل إلى المصفوفة، حيث يوجد القليل منها. يؤدي دوران أجزاء هذه الآلة الجزيئية إلى تجديد ATP من ADP. هذا التدفق من أيونات الهيدروجين عبر الغشاء من خلال سينثاس ATP يسمى التناضح الكيميائي.
يستخدم التناضح الكيميائي (الشكل\(\PageIndex{2}\) ج) لتوليد 90 في المائة من ATP الذي يتم إنتاجه أثناء تقويض الجلوكوز الهوائي. نتيجة التفاعلات هي إنتاج ATP من طاقة الإلكترونات التي تمت إزالتها من ذرات الهيدروجين. كانت هذه الذرات في الأصل جزءًا من جزيء الجلوكوز. في نهاية نظام نقل الإلكترون، تُستخدم الإلكترونات لتقليل جزيء الأكسجين إلى أيونات الأكسجين. تجذب الإلكترونات الإضافية على أيونات الأكسجين أيونات الهيدروجين (البروتونات) من الوسط المحيط، ويتشكل الماء. يُطلق على سلسلة نقل الإلكترون وإنتاج ATP من خلال التناضح الكيميائي مجتمعة اسم الفسفرة التأكسدية.
عائد ATP
يختلف عدد جزيئات ATP الناتجة عن تقويض الجلوكوز. على سبيل المثال، يختلف عدد أيونات الهيدروجين التي يمكن لمجمعات سلسلة نقل الإلكترون ضخها عبر الغشاء بين الأنواع. مصدر آخر للتباين ينبع من مكوك الإلكترونات عبر غشاء الميتوكوندريا. لا يمكن لـ NADH الناتج عن تحلل السكر دخول الميتوكوندريا بسهولة. وبالتالي، يتم التقاط الإلكترونات من داخل الميتوكوندريا إما عن طريق NAD + أو FAD +. يتم إنشاء عدد أقل من جزيئات ATP عندما يعمل FAD+ كحامل. يستخدم NAD+ كناقل للإلكترون في الكبد و FAD+ في الدماغ، لذلك يعتمد عائد ATP على الأنسجة التي يتم أخذها في الاعتبار.
هناك عامل آخر يؤثر على إنتاج جزيئات ATP الناتجة عن الجلوكوز وهو أن المركبات الوسيطة في هذه المسارات تستخدم لأغراض أخرى. يرتبط هدم الجلوكوز بالمسارات التي تبني أو تكسر جميع المركبات البيوكيميائية الأخرى في الخلايا، والنتيجة أكثر فوضوية إلى حد ما من الحالات المثالية الموصوفة حتى الآن. على سبيل المثال، يتم إدخال السكريات الأخرى غير الجلوكوز في المسار الجليكوليتي لاستخراج الطاقة. يمكن إزالة الجزيئات الأخرى التي يمكن استخدامها لحصاد الطاقة في تحلل السكر أو دورة حمض الستريك لتكوين الأحماض النووية أو الأحماض الأمينية أو الدهون أو المركبات الأخرى. بشكل عام، في الأنظمة الحية، تستخرج مسارات هدم الجلوكوز هذه حوالي 34 بالمائة من الطاقة الموجودة في الجلوكوز.
وظائف قيد العمل: طبيب أمراض الميتوكوندريا
ماذا يحدث عندما لا تستمر التفاعلات الحرجة للتنفس الخلوي بشكل صحيح؟ أمراض الميتوكوندريا هي اضطرابات وراثية في التمثيل الغذائي. يمكن أن تنشأ اضطرابات الميتوكوندريا من الطفرات في الحمض النووي النووي أو الميتوكوندريا، وتؤدي إلى إنتاج طاقة أقل مما هو طبيعي في خلايا الجسم. يمكن أن تشمل أعراض أمراض الميتوكوندريا ضعف العضلات ونقص التنسيق ونوبات تشبه السكتة الدماغية وفقدان الرؤية والسمع. يتم تشخيص معظم الأشخاص المصابين في مرحلة الطفولة، على الرغم من وجود بعض الأمراض التي تصيب البالغين. يعد تحديد وعلاج اضطرابات الميتوكوندريا مجالًا طبيًا متخصصًا. يتطلب الإعداد التعليمي لهذه المهنة تعليمًا جامعيًا، تليها كلية الطب مع التخصص في علم الوراثة الطبية. يمكن لعلماء الوراثة الطبية الحصول على شهادة البورد الأمريكي لعلم الوراثة الطبية والاستمرار في الارتباط بالمنظمات المهنية المكرسة لدراسة مرض الميتوكوندريا، مثل جمعية طب الميتوكوندريا وجمعية أمراض التمثيل الغذائي الموروثة.
