
15.8: מיוזה
- Page ID
- 208504
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)במטאזואה ישנם שני מצבים בהם תא מוליד תאי בת. הראשון, והנפוץ ביותר, הוא מיטוזה. השני הוא מיוזה. מיוזיס הוא התהליך שבו נוצרים גמטות (תאי מין). בעלי חיים וצמחים נוצרים על ידי רבייה מינית (אם זה חדשות עבורך, אנא שקול להתמחות במשהו אחר מלבד ביולוגיה). אורגניזמים אלה מתחילים את החיים באמצעות מיזוג של שני תאים: זרע וביצה. שניהם תורמים חומר גנטי לאורגניזם החדש. על מנת לשמור על המספר הנכון של הכרומוזומים בכל דור, הגמטות תורמות כל אחת קבוצה אחת של כרומוזומים, כך שלביצית המופרית ולכל שאר התאים באורגניזם יש שתי קבוצות של כרומוזומים - אחד מכל הורה. מטרת המיוזה, וההבדל העיקרי שלה עם מיטוזה, אינה יצירת תאי בת שהם שכפולים מדויקים, אלא יצירת תאי בת שיש להם רק מחצית מכמות החומר הגנטי כמו התא המקורי.
הבה נסתכל על המצב הזה באנוכיות: מיוזה בבני אדם. כמעט לכל תא בגופך יש גרעין המכיל 46 כרומוזומים, קבוצה של 23 מאביך, וקבוצה של 23 מאמך. היוצאים מן הכלל היחידים הם הגמטות: הזרעונים אצל גברים והביציות אצל נשים. אומרים שהתאים הסומטיים הם 2n או דיפלואידים, כלומר יש 2 קבוצות של כרומוזומים, והגמטות הן 1n או הפלואידיות, שיש להן רק קבוצה אחת של כרומוזומים. לפעמים, מיוזה יכולה להיות קצת מבלבלת את התלמידים מכיוון שהיא מתרחשת באותו חלק של מחזור התא כמו מיטוזה, כלומר אחרי G 2. בגלל זה, לתא הנכנס למיוזה יש למעשה 4 קבוצות של כרומוזומים, מכיוון שה- DNA כבר עבר שכפול בשלב S.
תאי דם אדומים בוגרים אינם מכילים גרעין, וכמה תאי שריר, בעוד שהם מרובי גרעינים מכיוון שהם נוצרים מהתמזגות של כמה מיובלסטים, בכל זאת יש 46 כרומוזומים בכל אחד מהגרעינים. פוליפלואידיה, למרות שהיא נדירה בבני אדם, היא מצב נורמלי עבור אורגניזמים רבים. הצפרדע, Xenopus laevis, חיית מחקר נפוצה, היא טטרפלואידית.
מיוזה מורכבת משתי חטיבות מיוטיות רצופות שלכל אחת מהן שלבים הדומים למיטוזה: פרופאזה, מטאפאזה, אנפאזה, טלופאז, וכל אחת מהן מסתיימת בציטוקינזיס מלא. שימו לב שמיד לאחר הטלופאז המיוטי I, התא מתחלק, ושני תאי הבת נמצאים מיד בפרופזה II. אין שלב G 1, S או G 2 מתערב.
פרופאזה I של מיוזה מתחילה באופן דומה מאוד לפרופזה של מיטוזה: הפעלת MPF (mitotic-cdk), עיבוי כרומוזומים, היווצרות ציר ופירוק מעטפת גרעינית. עם זאת, בהשוואה למיטוזה, פרופאזה מיוטית I נמשכת זמן רב מאוד וניתן לחלק אותה לחמישה שלבים: לפטוטן, זיגוטן, פצ'יטן, דיפלוטן ודיאקינזיס. במהלך הלפטוטן, שתי הקבוצות (אימהיות ואבהיות) של כרומטידות אחיות לכל כרומוזום מתעבות, מתיישרות ויוצרות מבנה המכונה דו ערכי. לשם הבהרה, דו-ערכי זה מורכב מארבעה עותקים של כרומוזום נתון: שני עותקים כל אחד מהכרומוזום האימהי ושל הכרומוזום האבהי. מכיוון שהגרסאות האימהיות והאבהיות של כרומוזום נתון נשמרות בסמיכות רבה לפרק זמן ממושך, יש סיכוי גדול יותר לשילוב מחדש, או מעבר והחלפה של חלקים הומולוגיים של כל כרומוזום.
