
9.1: איך חיידקים גדלים
- Page ID
- 208996

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)מטרות למידה
- הגדירו את זמן הייצור לצמיחה על סמך ביקוע בינארי
- זהה ותאר את פעילותם של מיקרואורגניזמים העוברים שלבים אופייניים של ביקוע בינארי (חלוקת תאים פשוטה) בעקומת גדילה
- הסבר מספר שיטות מעבדה המשמשות לקביעת ספירת תאים בת קיימא וסך הכל באוכלוסיות העוברות צמיחה מעריכית
- תאר דוגמאות לחלוקת תאים שאינה כרוכה בביקוע בינארי, כגון ניצנים או פיצול
- תאר את היווצרותם ומאפייניהם של ביופילמים
- זהה סיכונים בריאותיים הקשורים לביופילמים וכיצד מטפלים בהם
- תאר חישת מניין ותפקידה בתקשורת בין תא לתא ותיאום פעילויות סלולריות
ג'ני, אישה בהריון בת 24 בשליש השני שלה, מבקרת במרפאה עם תלונות על חום גבוה, 38.9° C (102° F), עייפות וכאבי שרירים-סימנים ותסמינים אופייניים לשפעת. ג'ני מתאמנת באופן קבוע ועוקבת אחר תזונה מזינה בדגש על מזון אורגני, כולל חלב גולמי שהיא רוכשת משוק איכרים מקומי. כל החיסונים שלה מעודכנים. עם זאת, נותן שירותי הבריאות שרואה את ג'ני מודאג ומורה לשלוח דגימת דם לבדיקה על ידי המעבדה למיקרוביולוגיה.
תרגיל \(\PageIndex{1}\)
מדוע הרופא מודאג מהסימנים והתסמינים של ג'ני?
מחזור תאי החיידק כרוך ביצירת תאים חדשים באמצעות שכפול של DNA וחלוקת רכיבים תאיים לשני תאי בת. בפרוקריוטים, הרבייה היא תמיד א -מינית, אם כי מתרחשת רקומבינציה גנטית נרחבת בצורה של העברת גנים אופקית, כפי שייחקר בפרק אחר. לרוב החיידקים יש כרומוזום מעגלי יחיד; עם זאת, קיימים כמה יוצאים מן הכלל. לדוגמה, Borrelia burgdorferi, סוכן סיבתי של מחלת ליים, יש כרומוזום ליניארי.
ביקוע בינארי
המנגנון הנפוץ ביותר של שכפול תאים בחיידקים הוא תהליך הנקרא ביקוע בינארי, המתואר באיור\(\PageIndex{1}\):. לפני החלוקה, התא גדל ומגדיל את מספר הרכיבים הסלולריים שלו. לאחר מכן, שכפול ה-DNA מתחיל במיקום על הכרומוזום המעגלי הנקרא מקור השכפול, שבו הכרומוזום מחובר לקרום התא הפנימי. השכפול ממשיך בכיוונים מנוגדים לאורך הכרומוזום עד שמגיעים לקצה.
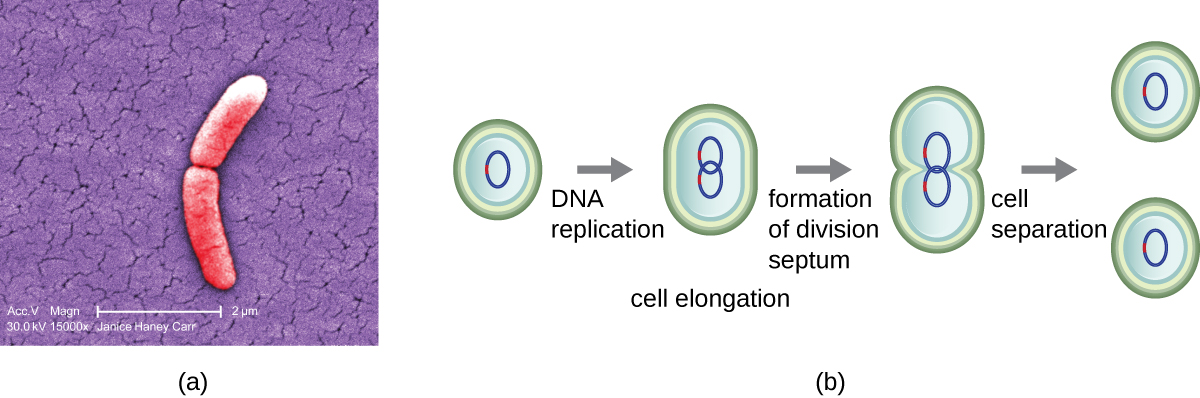
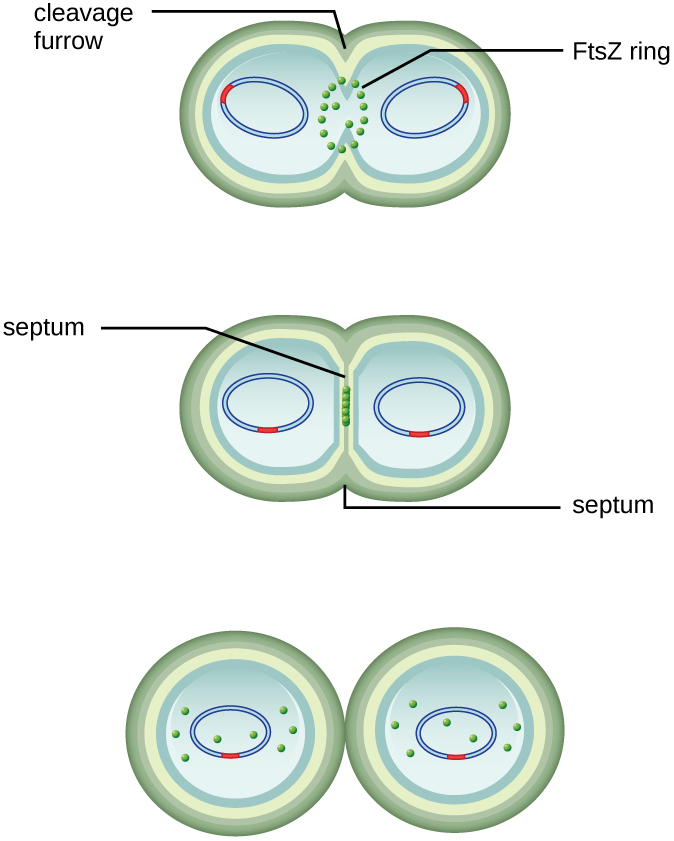
מרכז התא המוגדל מתכווץ עד שנוצרים שני תאי בת, כל צאצא מקבל עותק מלא של הגנום ההורי וחלוקה של הציטופלזמה (ציטוקינזיס). תהליך זה של ציטוקינזיס וחלוקת תאים מכוון על ידי חלבון הנקרא FtsZ. FtsZ מתאסף לטבעת Z על הממברנה הציטופלזמית (איור). \(\PageIndex{2}\) טבעת Z מעוגנת על ידי חלבונים קושרי FTSZ ומגדירה את מישור החלוקה בין שני תאי הבת. חלבונים נוספים הנדרשים לחלוקת תאים מתווספים לטבעת Z ליצירת מבנה הנקרא מחלק. החלוקה מופעלת כדי לייצר דופן תא פפטידוגליקן ולבנות מחיצה המחלקת את שני תאי הבת. תאי הבת מופרדים על ידי מחיצת החלוקה, שם יש לשפץ את כל השכבות החיצוניות של התאים (דופן התא והקרומים החיצוניים, אם קיימים) לחלוקה מלאה. לדוגמה, אנו יודעים שאנזימים ספציפיים שוברים קשרים בין המונומרים בפפטידוגליקנים ומאפשרים הוספת יחידות משנה חדשות לאורך מחיצת החלוקה.
תרגיל \(\PageIndex{2}\)
מה שמו של החלבון שמתאסף לטבעת Z כדי ליזום ציטוקינזיס וחלוקת תאים?
זמן דור
באורגניזמים אוקריוטיים, זמן הדור הוא הזמן בין אותן נקודות של מחזור החיים בשני דורות רצופים. לדוגמה, זמן הדור האופייני לאוכלוסיית האדם הוא 25 שנה. הגדרה זו אינה מעשית לחיידקים, שעלולים להתרבות במהירות או להישאר רדומים במשך אלפי שנים. בפרוקריוטים (Bacteria and Archaea), זמן הדור נקרא גם זמן ההכפלה ומוגדר כזמן שלוקח לאוכלוסייה להכפיל את עצמה באמצעות סיבוב אחד של ביקוע בינארי. זמני ההכפלה של חיידקים משתנים מאוד. בעוד ש - Escherichia coli יכול להכפיל את עצמו תוך 20 דקות בלבד בתנאי גידול אופטימליים במעבדה, חיידקים מאותו המין עשויים להזדקק למספר ימים כדי להכפיל אותם בסביבות קשות במיוחד. רוב הפתוגנים גדלים במהירות, כמו E. coli, אבל יש יוצאים מן הכלל. לדוגמה, שחפת Mycobacterium, סוכן סיבתי של שחפת, יש זמן דור של בין 15 ל 20 שעות. מצד שני, M. leprae, הגורם למחלת הנסן (צרעת), גדל לאט הרבה יותר, עם זמן הכפלה של 14 יום.