ملخص
دورة حمض الستريك عبارة عن سلسلة من التفاعلات الكيميائية التي تزيل الإلكترونات عالية الطاقة وتستخدمها في سلسلة نقل الإلكترون لتوليد ATP. يتم إنتاج جزيء واحد من ATP (أو ما يعادله) لكل دورة من الدورة.
سلسلة نقل الإلكترون هي جزء من التنفس الهوائي الذي يستخدم الأكسجين الحر كمستقبل للإلكترون النهائي للإلكترونات التي تمت إزالتها من المركبات الوسيطة في تقويض الجلوكوز. يتم تمرير الإلكترونات من خلال سلسلة من التفاعلات الكيميائية، مع استخدام كمية صغيرة من الطاقة الحرة في ثلاث نقاط لنقل أيونات الهيدروجين عبر الغشاء. هذا يساهم في التدرج المستخدم في التناضح الكيميائي. عندما تنتقل الإلكترونات من NADH أو FADH 2 إلى أسفل سلسلة نقل الإلكترون، فإنها تفقد الطاقة. منتجات سلسلة نقل الإلكترون هي الماء و ATP. يمكن تحويل عدد من المركبات الوسيطة إلى استقلاب الجزيئات البيوكيميائية الأخرى، مثل الأحماض النووية والأحماض الأمينية غير الأساسية والسكريات والدهون. يمكن أن تعمل هذه الجزيئات نفسها، باستثناء الأحماض النووية، كمصادر للطاقة لمسار الجلوكوز.
اتصالات فنية
الشكل\(\PageIndex{2}\): يمنع السيانيد أوكسيديز السيتوكروم سي، وهو أحد مكونات سلسلة نقل الإلكترون. في حالة حدوث تسمم بالسيانيد، هل تتوقع زيادة أو نقصان درجة الحموضة في الفضاء بين الأغشية؟ ما تأثير السيانيد على تخليق ATP؟
- إجابة
-
بعد التسمم بالسيانيد، لم تعد سلسلة نقل الإلكترون قادرة على ضخ الإلكترونات في الفضاء بين الأغشية. ستزداد درجة الحموضة في الفضاء بين الأغشية، وسيتوقف تخليق ATP.
مسرد المصطلحات
- أسيتيل أوكا
- مزيج من مجموعة الأسيتيل المشتقة من حمض البيروفيك والإنزيم المساعد A المصنوع من حمض البانتوثينك (فيتامين مجموعة B)
- سينثاز ATP
- مركب بروتيني مضمن في الغشاء يجدد ATP من ADP بالطاقة من البروتونات المنتشرة عبره
- التناضح الكيميائي
- حركة أيونات الهيدروجين إلى أسفل تدرجها الكهروكيميائي عبر الغشاء من خلال تركيب ATP لتوليد ATP
- دورة حمض الستريك
- سلسلة من التفاعلات الكيميائية المحفزة بالإنزيم ذات الأهمية المركزية في جميع الخلايا الحية التي تحصد الطاقة في روابط الكربون والكربون لجزيئات السكر لتوليد ATP؛ دورة حمض الستريك هي مسار استقلابي هوائي لأنها تتطلب الأكسجين في التفاعلات اللاحقة للمتابعة
- سلسلة نقل الإلكترون
- سلسلة من أربعة مجمعات كبيرة متعددة البروتينات مدمجة في غشاء الميتوكوندريا الداخلي الذي يقبل الإلكترونات من المركبات المانحة ويحصد الطاقة من سلسلة من التفاعلات الكيميائية لتوليد تدرج أيون الهيدروجين عبر الغشاء
- الفسفرة التأكسدية
- إنتاج ATP عن طريق نقل الإلكترونات إلى أسفل سلسلة نقل الإلكترون لإنشاء تدرج بروتوني يستخدمه سينثاس ATP لإضافة مجموعات الفوسفات إلى جزيئات ADP