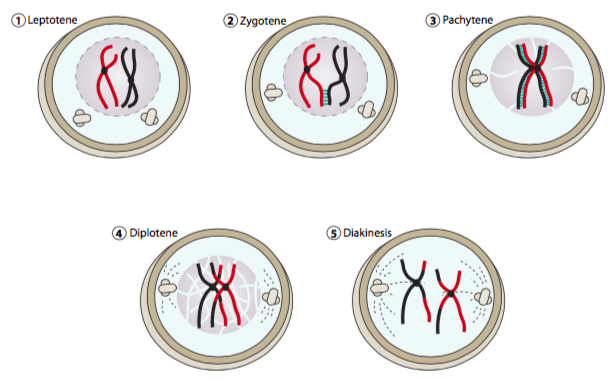
רקומבינציה מתרחשת כאשר חלק מהכרומוזום האבהי מוחלף בחתיכת ה-DNA ההומולוגית בכרומוזום האימהי התואם (או להיפך). שים לב שכרומטידות אחיות (כלומר עותקים מדויקים) אינן מתחברות מחדש - רק כרומטידות הומולוגיות שאינן אחיות יכולות לשלב מחדש. ברור שהחלפת DNA מסוג זה חייבת להיעשות בזהירות ובשוויון, כך שה-DNA המתקבל מכל צד יכיל את כל המידע הגנטי שהוא אמור להכיל, ולא יותר מידע ממה שהוא אמור. על מנת להבטיח דיוק זה ברקומבינציה, הכרומטידות ההומולוגיות שאינן אחיות מוחזקות יחד בקומפלקס סינפטונמלי (SC). קומפלקס דמוי סולם זה מתחיל להיווצר בשלב הזיגוטן של פרופזה I ומשלים בפצ'יטן. ה-SC השלם מורכב מאלמנטים רוחביים חלבוניים (aka אלמנטים ציריים) העוברים לאורך הכרומטידים ואלמנט מרכזי קצר המורכב מחלבונים סיביים היוצרים את שלבי הסולם בניצב לשני היסודות הרוחביים. האלמנט המרכזי נוצר מדימרי קינה רוחביים המקיימים אינטראקציה זה עם זה בצורה מקוזזת, כמו גם עם האלמנטים הרוחביים. לחלבוני קינה אלה (למשל SCP1 (עכבר), Zip1p (שמרים)) יש אזורים מרכזיים של סליל מפותל המתפקדים כתחומי אינטראקציה בין חלבונים. למרות ש- SCP3 ולכן היווצרות אלמנטים רוחביים מלאים מיותרים עבור קומפלקס סינפטונמלי פונקציונלי, נראה כי קונדנסין וקוהזין נחוצים לחיבור קינה רוחבי תקין של האלמנטים הרוחביים.