חישוב מספר התאים
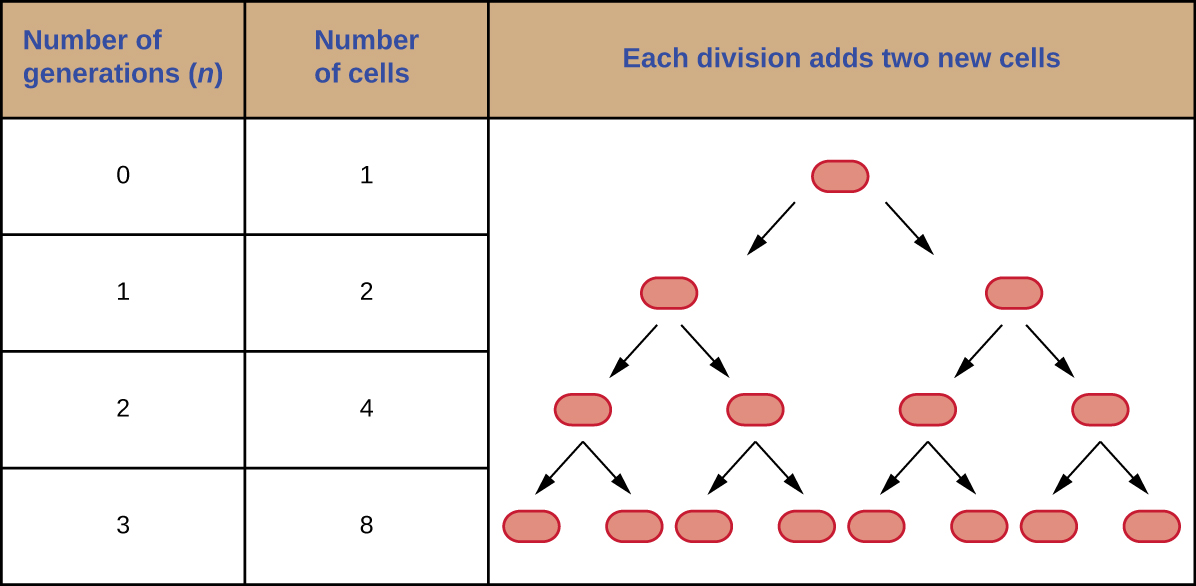
ניתן לחזות את מספר התאים באוכלוסייה כאשר הם מתחלקים בביקוע בינארי בקצב קבוע. כדוגמה, שקול מה קורה אם תא בודד מתחלק כל 30 דקות למשך 24 שעות. התרשים באיור \(\PageIndex{3}\) מציג את הגידול במספר התאים בשלושת הדורות הראשונים.
מספר התאים גדל באופן אקספוננציאלי ויכול לבוא לידי ביטוי כ - 2 n, כאשר n הוא מספר הדורות. אם התאים מתחלקים כל 30 דקות, לאחר 24 שעות, היו מתרחשות 48 חלוקות. אם ניישם את הנוסחה 2 n, כאשר n שווה ל 48, התא הבודד יוליד 2 48 או 281,474,976,710,656 תאים ב 48 דורות (24 שעות). כאשר מתמודדים עם מספרים עצומים כאלה, זה מעשי יותר להשתמש בסימון מדעי. לכן, אנו מבטאים את מספר התאים כ- 2.8 × 10 14 תאים.
בדוגמה שלנו, השתמשנו בתא אחד כמספר התאים הראשוני. עבור כל מספר של תאים מתחילים, הנוסחה מותאמת באופן הבא:
\[N_n = N_02^n\]
N n הוא מספר התאים בכל דור n, N 0 הוא מספר התאים הראשוני, ו - n הוא מספר הדורות.
תרגיל \(\PageIndex{3}\)
עם זמן הכפלה של 30 דקות וגודל אוכלוסייה התחלתי של 1 × 10 5 תאים, כמה תאים יהיו נוכחים לאחר שעתיים, בהנחה שאין מוות של תאים?
עקומת הצמיחה
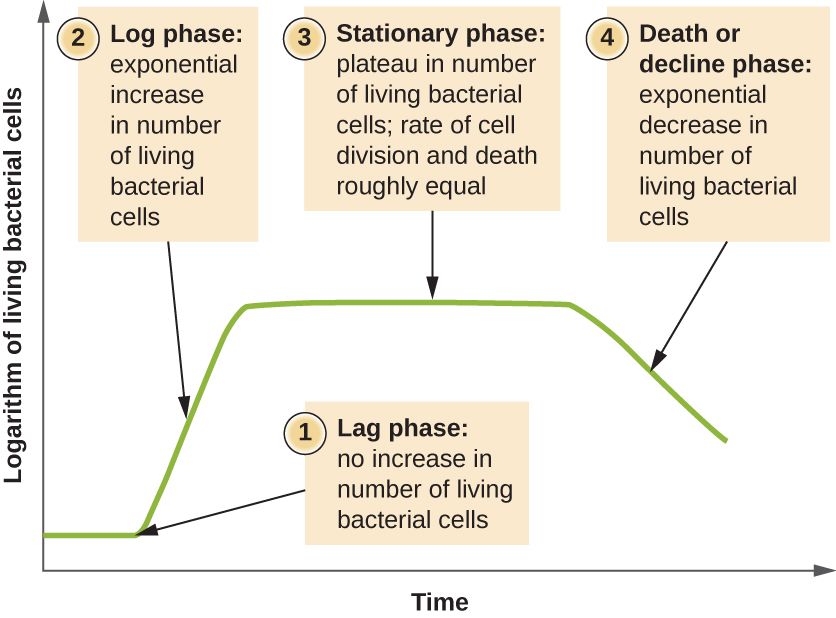
מיקרואורגניזמים הגדלים בתרבית סגורה (הידועה גם כתרבות אצווה), שבה לא מוסיפים חומרים מזינים ורוב הפסולת אינה מוסרת, עוקבים אחר דפוס גדילה הניתן לשחזור המכונה עקומת הצמיחה. דוגמה לתרבות אצווה בטבע היא בריכה שבה מספר קטן של תאים גדלים בסביבה סגורה. צפיפות התרבות מוגדרת כמספר התאים ליחידת נפח. בסביבה סגורה, צפיפות התרבות היא גם מדד למספר התאים באוכלוסייה. זיהומים בגוף לא תמיד עוקבים אחר עקומת הגדילה, אך מתאמים יכולים להתקיים בהתאם לאתר וסוג הזיהום. כאשר מספר התאים החיים מתווה כנגד הזמן, ניתן לראות שלבים נפרדים בעקומה (איור\(\PageIndex{4}\)).
שלב הפיגור
תחילת עקומת הגידול מייצגת מספר קטן של תאים, המכונים חיסון, המתווספים למדיום תרבות טרי, מרק תזונתי התומך בצמיחה. השלב הראשוני של עקומת הצמיחה נקרא שלב השהיה, שבמהלכו התאים מתכוננים לשלב הבא של הצמיחה. מספר התאים אינו משתנה בשלב הפיגור; עם זאת, התאים גדלים ופעילים מטבולית, ומסנתזים חלבונים הדרושים לצמיחה בתוך המדיום. אם תאים כלשהם נפגעו או נדהמו במהלך ההעברה למדיום החדש, התיקון מתרחש בשלב הפיגור. משך שלב הפיגור נקבע על ידי גורמים רבים, כולל המינים וההרכב הגנטי של התאים, הרכב המדיום וגודל החיסון המקורי.
שלב היומן
בשלב הצמיחה הלוגריתמי (יומן), הנקרא לפעמים שלב צמיחה מעריכי, התאים מתחלקים באופן פעיל על ידי ביקוע בינארי ומספרם גדל באופן אקספוננציאלי. עבור כל מין חיידקי נתון, זמן הייצור בתנאי גידול ספציפיים (חומרים מזינים, טמפרטורה, pH וכן הלאה) נקבע גנטית, וזמן הדור הזה נקרא קצב הגדילה הפנימי. במהלך שלב היומן, הקשר בין זמן למספר התאים אינו ליניארי אלא מעריכי; עם זאת, עקומת הצמיחה משורטטת לעתים קרובות על גרף סמילוגריתמי, כפי שמוצג באיור\(\PageIndex{5}\), המעניק מראה של קשר ליניארי.
