12.6: הובלה על שלד הציטוס
- Page ID
- 208446

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
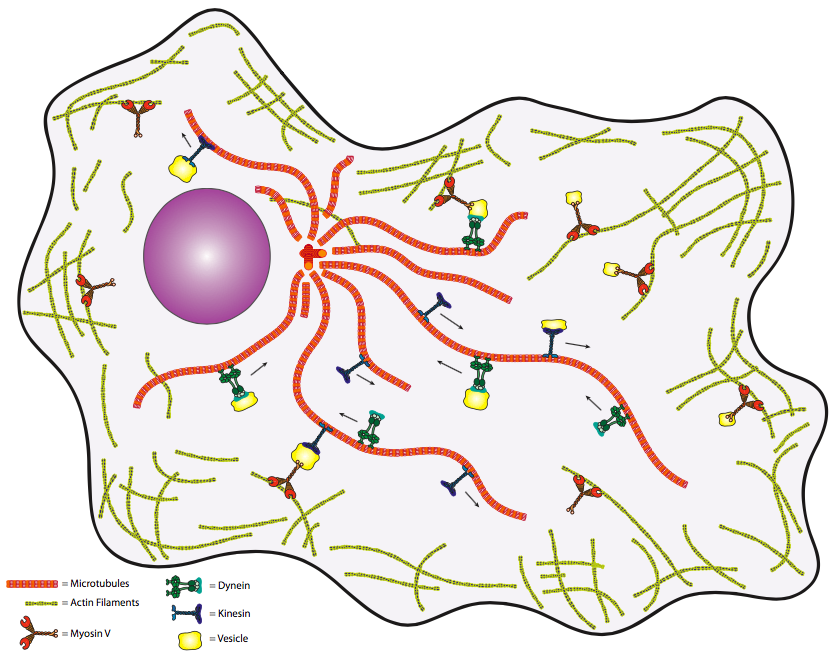
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)למרות שזה יכול להיות שימושי לחשוב על מבנים ציטו-שלד אלה כמקבילים לשלד של בעלי חיים, אולי דרך טובה יותר לזכור את המיקום היחסי של המיקרוטובולים והמיקרופילמנטים היא על ידי תפקידם בהובלת מטען תוך תאי מחלק אחד של התא לחלק אחר. לפי האנלוגיה הזו, אנו עשויים לשקול את המיקרוטובולים כמערכת פסי רכבת, בעוד שהמיקרופילמנטים דומים יותר לרחובות. באותה אנלוגיה, אנו יכולים להציע שרשת המיקרוטובוליות ורשת המיקרופילמנט מחוברים בנקודות מסוימות כך שכאשר המטען מגיע ליעדו הכללי באמצעות מיקרוטובולה (מסילה), ניתן לקחת אותו לכתובת הספציפית שלו באמצעות מיקרופילמנט. בואו נרחיב את האנלוגיה הזו עוד קצת. אם המיקרוטובולים והמיקרופילמנטים הם המסילה והרחובות, אז מהן הרכבות והמשאיות? אה, שאלה נבונה, גראסהופר. על המיקרו-צינורות, "הרכבות" הן אחת משתי משפחות של מנועים מולקולריים: הקינסינים והדינינים.
אנו יכולים להכליל מעט ולומר שהקינסינים נוסעים לכיוון הקצה (+) (לכיוון פריפריה של התא) בעוד הדינינים הולכים לכיוון הקצה (-) (לכיוון ה- MTOC). במיקרופילמנטים של אקטין, המנועים המולקולריים הם חלבונים ממשפחת המיוזינים. בשלב זה, האנלוגיות מסתיימות, שכן תפקודם של מנועים מולקולריים אלה שונה מאוד מתנועה ברכבת או במשאית. לבסוף, אפשר להטיל ספק בצורך הביולוגי במערכת תחבורה כזו. שוב, אם נעשה אנלוגיה לתחבורה אנושית, נוכל לומר שהובלה באמצעות דיפוזיה פשוטה דומה לאנשים הנושאים חבילות באופן אקראי על התא. כלומר, המשלוחים יבוצעו בסופו של דבר, אך לא תרצו לסמוך על שיטה זו לחומרים קריטיים לזמן. לפיכך יש צורך במערכת מכוונת ומהירה כדי לשמור על תאים (במיוחד תאים אוקריוטיים גדולים יותר) בחיים.
