
12.2: חוטי ביניים
- Page ID
- 208454

"חוטי ביניים" הוא למעשה שם גנרי למשפחת חלבונים (מקובצים ל -6 מחלקות המבוססות על רצף ומבנה ביוכימי) המשרתים פונקציות דומות בהגנה ועיצוב התא או מרכיביו. מעניין, הם יכולים אפילו להימצא בתוך הגרעין. הפילאמינים הגרעיניים, המהווים חוטי ביניים מסוג V, יוצרים רשת הגנה חזקה המחוברת לפנים הפנימיות של הממברנה הגרעינית. לנוירונים יש נוירופילמנטים (מחלקה IV), המסייעים לספק מבנה לאקסונים - הרחבות ארוכות, דקות ועדינות של התא שיכולות לרוץ באורך מטרים בבעלי חיים גדולים. לתאי העור יש ריכוז גבוה של קרטין (מחלקה I), שלא רק עובר בתא, אלא מתחבר כמעט ישירות לסיבי הקרטין של תאים שכנים באמצעות סוג של מבנה הידבקות תאי הנקרא דסמוזום (מתואר בפרק הבא). זה מאפשר ללחץ שעשוי לפרוץ תא בודד להתפשט על פני תאים רבים, לחלוק את הנטל ובכך להגן על כל חבר. למעשה, מומים של קרטינים או של החלבונים היוצרים את הדסמוזומים יכולים להוביל למצבים המכונים ביחד epidermolysis bullosa, שבהם העור שביר בצורה יוצאת דופן, שלפוחית ומתפרק במגע קל בלבד, תוך פגיעה בקו ההגנה הראשון של המטופל מפני זיהום.
רוב חוטי הביניים נופלים בין 50-100 kDa, כולל קרטינים (40-67 kDa), למינים (60-70 kDa) ונוירופילמנטים (62-110 kDa). נסטין (מחלקה VI), המצוי בעיקר בנוירונים, הוא יוצא מן הכלל, בערך 240 kDa.
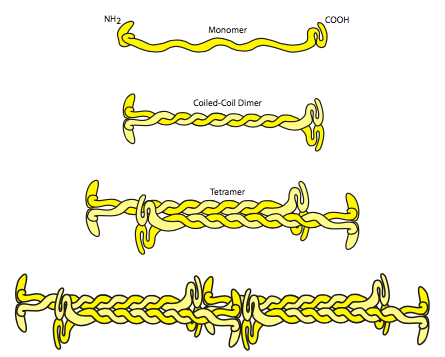
מבחינה מבנית, כפי שהוזכר קודם לכן, כל קינות הביניים מתחילות מתת-יחידה סיבית (איור). \(\PageIndex{2}\) לאחר מכן זה מתפתל סביב יחידת משנה חוטית אחרת ליצירת דימר סליל מפותל, או פרוטופילמנט. פרוטופילמנטים אלה מקיימים אינטראקציה ליצירת טטרמרים, הנחשבים ליחידה הבסיסית של בניית נימה ביניים. באמצעות חלבונים הנקראים פלקטינים, ניתן לחבר את קינות הביניים זו לזו ליצירת יריעות ורשתות. פלקטינים יכולים גם לחבר את קינות הביניים לחלקים אחרים של השלד, בעוד שחלבונים אחרים יכולים לעזור לחבר את שלד הציטוס IF לקרום התא (למשל desmoplakin). המאפיין הבולט ביותר של חוטי ביניים הוא אורך החיים היחסי שלהם. ברגע שהם נעשים, הם משתנים ונעים לאט מאוד. הם יציבים מאוד ולא נשברים בקלות. הם בדרך כלל אינם אינרטיים לחלוטין, אך בהשוואה למיקרו-צינורות ומיקרופילמנטים, נראה שהם לפעמים.
אפידרמוליזה בולוזה סימפלקס היא אוסף של מחלות מולדות הנגרמות על ידי מוטציות בגנים הקרטין KRT5 או KRT14, או לגן הפלקטין PLEC1. מוטציות אלה מחלישות את הפילמור של קרטין לחוטים, או את האינטראקציה בין חוטי קרטין. זה מוביל לחוסר היכולת של כל תא בודד לשמור על שלמות מבנית בלחץ. סוג אחר של EB, בולוזה אפידרמוליזה צומתית (JEB), נגרם על ידי מוטציות לקולטני אינטגרין (b4, a6) או למינינים. זה כולל JEB gravis או מחלת הרליץ, שהיא החמורה ביותר, המובילה לרוב למוות מוקדם לאחר הלידה. JEB קשור גם למחלות דיסטרופיות של אפידרמוליזה בולוזה (DEB) כגון קוקיין-טוריין, שכל אחת מהן נובעת ממוטציה בקולגן מסוג VII. תוצרי הגנים המעורבים ב-JEB ו-DEB נדונים ביתר פירוט בפרק הבא. הם ממלאים תפקיד בהדבקת התאים לקרום המרתף, ובלעדיהם, חוסר הארגון של התאים מוביל לקשרים לא שלמים בין תאי האפידרמיס, ולכן לקוי בשיתוף הלחץ.
צורות מסוימות של מחלת שארקו-מארי-שן, מחלת העצבים ההיקפית התורשתית הנפוצה ביותר, קשורות גם למוטציות של גנים של נימה ביניים. מחלה זו, הידועה גם בשם ניוון שרירים נקבי או נוירופתיה חושית מוטורית תורשתית, היא מחלה ניוונית לא קטלנית הפוגעת בעיקר בעצבי הזרועות והרגליים הדיסטליות. יש מגוון רחב של סוגים וסיבות CMT, כאשר הנפוצה ביותר היא מומים בתאי שוואן ומעטפת המיאלין שהם יוצרים. סוג CMT 2 מאופיין במומים באקסונים העצביים ההיקפיים, והוא קשור למוטציות של חלבוני למין A ושל נוירופילמנטים קלים. המנגנון הסיבתי טרם נקבע; עם זאת, הנוירופילמנטים הם מרכיבים משמעותיים בשמירה על שלמותם של אקסונים ארוכים.