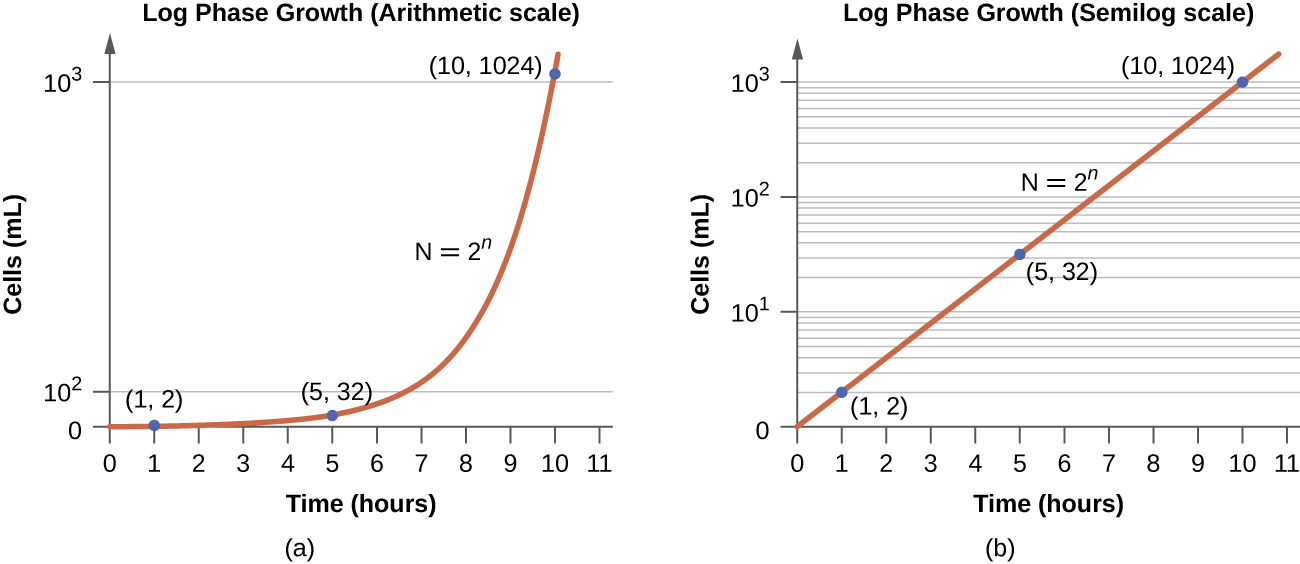
תאים בשלב היומן מראים קצב גדילה קבוע ופעילות מטבולית אחידה. מסיבה זו, תאים בשלב היומן משמשים באופן מועדף ליישומים תעשייתיים ועבודות מחקר. שלב היומן הוא גם השלב שבו החיידקים הם הרגישים ביותר לפעולה של חומרי חיטוי ואנטיביוטיקה נפוצה המשפיעים על סינתזת חלבון, DNA ודופן התא.
שלב נייח
ככל שמספר התאים גדל במהלך שלב היומן, מספר גורמים תורמים להאטת קצב הגדילה. מוצרי פסולת מצטברים וחומרים מזינים מנוצלים בהדרגה. בנוסף, דלדול הדרגתי של חמצן מתחיל להגביל את צמיחת התאים האירוביים. שילוב זה של תנאים שליליים מאט ולבסוף מעכב את גידול האוכלוסייה. המספר הכולל של תאים חיים מגיע לרמה המכונה השלב הנייח (איור\(\PageIndex{4}\)). בשלב זה, מספר התאים החדשים שנוצרו על ידי חלוקת תאים שווה כעת למספר התאים הגוססים; לפיכך, אוכלוסיית התאים החיים הכוללת עומדת יחסית. צפיפות התרבות בתרבות נייחת קבועה. כושר הנשיאה של התרבות, או צפיפות התרבות המרבית, תלוי בסוגי המיקרואורגניזמים בתרבית ובתנאים הספציפיים של התרבות; עם זאת, כושר הנשיאה קבוע לאורגניזם נתון שגדל באותם תנאים.
במהלך השלב הנייח, התאים עוברים למצב הישרדותי של חילוף החומרים. ככל שהצמיחה מאטה, כך גם הסינתזה של פפטידוגליקנים, חלבונים וחומצות גרעין; לפיכך, תרבויות נייחות פחות רגישות לאנטיביוטיקה המשבשת תהליכים אלה. בחיידקים המסוגלים לייצר אנדוספורים, תאים רבים עוברים ספורולציה בשלב הנייח. מטבוליטים משניים, כולל אנטיביוטיקה, מסונתזים בשלב הנייח. בחיידקים פתוגניים מסוימים, השלב הנייח קשור גם לביטוי של גורמי ארסיות, מוצרים התורמים ליכולתו של חיידק לשרוד, להתרבות ולגרום למחלות באורגניזם מארח. לדוגמה, חישת מניין ב Staphylococcus aureus יוזמת ייצור אנזימים שיכולים לפרק רקמות אנושיות ופסולת תאית, ומפנה את הדרך לחיידקים להתפשט לרקמה חדשה שבה חומרים מזינים נמצאים בשפע יותר.
שלב המוות
כאשר מדיום תרבות צובר פסולת רעילה וחומרים מזינים מותשים, התאים מתים במספרים גדולים יותר ויותר. בקרוב, מספר התאים הגוססים עולה על מספר התאים המתחלקים, מה שמוביל לירידה מעריכית במספר התאים (איור\(\PageIndex{4}\)). זהו שלב המוות הנקרא כראוי, המכונה לפעמים שלב הירידה. תאים רבים מחליקים ומשחררים חומרים מזינים לתוך המדיום, ומאפשרים לתאים ששרדו לשמור על כדאיות וליצור אנדוספורים. כמה תאים, מה שנקרא persisters, מאופיינים בקצב חילוף חומרים איטי. תאים מתמידים חשובים מבחינה רפואית מכיוון שהם קשורים לזיהומים כרוניים מסוימים, כמו שחפת, שאינם מגיבים לטיפול אנטיביוטי.
שמירה על צמיחה מיקרוביאלית
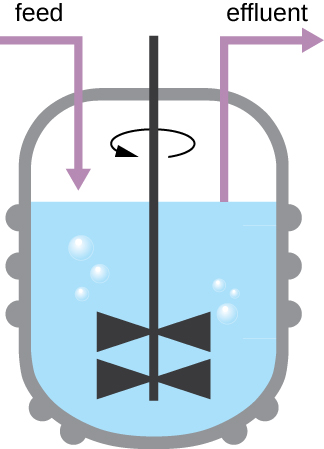
דפוס הצמיחה המוצג באיור \(\PageIndex{4}\) מתרחש בסביבה סגורה; חומרים מזינים אינם מתווספים ופסולת ותאים מתים אינם מוסרים. עם זאת, במקרים רבים, יתרון לשמור על תאים בשלב הלוגריתמי של הצמיחה. דוגמה אחת היא בתעשיות הקוצרות מוצרים מיקרוביאליים. כימוסטט (איור\(\PageIndex{6}\)) משמש לשמירה על תרבית רציפה שבה חומרים מזינים מסופקים בקצב קבוע. כמות מבוקרת של אוויר מעורבת בתהליכים אירוביים. תרחיף חיידקי מוסר באותו קצב שבו חומרים מזינים זורמים פנימה כדי לשמור על סביבת צמיחה אופטימלית.
תרגיל \(\PageIndex{4}\)
- באיזה שלב מתרחשת הצמיחה בקצב המהיר ביותר?
- ציין שני גורמים המגבילים את צמיחת החיידקים.
מדידת צמיחת חיידקים
הערכת מספר תאי החיידקים בדגימה, המכונה ספירת חיידקים, היא משימה נפוצה המבוצעת על ידי מיקרוביולוגים. מספר החיידקים במדגם קליני משמש אינדיקציה להיקף הזיהום. בקרת איכות של מי שתייה, מזון, תרופות ואפילו קוסמטיקה מסתמכת על הערכות של ספירת חיידקים כדי לזהות זיהום ולמנוע את התפשטות המחלה. שתי גישות עיקריות משמשות למדידת מספר התא. השיטות הישירות כוללות ספירת תאים, בעוד שהשיטות העקיפות תלויות במדידת נוכחות או פעילות התא מבלי לספור תאים בודדים בפועל. לשיטות ישירות ועקיפות יש יתרונות וחסרונות ליישומים ספציפיים.
ספירת תאים ישירה
ספירת תאים ישירה מתייחסת לספירת התאים בתרבית נוזלית או מושבות על צלחת. זוהי דרך ישירה להעריך כמה אורגניזמים קיימים במדגם. בואו נסתכל תחילה על שיטה פשוטה ומהירה הדורשת רק שקופית מיוחדת ומיקרוסקופ מורכב.
הדרך הפשוטה ביותר לספור חיידקים נקראת ספירת תאים מיקרוסקופית ישירה, הכוללת העברת נפח ידוע של תרבית לשקופית מכוילת וספירת התאים תחת מיקרוסקופ אור. השקופית המכוילת נקראת תא פטרוף-האוסר (איור\(\PageIndex{7}\)) והיא דומה להמוציטומטר המשמש לספירת תאי דם אדומים. האזור המרכזי של תא הספירה נחרט לריבועים בגדלים שונים. דגימה של השעיית התרבות מתווספת לתא מתחת לכיסוי הממוקם בגובה מסוים מפני השטח של הרשת. ניתן להעריך את ריכוז התאים במדגם המקורי על ידי ספירת תאים בודדים במספר ריבועים וקביעת נפח המדגם שנצפה. שטח הריבועים והגובה שבו ממוקם הכיסוי מוגדרים עבור החדר. הריכוז חייב להיות מתוקן לדילול אם המדגם היה מדולל לפני הספירה.