למרות שסוג זה של הובלה מתרחש בכל התאים האוקריוטיים, מקרה שנחקר במיוחד הוא הובלה אקסונלית (נקראת גם הובלה אקסופלסמית) בנוירונים. כאן, הובלת חומרים מגוף התא (סומה) לקצות האקסונים יכולה לפעמים לחצות מרחקים ארוכים מאוד עד כמה מטרים בבעלי חיים גדולים יותר, ועליה לעשות זאת בזמן. הובלה אקסונלית מסווגת בדרך כלל כאנטרוגרד (מסומה למסוף האקסון) או רטרוגרדית (מהטרמינלים לאחור). סוגי החומרים המועברים בשני הכיוונים הללו שונים מאוד: חלק ניכר מההובלה האנטרוגרדית היא אבני בניין חלבוניות להארכת האקסון או שלפוחיות סינפטיות המכילות נוירוטרנסמיטורים; הובלה רטרוגרדית היא בעיקר שלפוחיות אנדוציטיות ומולקולות איתות. הובלה אקסונלית מסווגת גם כמהירה ואיטית. הובלה איטית היא בעיקר תנועה של חלבונים הקשורים ישירות למנועים, והם יכולים לנוע מ-100 מ"מ ליום (ScA, רכיב איטי א) עד 3 מ"מ/יום (ScB). לשם השוואה, הובלה מהירה היא בדרך כלל תנועה של שלפוחית, ויכולה לנוע בין 50 ל -400 מ"מ ליום. מנגנון ההובלה האיטית נדון במשך למעלה מעשור עד שנת 2000, כאשר הדמיה ישירה של נוירופילמנטים מסומנים פלואורסצנטית בהובלה הראתה שהתנועה בפועל של החלבונים דומה מאוד לתנועה בהובלה אקסונלית מהירה, אך היו הפסקות רבות בהובלה, מנגנון "עצור והלך" במקום לנוע ממקור ליעד ברציפות.
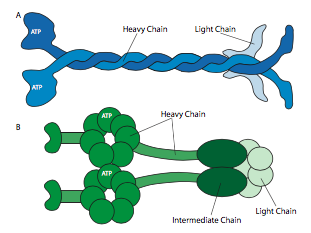
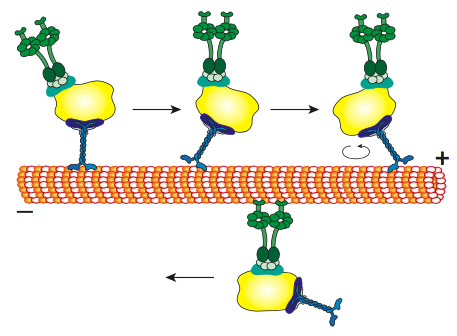
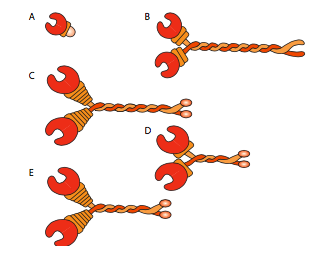
לכל הקינסינים והדינינים יש כמה מאפיינים משותפים מרכזיים. יש "ראש" משחרר אנרגיה קטליטי המחובר לציר או לאזור צוואר המאפשר למולקולה להתגמש או "לצעוד", ויש זנב נושא מטען מעבר לזה (איור). \(\PageIndex{8}\) ראש קינסין או דינין מזרז את ההידרוליזה של ATP, משחרר אנרגיה כדי לשנות את הקונפורמציה שלו ביחס לצוואר ולזנב של המולקולה, ומאפשר לה לשחרר זמנית את אחיזתה במיקרוטובולה, לסובב את "הירכיים" שלה סביב לשתול את עצמה "צעד" משם, ולהיקשר מחדש למיקרוטובולה (איור). \(\PageIndex{9}\) על מיקרופילמנטים של אקטין, המיוזינים, שישנם גם סוגים רבים (חלקם מתוארים באיור\(\PageIndex{10}\)) הם המנועים המולקולריים. התנועה שלהם שונה מדינינים וקינסינים, כפי שיתואר בסעיף הבא, אך משתמשת גם באנרגיה של הידרוליזה של ATP כדי לספק אנרגיה לשינויים הקונפורמטיביים הדרושים לתנועה. הצגנו את המנועים, אך בהתחשב במגוון העצום במולקולות שצריך להעביר סביב תא, זה יהיה בלתי אפשרי עבור המנועים להיקשר ישירות לכולם. למעשה, המנועים נקשרים למטען שלהם באמצעות מולקולות מתאם הקושרות את המנוע בצד אחד, ומולקולת מטען או שלפוחית מצד שני. בדיקה נוספת של המטען וניתוב המטען לפי סמני כתובות (SNAREs) נדונה בפרק ההובלה שלפוחית.