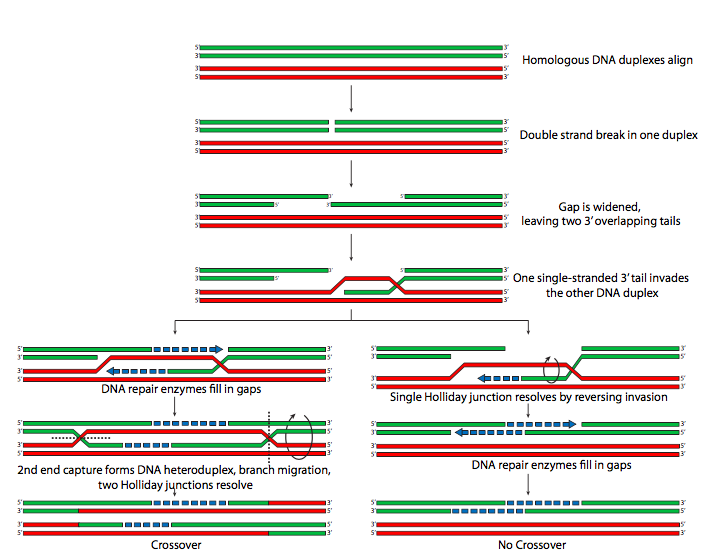
רקומבינציה עשויה להתרחש עם או בלי היווצרות של שבירות כפולות, ולמעשה, יכולה להתרחש ללא היווצרות הקומפלקס הסינפטונמלי, אם כי ה- SC כנראה משפר את יעילות הרקומבינציה. ב S. pombe, מיוזה מתרחשת ללא היווצרות של קומפלקס סינפטונמלי, אך ישנם מבנים קטנים ובלתי רציפים הדומים במקצת לחלקים של ה- SC. בזבוב הפירות, תסיסנית מלנוגסטר, הנקבות עוברות מיוזה באמצעות קומפלקס סינפטונמלי, אך זכרים אינם עוברים רקומבינציה מיוטית, והכרומוזומים שלהם אינם יוצרים קומפלקסים סינפטונמליים. ברוב המקרים, רקומבינציה קדמה להיווצרות גושים רקומבינציה, שהם מתחמי חלבון הנוצרים בנקודות פוטנציאליות לרקומבינציה. המנגנון הנחקר הטוב ביותר לרקומבינציה מיוטית כולל שבירה דו-גדילית של אחד הכרומוזומים שיזם האנדונוקלאז הספציפי למיוזה, Spo11. קצוות 5' (אחד לכל כיוון) של חתך זה מושפלים מעט ליצירת תליונים חד-גדיליים בגודל 3'. זה מוביל להיווצרות צמתים של הולידיי כאשר גדיל מכרומוזום אחד משמש כתבנית לחלק חסר בכרומוזום החתך ההומולוגי. ניתן לפתור זאת באחת משתי דרכים, עם או בלי הצלבה, כפי שמודגם (איור\(\PageIndex{17}\)).
הרקומבינציה מתחילה בפצ'יטן ומשלימה בדיפלוטן, אז הקומפלקס הסינפטונמלי מתפרק. כאשר הכרומטידות מתחילות להיפרד, chias-mata מתגלה בחלק מאתרי הרקומבינציה. עם השלמת הפרופזה, הצ'יאסמאטה נפתרת ממרכז הכרומוזומים עד הקצוות.
כאשר התא עובר מנבואה מיוטית I למטאפאזה מיוטית I, מתגלה הבדל נוסף בין מיטוזה למיוזה: הכרומוזומים מסתדרים בשורה בצלחת המטאפאזה כטטרדים ולא כזוגות. בגלל זה, כשהם מתפרקים באנפאזה, קבוצות של כרומטידות אחיות נפרדות לקטבים מנוגדים. כמובן, עקב רקומבינציה, סביר להניח שהכרומטידות האחיות עדיין לא יהיו זהות.
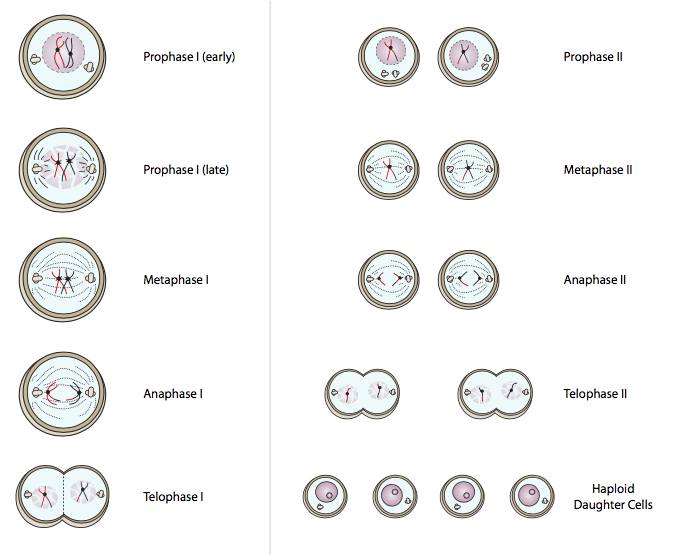
לאחר אנאפאזה וטלופאז קונבנציונלית, התא מתפצל, ומיד תאי הבת מתחילים בחלוקה המיוטית השנייה (איור\(\PageIndex{18}\), צד ימין). בסוגי תאים מסוימים, הכרומוזומים אינם מתעבים בטלופאז I מיוטי, אך אם יש להם, הם מתעבים מחדש בפרופזה מיוטית II. פרופזה II מתקדמת באופן דומה לפרופזה מיטוטית, בכך שאין היווצרות של קומפלקסים סינפטונמליים או רקומבינציה. במטאפאזה II, הכרומטידות האחיות מסתדרות לאורך צלחת המטאפאזה בדיוק כמו במיטוזה, אם כי כעת יש רק 2n כרומוזומים בתא, בעוד שבמיטוזה היו 4n (כי ה-DNA השתכפל). שוב, כשהם מכרסמים את שאר החלוקה כמעט בדיוק כמו מיטוזה, הכרומטידות האחיות מתפרקות באנפאזה II, הגרעין מתחדש בטלופאז II, והציטוקינזיס הסופי מייצר בסך הכל ארבעה תאים מהמקור שנכנס למיוזה, כל אחד מכיל 1n כרומוזומים.