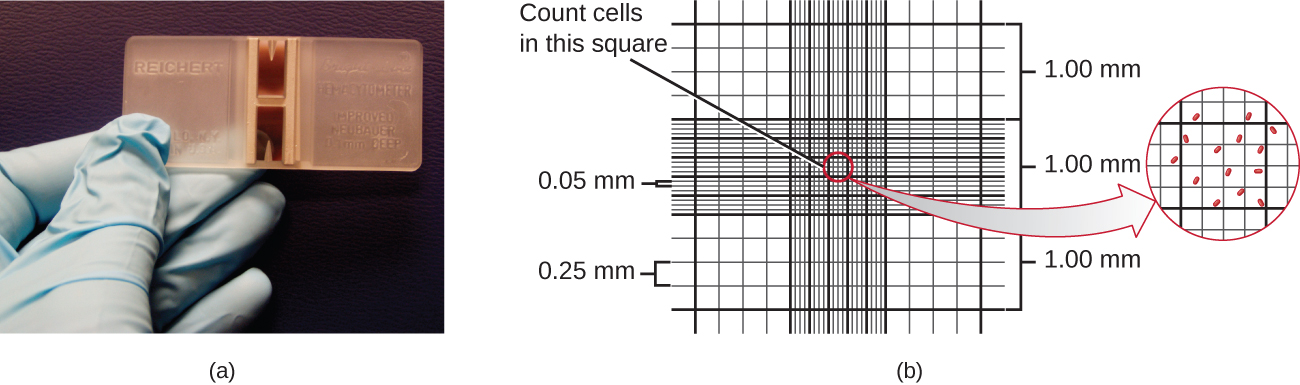
יש לספור תאים בכמה ריבועים קטנים ולקחת את הממוצע כדי לקבל מדידה אמינה. היתרונות של החדר הם שהשיטה קלה לשימוש, מהירה יחסית וזולה. בצד החיסרון, תא הספירה לא עובד טוב עם תרבויות מדוללות מכיוון שאולי אין מספיק תאים לספור.
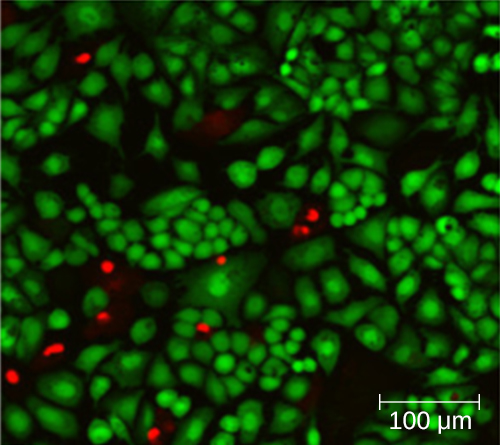
שימוש בתא ספירה אינו מניב בהכרח ספירה מדויקת של מספר התאים החיים מכיוון שלא תמיד ניתן להבחין בין תאים חיים, תאים מתים ופסולת באותו גודל מתחת למיקרוסקופ. עם זאת, טכניקות צביעת פלואורסצנטיות שפותחו לאחרונה מאפשרות להבחין בין חיידקים ברי קיימא ומתים. כתמי כדאיות אלה (או כתמים חיים) נקשרים לחומצות גרעין, אך הכתמים הראשוניים והמשניים שונים ביכולתם לחצות את הממברנה הציטופלזמית. הכתם הראשוני, המאיר ירוק, יכול לחדור לממברנות ציטופלזמיות שלמות, ולהכתים תאים חיים ומתים כאחד. הכתם המשני, המאיר אדום, יכול להכתים תא רק אם הממברנה הציטופלסמית ניזוקה במידה ניכרת. לפיכך, תאים חיים זורחים בירוק מכיוון שהם סופגים רק את הכתם הירוק, בעוד שתאים מתים נראים אדומים מכיוון שהכתם האדום מחליף את הכתם הירוק על חומצות הגרעין שלהם (איור\(\PageIndex{8}\)).
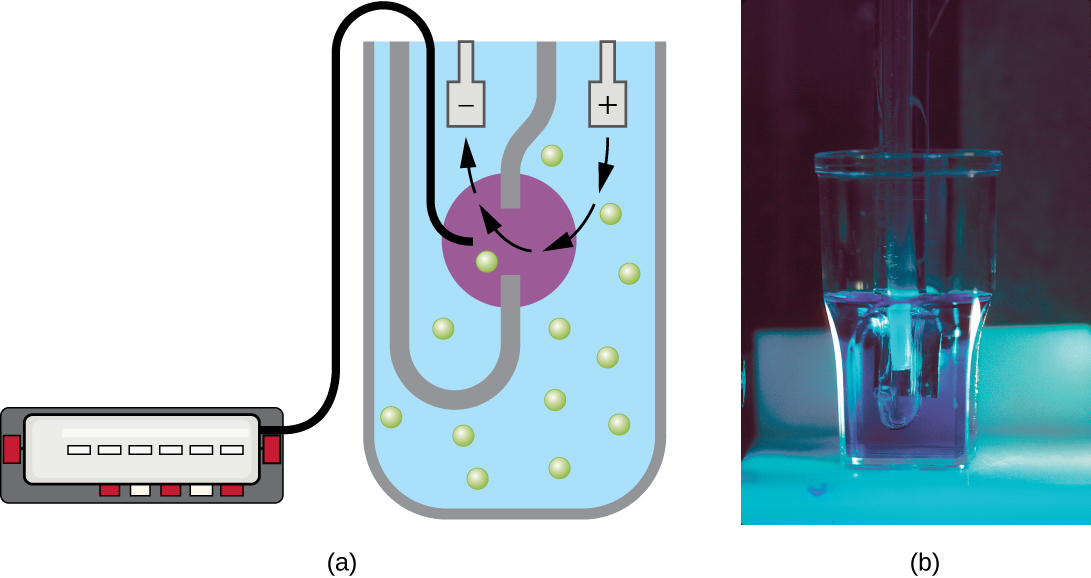
טכניקה נוספת משתמשת במכשיר ספירת תאים אלקטרוני (מונה קולטר) כדי לזהות ולספור את השינויים בהתנגדות החשמלית בתמיסת מלח. צינור זכוכית עם פתח קטן טובל בתמיסת אלקטרוליט. אלקטרודה ראשונה מושעה בצינור הזכוכית. אלקטרודה שנייה ממוקמת מחוץ לצינור. כאשר תאים נמשכים דרך הצמצם הקטן בצינור הזכוכית, הם משנים בקצרה את ההתנגדות הנמדדת בין שתי האלקטרודות והשינוי נרשם על ידי חיישן אלקטרוני (איור\(\PageIndex{9}\)); כל שינוי התנגדות מייצג תא. השיטה מהירה ומדויקת בטווח ריכוזים; עם זאת, אם התרבות מרוכזת מדי, יותר מתא אחד עשוי לעבור דרך הצמצם בכל זמן נתון ולהטות את התוצאות. שיטה זו גם אינה מבדילה בין תאים חיים ומתים.
ספירות ישירות מספקות אומדן של המספר הכולל של תאים במדגם. עם זאת, במצבים רבים, חשוב לדעת את מספר התאים החיים, או בת קיימא. יש צורך בספירות של תאים חיים בעת הערכת היקף הזיהום, יעילותן של תרכובות אנטי-מיקרוביאליות ותרופות, או זיהום מזון ומים.
תרגיל \(\PageIndex{5}\)
- מדוע שתספור את מספר התאים ביותר מריבוע אחד בתא פטרוף-האוסר כדי להעריך את מספרי התאים?
- בשיטת צביעת הכדאיות, מדוע תאים מתים נראים אדומים?
ספירת צלחות
ספירת הלוחות בת קיימא, או פשוט ספירת הצלחות, היא ספירה של תאים ברי קיימא או חיים. הוא מבוסס על העיקרון שתאים ברי קיימא משכפלים ומולידים מושבות גלויות כאשר הם מודגרים בתנאים מתאימים לדגימה. התוצאות מתבטאות בדרך כלל כיחידות יוצרות מושבה למיליליטר (CFU/mL) ולא כתאים למיליליטר מכיוון שיותר מתא אחד נחת באותה נקודה כדי להוליד מושבה אחת. יתר על כן, דגימות של חיידקים הגדלים באשכולות או בשרשראות קשות לפיזור ומושבה אחת עשויה לייצג מספר תאים. חלק מהתאים מתוארים כברי קיימא אך לא ניתנים לתרבות ולא יהוו מושבות על מדיה מוצקה. מכל הסיבות הללו, ספירת הלוחות בת קיימא נחשבת לאומדן נמוך של מספר התאים החיים בפועל. מגבלות אלה אינן גורעות מהתועלת של השיטה, המספקת הערכות של מספר חיידקים חיים.
מיקרוביולוגים סופרים בדרך כלל צלחות עם 30-300 מושבות. דוגמאות עם מעט מדי מושבות 300 <30) do not give statistically reliable numbers, and overcrowded plates (> מושבות) מקשות על ספירה מדויקת של מושבות בודדות. כמו כן, ספירות בטווח זה ממזערות התרחשויות של יותר מתא חיידקי אחד היוצר מושבה אחת. לפיכך, ה- CFU המחושב קרוב יותר למספר האמיתי של חיידקים חיים באוכלוסייה.
