
12.1: מבוא לשלד הציטוס
- Page ID
- 208459

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)כאשר תא אוקריוטי נלקח מהקשרו הפיזיולוגי ומונח בצלחת פטרי מפלסטיק או מזכוכית, נראה בדרך כלל שהוא משתטח במידה מסוימת. על תהום, זה היה מתנהג כמו שעון סלבדור דאלי, נוטף מעבר לקצה. ההנחה המיידית, במיוחד לאור העובדה שהתא ידוע כבעיקר מים במסה ובנפח, היא שהתא הוא פשוט שקית נוזלים. עם זאת, לתא יש למעשה מיקרו-מבנה מורכב בתוכו, הממוסגר באופן פנימי על ידי מרכיבי השלד.
למרות שהגנים אינם נשמרים במיוחד, שילוב של דמיון גנטי ומבנה חלבון אישר את נוכחותם של חלבונים פרוקריוטים הקשורים לחלבונים ציטוסקלטליים אוקריוטיים הן בצורה והן בתפקוד. בהשוואה לציטוס השלד האוקריוטי, מחקר של חלבונים פרוקריוטיים הוא עדכני מאוד, ובמשך זמן רב הייתה הנחה שלפרוקריוטים אין או זקוקים לארכיטקטורה ציטוסקלטלית. FtsZ, המקבילה החיידקית לטובולין, התגלה בשנת 1980 אך רוב העבודה עליו התרחשה בעשור האחרון. MreB הוא חלבון דמוי אקטין, בהשוואה לראשונה לאקטין בשנת 1992, וקרסנטין, חלבון מסוג נימה ביניים, תואר רק בשנת 2003. לסקירה מקיפה של חלבוני שלד פרוקריוטיים, ראה Graumann, P.L., Ann. הכומר מיקרוביולוגיה 61:589-618, 2007.
כפי שהשם מרמז, שלד הציטוס פועל בדומה לשלדים שלנו בתמיכה בצורתו הכללית של תא. בניגוד לשלדים שלנו, השלד הוא דינמי מאוד ותנועתי פנימי, משתנה ומתארגן מחדש בתגובה לצרכי התא. יש לו גם מגוון מטרות מעבר פשוט לספק את צורת התא. באופן כללי, ניתן לסווג אותם כמבנים ותחבורתיים. בעוד שכל שלושת המרכיבים העיקריים של השלד מבצעים כל אחת מהפונקציות הללו, הם אינם עושים זאת באופן שווה, מכיוון שהמאפיינים הביופיזיים שלהם שונים בתכלית. ביחס למבנה, בשלב כלשהו בחייו של כל תא, עליו לשנות צורה, בין אם פשוט גדל או יורד בגודלו, או שינוי דרסטי יותר כמו הצורה הסופר-מוארכת של נוירונים עם אקסונים, השלד הציטוס חייב להיות מסוגל להגיב על ידי הגדלה והקטנה דינמית של גודל המבנים הפנימיים לפי הצורך. המבנה חל גם על המיקום היחסי של יסודות תאיים פנימיים, כגון אברונים או חלבונים, זה לזה. בתאים רבים מאוד מיוחדים, ההפרדה של מבנים מסוימים בתוך חלקים מסוימים של התא חיונית לתפקודו. הובלה מתייחסת לתנועה של מולקולות ואברונים בתוך התא וכן לתנועה של התא בכללותו. זה עתה דנו בתנועה תוך תאית של חלבונים ושומנים באמצעות שלפוחיות בפרק האחרון. שלפוחיות אלה, כפי שנראה בפרק זה, אינן צפות רק ממקום למקום; הן נעות בכוונה ובכיוון לאורך השלד כמו מטען בכבישים מהירים או פסי רכבת. ביחס לתנועת תאים שלמים, זה יכול לנוע בין חתירה או שחייה על ידי אורגניזמים חד תאיים לזחילה סטריאוטיפית ומתואמת מאוד של תאים רבים מנקודת המוצא שלהם ליעדם בסופו של דבר במהלך התפתחות אורגניזם מטאזואי או תנועת פיברובלסטים לרפא חתך בעור שלך.
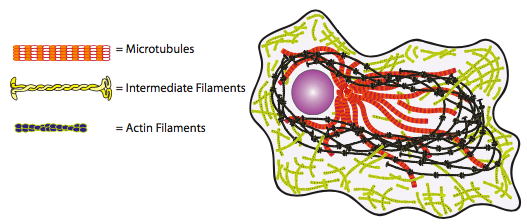
שלושת המרכיבים העיקריים של השלד הם מיקרוטובולים, מיקרופילמנטים וחוטי ביניים. כל אחד מאלה הוא פולימרים המורכבים מיחידות משנה חוזרות בסידורים ספציפיים. במבט מהיר בלבד (איור\(\PageIndex{1}\)), ברור מאוד שסביר להניח כי חוטי הביניים ימלאו תפקיד שונה באופן משמעותי ממיקרוטובולים או מיקרופילמנטים. מכיוון שה- IF עשויים מיחידות משנה סיביות ארוכות המתפתלות זו סביב זו ליצירת החוט, ברור שיש מגע רב (המאפשר יצירת קשרי מימן, המכונה וולקרו מולקולרי TM) בין יחידות משנה המספקות חוזק מתיחה רב. קשה מאוד לפרק את יחידות המשנה הללו, ולכן ה- IF משמשים בעיקר למטרות נושאות עומס ארוכות טווח או קבועות. כשמסתכלים על שני המרכיבים האחרים של השלד, ניתן לראות שעם הצורה הכדורית במקום הסיבית של יחידות המשנה, שטח המגע המרבי בין יחידות המשנה מוגבל מאוד (חשבו על אזור המגע כשאתם דוחפים שני כדורי סל זה לזה), מה שמקל על הפרדת יחידות המשנה או שבירת המיקרופילמנט או המיקרוטובולה. התא יכול להשתמש במאפיין זה לטובתו, על ידי ניצול סוגים אלה של סיבי ציטושלד במצבים דינמיים שבהם היווצרות או הרס של חוטי ביניים ייקח הרבה יותר מדי זמן. כעת אנו מתייחסים לשלוש הקבוצות הללו של אלמנטים ציטו-שלד בפירוט רב יותר.