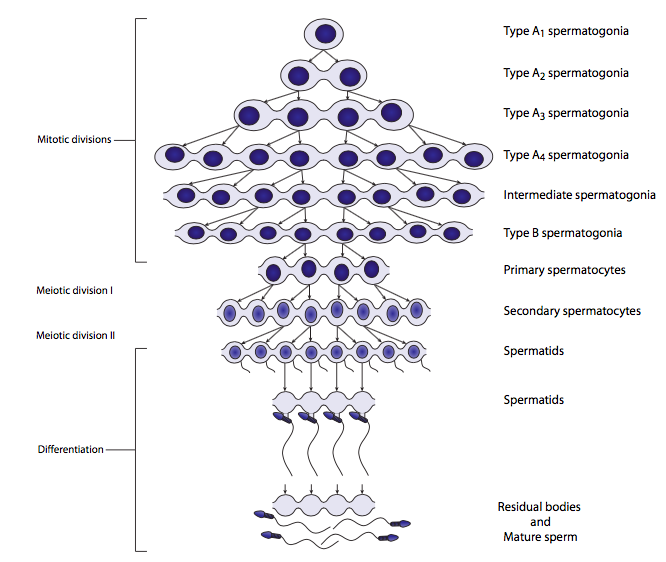
תאי ביצה, כתורמים חומריים גנטיים ותפזורת, צריכים להיות גדולים אך תאי זרע, כתורמים גנטיים בלבד, אינם. התרשים שלהלן מתאר את יצירת תאי הביצה. רק ביצית אחת נוצרת מאירוע מיוטי; שלושת תאי הבת האחרים נקראים גופים קוטביים, ומכילים כל כך מעט חומר ציטופלסמי שהם קיימים רק לזמן קצר. ההתפלגות הא-סימטרית של הציטופלזמה בחלוקה המיוטית הראשונה לביציות נובעת ממיקום הציר המיוטי בפריפריה של התא ולא במרכז. מכיוון שמרכז הציר קובע את מיקום הטבעת המתכווצת לציטוקינזיס, הדבר מוביל לתאי בת בגודל לא אחיד.
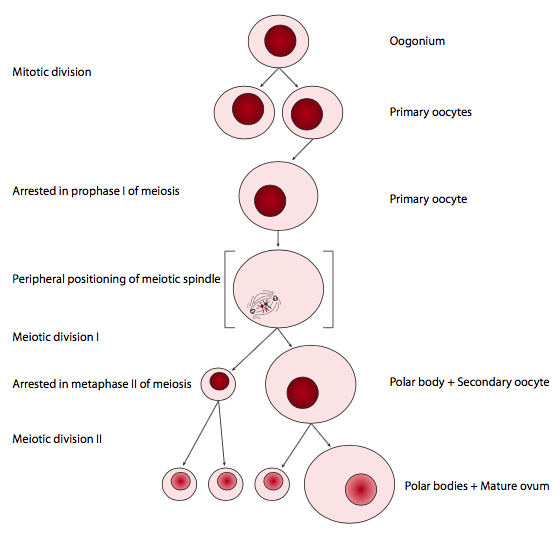
הדור של הזרע הקטן מאוד הוא מנגנון אחר לגמרי. בשלבים המיוטיים של spermatogenesis, חלוקות התאים שוות, כאשר הציר המיוטי מיושר למרכז התא, ולתאים יש כמויות שוות של ציטופלזמה, בדומה לתא ממוצע שעבר מיטוזה. הזרע הבוגר היעיל והמינימלי-ציטופלזמה הוא תוצר של התמיינות פוסט-מיוטית, שבה הוא זוכה בזנב הדגל, ופולט את רוב החומר הציטופלזמי שלו, ושומר רק על חלק מהמיטוכונדריה להניע את הדגל, ושלפוחית אקרוזומלית, המכילה את אנזימים ומולקולות אחרות הדרושות כדי להגיע ולהתמזג עם (כלומר להפרות) ביצית.