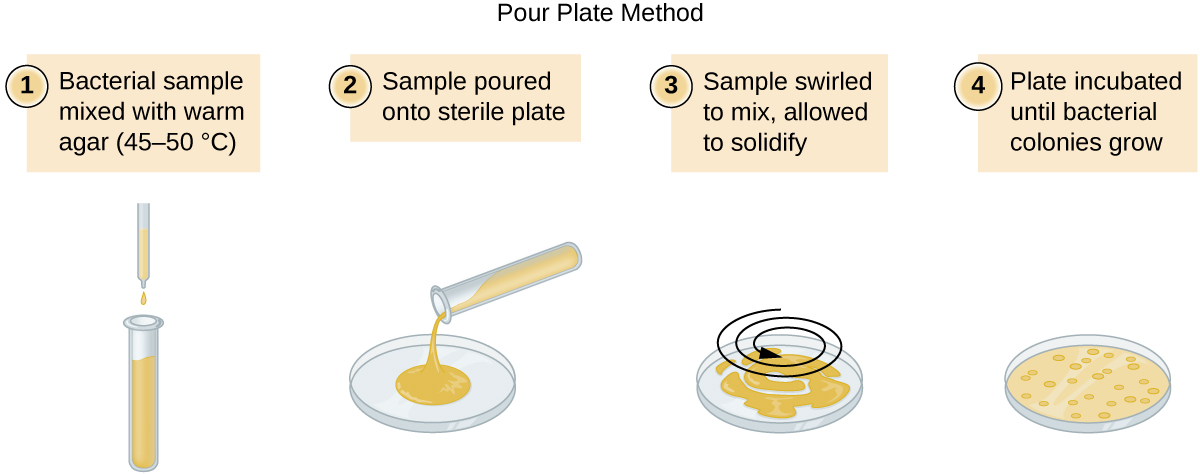
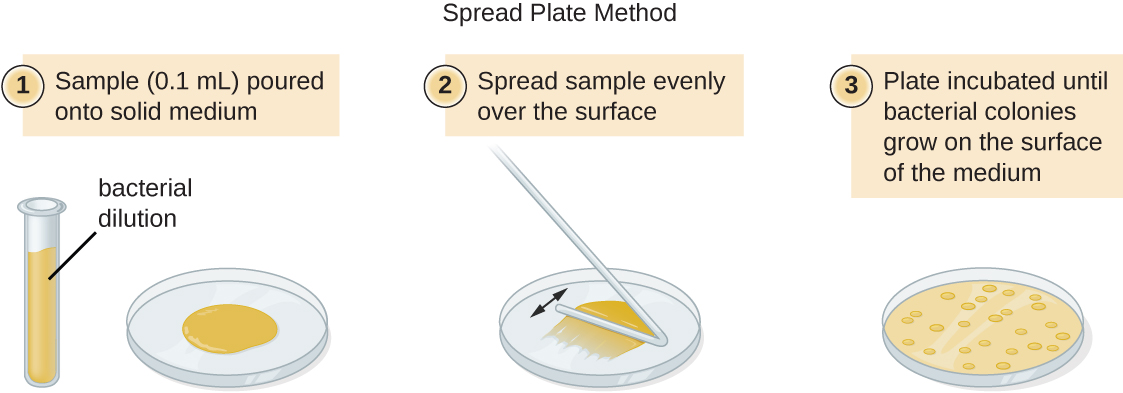
ישנן שתי גישות נפוצות לחיסון צלחות לספירות קיימא: צלחת המזיגה ושיטות צלחת הממרח. למרות שתהליך החיסון הסופי שונה בין שתי השיטות הללו, שתיהן מתחילות בדילול סדרתי של התרבות.
דילול סדרתי
הדילול הסדרתי של תרבות הוא צעד ראשון חשוב לפני שתמשיך לשיטת צלחת המזיגה או שיטת צלחת המריחה. מטרת תהליך הדילול הסדרתי היא להשיג צלחות עם CFUs בטווח של 30-300, והתהליך כולל בדרך כלל מספר דילולים בכפולות של 10 כדי לפשט את החישוב. מספר הדילולים הסדרתיים נבחר על פי הערכה ראשונית של צפיפות התרבות. איור \(\PageIndex{10}\) ממחיש את שיטת הדילול הסדרתי.
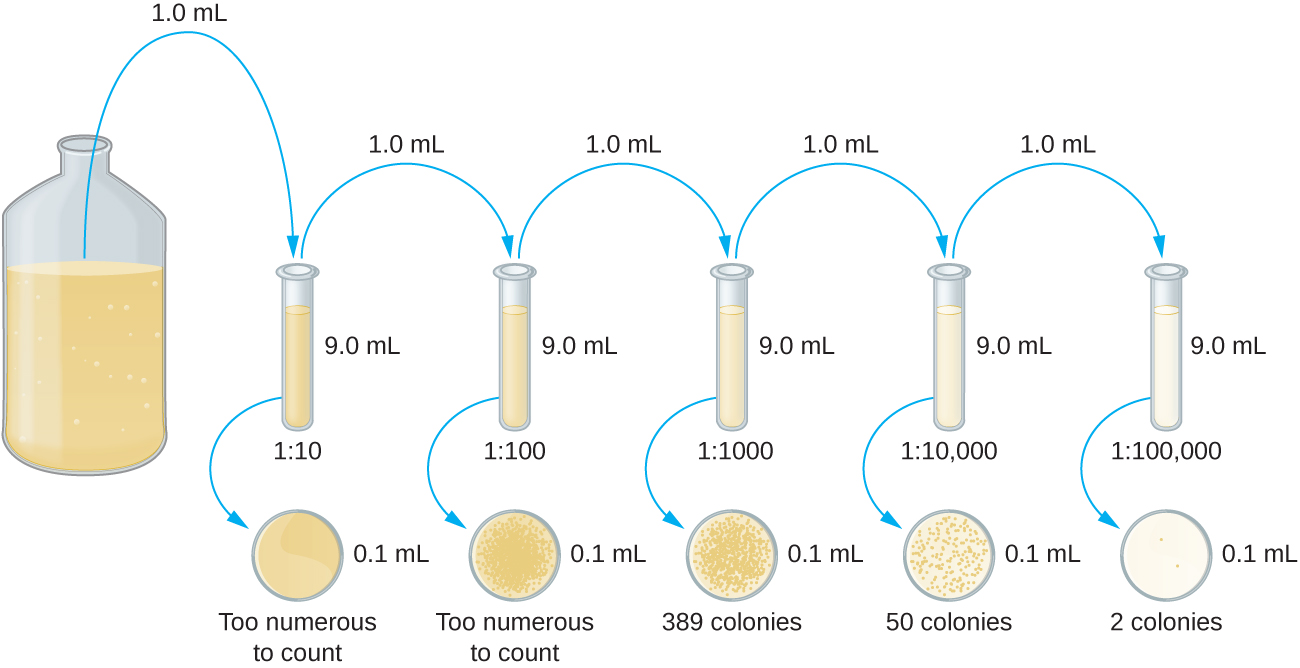
נפח קבוע של התרבות המקורית, 1.0 מ"ל, מתווסף ומעורבב היטב עם פתרון צינור הדילול הראשון, המכיל 9.0 מ"ל של מרק סטרילי. שלב זה מייצג גורם דילול של 10, או 1:10, בהשוואה לתרבות המקורית. מהדילול הראשון הזה, אותו נפח, 1.0 מ"ל, נסוג ומערבב עם צינור טרי של 9.0 מ"ל של תמיסת דילול. גורם הדילול הוא כעת 1:100 בהשוואה לתרבות המקורית. תהליך זה נמשך עד להפקת סדרה של דילולים שיסגרו את ריכוז התא הרצוי לספירה מדויקת. מכל צינור, מדגם מצופה על מדיום מוצק באמצעות שיטת צלחת לשפוך (איור\(\PageIndex{11}\)) או שיטת צלחת התפשטות (איור\(\PageIndex{12}\)). הלוחות מודגרים עד להופעת מושבות. בדרך כלל מכינים שתיים עד שלוש צלחות מכל דילול ומספרי המושבות הנספרות על כל צלחת ממוצעים. בכל המקרים, ערבוב יסודי של דגימות עם מדיום הדילול (כדי להבטיח את התפלגות התא בצינור הוא אקראי) הוא בעל חשיבות עליונה להשגת תוצאות אמינות.
גורם הדילול משמש לחישוב מספר התאים בתרבית התאים המקורית. בדוגמה שלנו, נספרו בממוצע 50 מושבות על הלוחות שהתקבלו מהדילול של 1:10,000. מכיוון שרק 0.1 מ"ל של השעיה הועברו על הצלחת, המכפיל הנדרש לשחזור הריכוז המקורי הוא 10 × 10,000. מספר CFU למ"ל שווה ל 50 × 100 × 10,000 = 5,000,000. מספר החיידקים בתרבית מוערך כ-5 מיליון תאים/מ"ל. ספירת המושבות שהתקבלה מהדילול של 1:1000 הייתה 389, הרבה מתחת ל -500 הצפויים להפרש פי 10 בדילולים. זה מדגיש את נושא אי הדיוק כאשר ספירת המושבות גדולה מ -300 ויותר מתא חיידקי אחד גדל למושבה אחת.
דגימה מדוללת מאוד - מי שתייה, למשל - עשויה שלא להכיל מספיק אורגניזמים כדי להשתמש באחת משיטות ספירת הלוחות המתוארות. במקרים כאלה, המדגם המקורי חייב להיות מרוכז ולא מדולל לפני ציפוי. ניתן להשיג זאת באמצעות שינוי של טכניקת ספירת הצלחות הנקראת טכניקת סינון הממברנה. נפחים ידועים מסוננים בוואקום באופן אספטי דרך קרום עם גודל נקבוביות קטן מספיק כדי ללכוד מיקרואורגניזמים. הממברנה מועברת לצלחת פטרי המכילה מצע גידול מתאים. מושבות נספרות לאחר הדגירה. חישוב צפיפות התא נעשה על ידי חלוקת ספירת התאים בנפח הנוזל המסונן.
צפו בסרטון זה להדגמות של דילולים סדרתיים וטכניקות צלחות פיזור.
המספר הסביר ביותר
מספר המיקרואורגניזמים בדגימות מדוללות בדרך כלל נמוך מכדי שניתן יהיה לזהות אותו בשיטות ספירת הלוחות שתוארו עד כה. עבור דגימות אלה, מיקרוביולוגים משתמשים באופן שגרתי בשיטת המספר הסביר ביותר (MPN), הליך סטטיסטי להערכת מספר המיקרואורגניזמים ברי קיימא במדגם. שיטת MPN, המשמשת לעתים קרובות לדגימות מים ומזון, מעריכה צמיחה ניתנת לזיהוי על ידי התבוננות בשינויים בעכירות או בצבע עקב פעילות מטבולית.
יישום טיפוסי של שיטת MPN הוא הערכת מספר הקוליפורמים במדגם של מי בריכה. קוליפורמים הם חיידקי מוט גרם שליליים המתסיסים לקטוז. נוכחות של קוליפורמים במים נחשב סימן של זיהום על ידי חומר צואה. עבור השיטה המוצגת באיור\(\PageIndex{13}\), סדרה של שלושה דילולים של דגימת המים נבדקת על ידי חיסון חמישה צינורות מרק לקטוז עם 10 מ"ל של מדגם, חמישה צינורות מרק לקטוז עם 1 מ"ל של מדגם, וחמישה צינורות מרק לקטוז עם 0.1 מ"ל של מדגם. צינורות מרק הלקטוז מכילים מחוון pH שמשנה את צבעו מאדומה לצהוב כאשר הלקטוז מותסס. לאחר חיסון ודגירה, הצינורות נבדקים לאינדיקציה לצמיחת קוליפורם על ידי שינוי צבע במדיה מאדומה לצהוב. קבוצת הצינורות הראשונה (מדגם 10 מ"ל) הראתה צמיחה בכל הצינורות; קבוצת הצינורות השנייה (1 מ"ל) הראתה צמיחה בשני צינורות מתוך חמישה; בקבוצה השלישית של הצינורות, לא נצפתה צמיחה באף אחד מהצינורות (דילול 0.1 מ"ל). המספרים 5, 2 ו- 0 מושווים לאיור B1 בנספח ב ', שנבנה באמצעות מודל הסתברות של הליך הדגימה. מקריאת הטבלה שלנו, אנו מסיקים כי 49 הוא המספר הסביר ביותר של חיידקים לכל 100 מ"ל מי בריכה.
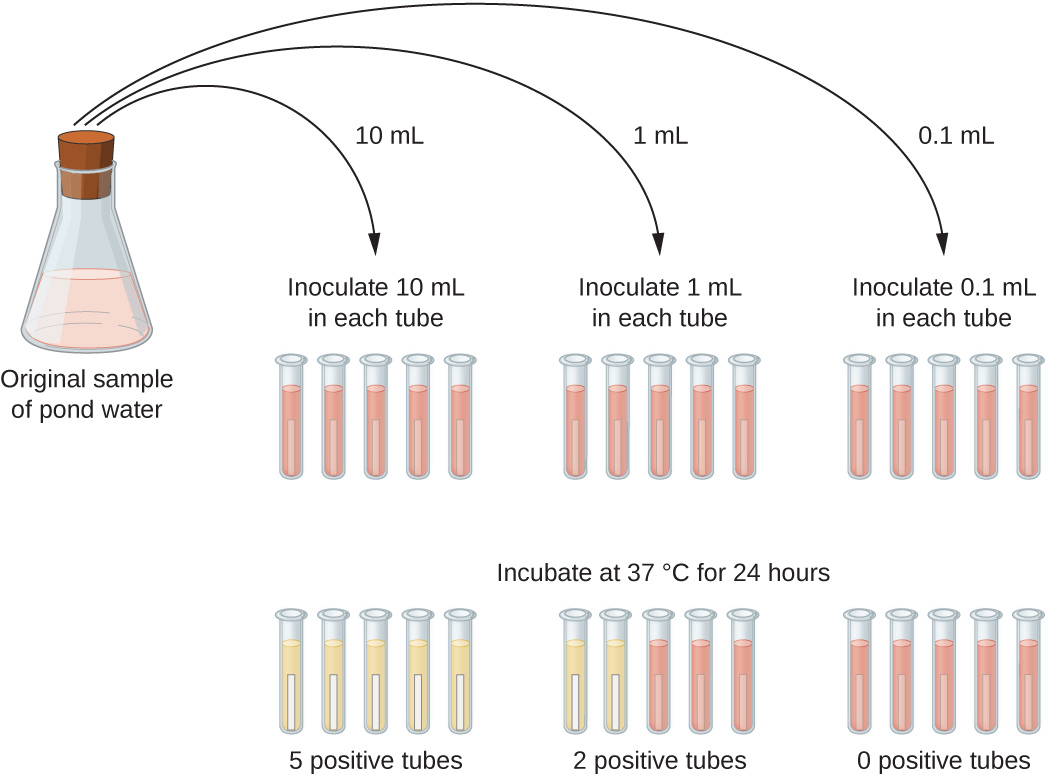
תרגיל \(\PageIndex{6}\)
- מהי יחידה יוצרת מושבה?
- אילו שתי שיטות משמשות לעתים קרובות להערכת מספר חיידקים בדגימות מים?
ספירת תאים עקיפה
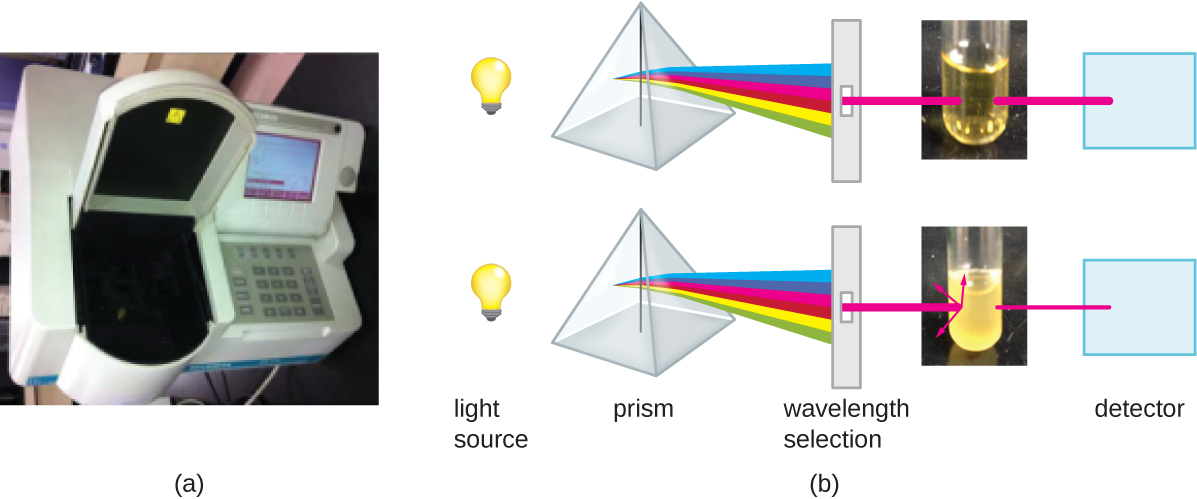
מלבד שיטות ישירות לספירת תאים, שיטות אחרות, המבוססות על זיהוי עקיף של צפיפות התאים, משמשות בדרך כלל להערכת והשוואת צפיפות התאים בתרבית. הגישה העיקרית היא למדוד את העכירות (עננות) של דגימת חיידקים בתרחיף נוזלי. מכשיר המעבדה המשמש למדידת עכירות נקרא ספקטרופוטומטר (איור). \(\PageIndex{14}\) בספקטרופוטומטר מועברת קרן אור דרך תרחיף חיידקי, האור העובר דרך המתלה נמדד על ידי גלאי, וכמות האור העוברת דרך הדגימה ומגיעה לגלאי מומרת לאחוז שידור או לערך לוגריתמי הנקרא ספיגה (צפיפות אופטית). ככל שמספר החיידקים בתרחיף עולה, גם העכירות עולה וגורמת פחות אור להגיע לגלאי. הירידה באור העובר דרך הדגימה ומגיעה לגלאי קשורה לירידה באחוזים שידור ועלייה בספיגה הנמדדת על ידי הספקטרופוטומטר.