לא כל האורגניזמים מתרבים עם מנגנון הביצית והזרע דמוי האדם, כלומר מיוזה גמטית. כפי שתואר זה עתה, במחזור חיים של מיוזה גמטית, מיוזה מייצרת גמטות הפלואידיות, אשר לאחר מכן מתמזגות/מפרות כדי להפוך לזיגוטה דיפלואידית. הזיגוטה הופכת לאורגניזם דיפלואידי רב תאי, וברגע שהיא מגיעה לבגרות מינית יכולה ליצור יותר גמטות הפלואידיות באמצעות מיוזה. המצב הרב-תאי היחיד הוא דיפלואידי, והגמטות הן הפלואידיות.
וריאציה נפוצה היא מיוזה ספורית, המשמשת בכל הצמחים וסוגים רבים של אצות. בשימוש זה, "נבג" מתייחס לנבגים אוקריוטיים, ולא לאנדוספורים חיידקיים, שהם פשוט חיידקים רדומים. מיוזה ספורית אינה מייצרת ישירות גמטות. במקום זאת, מיוזה מייצרת נבגים הפלואידים, שיכולים להתפתח על ידי מיטוזה באורגניזמים רב-תאיים הפלואידים. אורגניזמים אלה (המכונים גמטופיטים) יכולים לייצר גמטות (עדיין הפלואידיות) על ידי מיטוזה, שכאשר מתמזגים/מופרים יוצרים זיגוטה דיפלואידית. לאחר מכן זיגוטה זו יכולה להתפתח לצורה רב-תאית דיפלואידית הנקראת ספורופיט. לבסוף, הספורופיט מסוגל לייצר נבגים נוספים על ידי מיוזה.
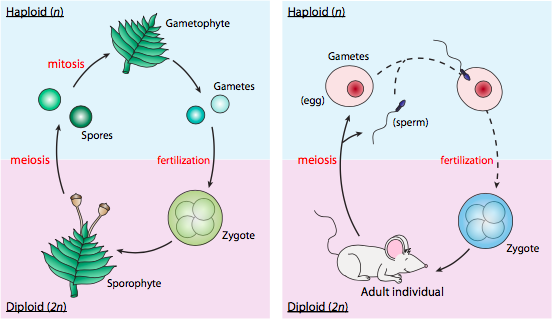
דוגמה לסוג זה של מחזור חיים ותפקיד המיוזה נמצאת בטחב. מה שאנו חושבים עליו כגוף הטחב הוא למעשה גמטופיט, המורכב מתאים הפלואידים הנוצרים על ידי חלוקה מיטוטית של נבג הפלואידי. גמטופיטים אלה מייצרים זרע או ביצים במבנים מיוחדים בקצותיהם הדיסטליים, ובתנאים הנכונים (למשל גשם) הזרע נישא לביצים ומתרחשת הפריה. הביצית המופרית (הדיפלואידית) מתפתחת כעת על ידי חלוקה מיטוטית והתמיינות לספורופיט. במקרה זה, הספורופיט הוא מבנה רבייה מיוחד בקצה הטחב, והוא גם דיפלואידי. בקצה הספורופיט נמצא הספורנגיום, שם מתרחשת מיוזה ליצירת נבגים הפלואידים. לאחר מכן ניתן לפזר את הנבגים (על ידי רוח או גשם) ולהתחיל את המחזור שוב על ידי חלוקה ויצירת גמטופיט חדש.