מדידת עכירות היא שיטה מהירה להערכת צפיפות התאים כל עוד יש מספיק תאים במדגם כדי לייצר עכירות. ניתן לתאם קריאות עכירות למספר התאים בפועל על ידי ביצוע ספירת צלחות בת קיימא של דגימות שנלקחו מתרבויות בעלות מגוון ערכי ספיגה. באמצעות ערכים אלה, עקומת כיול נוצרת על ידי התוויית עכירות כפונקציה של צפיפות התא. לאחר יצירת עקומת הכיול, ניתן להשתמש בה להערכת ספירת תאים עבור כל הדגימות שהושגו או מתורבתות בתנאים דומים ועם צפיפות בטווח הערכים המשמשים לבניית העקומה.
מדידת משקל יבש של דגימת תרבית היא שיטה עקיפה נוספת להערכת צפיפות התרבות מבלי למדוד ישירות את ספירת התאים. יש לרכז את תרחיף התא המשמש לשקילה על ידי סינון או צנטריפוגה, לשטוף ולאחר מכן לייבש לפני ביצוע המדידות. יש לתקנן את מידת הייבוש כדי להסביר את תכולת המים השיורית. שיטה זו שימושית במיוחד עבור מיקרואורגניזמים חוטים, שקשה למנות אותם על ידי ספירת צלחות ישירה או בת קיימא.
כפי שראינו, שיטות להערכת מספרי תאים ברי קיימא יכולות להיות עתירות עבודה ולקחת זמן מכיוון שיש לגדל תאים. לאחרונה פותחו דרכים עקיפות למדידת תאים חיים שהן מהירות וקלות ליישום. שיטות אלה מודדות את פעילות התא על ידי ביצוע ייצור של מוצרים מטבוליים או היעלמות של מגיבים. ניתן לעקוב אחר היווצרות אדנוסין טריפוספט (ATP), ביוסינתזה של חלבונים וחומצות גרעין וצריכת חמצן כדי להעריך את מספר התאים.
תרגיל \(\PageIndex{7}\)
- מהי המטרה של עקומת כיול בעת הערכת ספירת תאים ממדידות עכירות?
- מהן השיטות העקיפות החדשות יותר לספירת תאים חיים?
דפוסים חלופיים של חלוקת תאים
ביקוע בינארי הוא הדפוס הנפוץ ביותר של חלוקת תאים בפרוקריוטים, אך הוא אינו היחיד. מנגנונים אחרים כוללים בדרך כלל חלוקה א-סימטרית (כמו בניצנים) או ייצור נבגים בחוטים אוויריים.
בחלק מהציאנובקטריה, נוקלאואידים רבים עשויים להצטבר בתא עגול מוגדל או לאורך נימה, מה שמוביל ליצירת תאים חדשים רבים בבת אחת. התאים החדשים מתפצלים לעתים קרובות מחוט האב וצפים משם בתהליך שנקרא פיצול (איור\(\PageIndex{15}\)). פיצול נצפה בדרך כלל ב-Actinomycetes, קבוצה של חיידקים אנאירוביים חיוביים לגרם הנמצאים בדרך כלל באדמה. דוגמה מוזרה נוספת לחלוקת תאים בפרוקריוטים, המזכירה לידה חיה בבעלי חיים, מוצגת על ידי החיידק הענק Epulopiscium. מספר תאי בת גדלים במלואם בתא האב, שבסופו של דבר מתפרק ומשחרר את התאים החדשים לסביבה. מינים אחרים עשויים ליצור שלוחה צרה וארוכה בקוטב אחד בתהליך הנקרא ניצנים. קצה הרחבה מתנפח ויוצר תא קטן יותר, הניצן שמתנתק בסופו של דבר מתא האב. ניצנים נפוצים בעיקר בשמרים (איור\(\PageIndex{15}\)), אך הם נצפים גם בחיידקי תותבות ובכמה ציאנובקטריה.

חיידקי הקרקע Actinomyces גדלים בחוטים ארוכים המחולקים על ידי מחיצות, בדומה לתפטיר הנראה בפטריות, וכתוצאה מכך תאים ארוכים עם נוקלאואידים מרובים. אותות סביבתיים, הקשורים כנראה לזמינות נמוכה של חומרים מזינים, מובילים להיווצרות חוטים אוויריים. בתוך חוטים אוויריים אלה, תאים מוארכים מתחלקים בו זמנית. התאים החדשים, המכילים נוקלאואיד יחיד, מתפתחים לנבגים המולידים מושבות חדשות.
תרגיל \(\PageIndex{8}\)
זהה לפחות הבדל אחד בין פיצול לנביטה.
ביופילמים
בטבע, מיקרואורגניזמים גדלים בעיקר בביופילמים, במערכות אקולוגיות מורכבות ודינאמיות הנוצרות על מגוון משטחים סביבתיים, החל מצינורות תעשייתיים וצינורות טיפול במים ועד סלעים בערוצי נהר. עם זאת, ביופילמים אינם מוגבלים למצעי משטח מוצקים. כמעט כל משטח בסביבה נוזלית המכיל כמה חומרים מזינים מינימליים יפתח בסופו של דבר ביופילם. מחצלות מיקרוביאליות שצפות על מים, למשל, הן ביופילמים המכילים אוכלוסיות גדולות של מיקרואורגניזמים פוטוסינתטיים. ביופילמים המצויים בפה האנושי עשויים להכיל מאות מיני חיידקים. ללא קשר לסביבה בה הם מתרחשים, ביופילמים אינם אוספים אקראיים של מיקרואורגניזמים; במקום זאת, מדובר בקהילות מובנות מאוד המספקות יתרון סלקטיבי למיקרואורגניזמים המרכיבים אותן.
מבנה ביופילם
תצפיות באמצעות מיקרוסקופיה קונפוקלית הראו שתנאי הסביבה משפיעים על המבנה הכללי של ביופילמים. ביופילמים חוטיים הנקראים סטרימרים נוצרים במים זורמים במהירות, כגון זרמי מים מתוקים, מערבולות ותאי זרימת מעבדה שתוכננו במיוחד המשכפלים תנאי גדילה בנוזלים הנעים במהירות. הסטרימרים מעוגנים למצע על ידי "ראש" וה"זנב "צף במורד הזרם בזרם. במים דוממים או איטיים, ביופילמים מקבלים בעיקר צורה דמוית פטריות. מבנה הביופילמים עשוי להשתנות גם עם תנאים סביבתיים אחרים כגון זמינות חומרים מזינים.
תצפיות מפורטות על ביופילמים תחת לייזר קונפוקלי ומיקרוסקופים אלקטרונים סורקים חושפות אשכולות של מיקרואורגניזמים המוטבעים במטריצה שזורה בתעלות מים פתוחות. המטריצה החוץ -תאית מורכבת מחומרים פולימריים חוץ-תאיים (EPS) המופרשים על ידי האורגניזמים בביופילם. המטריצה החוץ -תאית מייצגת חלק גדול מהביופילם, המהווה 50% — 90% מכלל המסה היבשה. תכונות ה- EPS משתנות בהתאם לאורגניזמים התושבים ולתנאי הסביבה.
EPS הוא ג'ל מיובש המורכב בעיקר מפוליסכרידים ומכיל מקרומולקולות אחרות כגון חלבונים, חומצות גרעין ושומנים. זה ממלא תפקיד מפתח בשמירה על שלמותו ותפקודו של הביופילם. תעלות ב-EPS מאפשרות תנועה של חומרים מזינים, פסולת וגזים ברחבי הביופילם. זה שומר על התאים לחות, ומונע התייבשות. EPS גם מחסה אורגניזמים בביופילם מפני טריפה על ידי חיידקים או תאים אחרים (למשל, פרוטוזואנים, תאי דם לבנים בגוף האדם).
היווצרות ביופילם
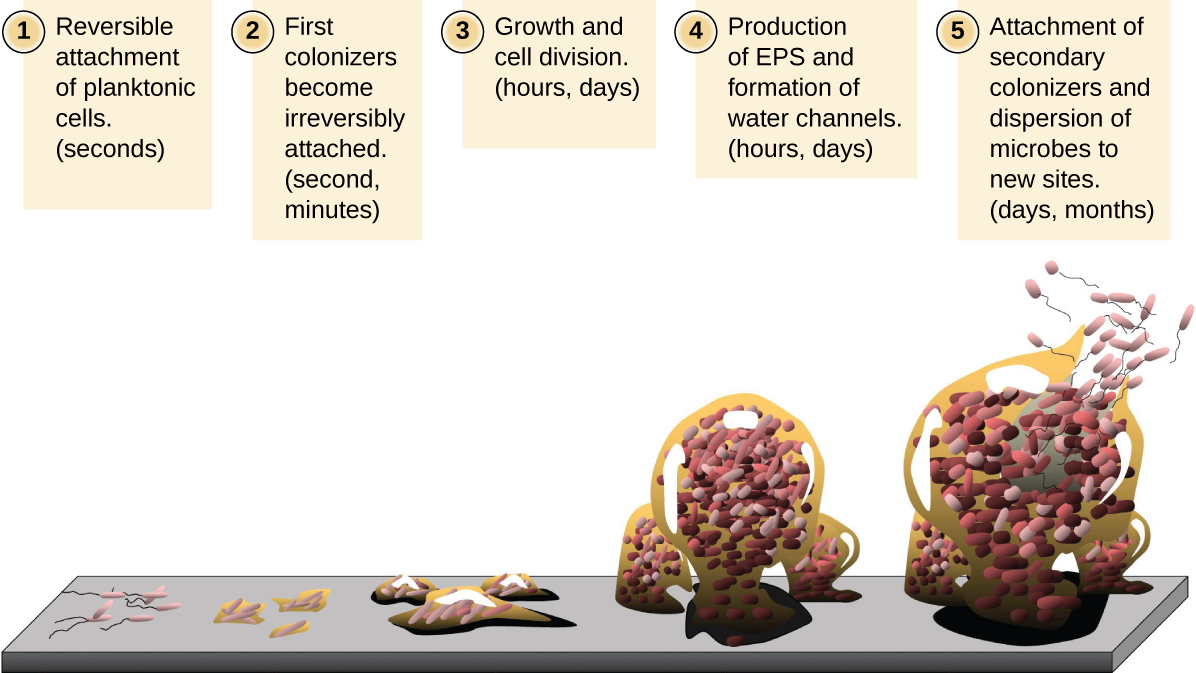
תאים מיקרוביאליים צפים חופשיים החיים בסביבה מימית נקראים תאים פלנקטוניים. היווצרות ביופילם כרוכה בעצם בהתקשרות של תאים פלנקטוניים למצע, שם הם הופכים לנייחים (מחוברים למשטח). זה קורה בשלבים, כפי שמתואר באיור\(\PageIndex{16}\). השלב הראשון כולל חיבור של תאים פלנקטוניים למשטח המצופה בסרט מיזוג של חומר אורגני. בשלב זה, ההתקשרות למצע היא הפיכה, אך כאשר התאים מבטאים פנוטיפים חדשים המקלים על היווצרות EPS, הם עוברים מאורח חיים פלנקטוני לאורח חיים שובל. הביופילם מפתח מבנים אופייניים, כולל מטריצה נרחבת ותעלות מים. נספחים כגון fimbriae, pili ו-flagella מקיימים אינטראקציה עם ה-EPS, ומיקרוסקופיה וניתוח גנטי מצביעים על כך שמבנים כאלה נדרשים להקמת ביופילם בוגר. בשלב האחרון של מחזור החיים הביופילם, תאים בפריפריה של הביופילם חוזרים לאורח חיים פלנקטוני, ומחליקים את הביופילם הבוגר כדי ליישב אתרים חדשים. שלב זה מכונה פיזור.
בתוך ביופילם, מינים שונים של מיקרואורגניזמים יוצרים שיתופי פעולה מטבוליים שבהם תוצר הפסולת של אורגניזם אחד הופך לחומר התזונתי של אחר. לדוגמה, מיקרואורגניזמים אירוביים צורכים חמצן ויוצרים אזורים אנאירוביים המקדמים את צמיחתם של אנאירובים. זה מתרחש בזיהומים פולימיקרוביאליים רבים הכוללים פתוגנים אירוביים ואנאירוביים כאחד.
המנגנון שבאמצעותו תאים בביופילם מתאמים את פעילותם בתגובה לגירויים סביבתיים נקרא חישת מניין. חישה מניין - שיכולה להתרחש בין תאים ממינים שונים בתוך ביופילם - מאפשרת למיקרואורגניזמים לזהות את צפיפות התאים שלהם באמצעות שחרור וקשירה של מולקולות קטנות וניתנות לפיזור הנקראות אוטואינדוקרים. כאשר אוכלוסיית התאים מגיעה לסף קריטי (מניין), גורמים אוטומטיים אלה יוזמים מפל של תגובות המפעילות גנים הקשורים לתפקודים תאיים המועילים רק כאשר האוכלוסייה מגיעה לצפיפות קריטית. לדוגמה, בחלק מהפתוגנים, סינתזה של גורמי ארסיות מתחילה רק כאשר קיימים מספיק תאים כדי להציף את ההגנה החיסונית של המארח. למרות שנחקרה בעיקר באוכלוסיות חיידקים, חישת מניין מתרחשת בין חיידקים לאאוקריוטים ובין תאים אוקריוטיים כגון הפטרייה קנדידה אלביקנס, חבר נפוץ במיקרוביוטה האנושית שעלולה לגרום לזיהומים אצל אנשים מדוכאי חיסון.
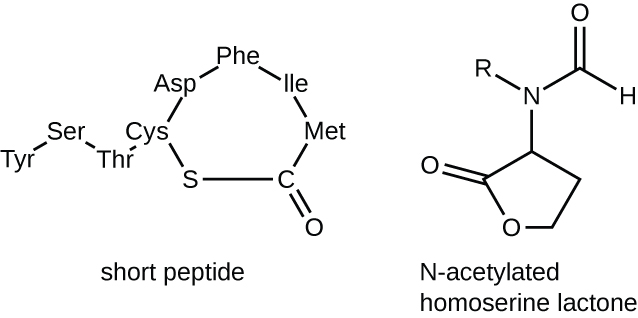
מולקולות האיתות בחישת המניין שייכות לשתי מחלקות עיקריות. חיידקים גראם שליליים מתקשרים בעיקר באמצעות לקטונים הומוסרין N-acylated, בעוד שחיידקים גרם חיוביים משתמשים בעיקר בפפטידים קטנים (איור). \(\PageIndex{17}\) בכל המקרים, השלב הראשון בחישת הקוורום מורכב מקשירה של ה-autoinducer לקולטן הספציפי שלו רק כאשר מגיעים לריכוז סף של מולקולות איתות. ברגע שהקישור לקולטן מתרחש, מפל של אירועי איתות מוביל לשינויים בביטוי הגנים. התוצאה היא הפעלת תגובות ביולוגיות הקשורות לחישת מניין, בעיקר עלייה בייצור מולקולות האיתות עצמן, ומכאן המונח autoinducer.
ביופילמים ובריאות האדם
גוף האדם מכיל סוגים רבים של ביופילמים, חלקם מועילים וחלקם מזיקים. לדוגמה, שכבות המיקרוביוטה התקינה המצפות את רירית המעי והנשימה ממלאות תפקיד בהדחת זיהומים על ידי פתוגנים. עם זאת, ביופילמים אחרים בגוף יכולים להשפיע לרעה על הבריאות. לדוגמא, הרובד שנוצר על השיניים הוא ביופילם שיכול לתרום למחלות שיניים וחניכיים. ביופילמים יכולים להיווצר גם בפצעים, ולעיתים לגרום לזיהומים חמורים העלולים להתפשט. החיידק Pseudomonas aeruginosa מיישב לעיתים קרובות ביופילמים בדרכי הנשימה של חולים עם סיסטיק פיברוזיס, וגורם לזיהומים כרוניים ולעיתים קטלניים של הריאות. ביופילמים יכולים להיווצר גם במכשירים רפואיים המשמשים בגוף או בגוף, ולגרום לזיהומים בחולים עם צנתרים, מפרקים מלאכותיים או עדשות מגע.
פתוגנים המוטבעים בתוך ביופילמים מפגינים עמידות גבוהה יותר לאנטיביוטיקה מאשר עמיתיהם הצפים חופשיים. הוצעו מספר השערות כדי להסביר מדוע. תאים בשכבות העמוקות של ביופילם אינם פעילים מטבולית ועשויים להיות פחות רגישים לפעולה של אנטיביוטיקה המשבשת את הפעילות המטבולית. ה-EPS עשוי גם להאט את התפשטות האנטיביוטיקה והחיטוי, ולמנוע מהם להגיע לתאים בשכבות העמוקות יותר של הביופילם. שינויים פנוטיפיים עשויים לתרום גם לעמידות המוגברת שמציגים תאים חיידקיים בביופילמים. לדוגמה, הייצור המוגבר של משאבות זרימה, חלבונים משובצים בממברנה המוציאים אנטיביוטיקה באופן פעיל מתאי חיידקים, הוכח כמנגנון חשוב של עמידות לאנטיביוטיקה בקרב חיידקים הקשורים לביופילם. לבסוף, ביופילמים מספקים סביבה אידיאלית לחילופי DNA חוץ-כרומוזומלי, הכולל לעתים קרובות גנים המעניקים עמידות לאנטיביוטיקה.
תרגיל \(\PageIndex{9}\)
- ממה מורכבת המטריצה של ביופילם?
- מהו תפקידה של חישת מניין בביופילם?
מושגי מפתח וסיכום
- רוב תאי החיידקים מתחלקים בביקוע בינארי. זמן הדור בגידול חיידקים מוגדר כזמן ההכפלה של האוכלוסייה.
- תאים במערכת סגורה עוקבים אחר דפוס צמיחה עם ארבעה שלבים: פיגור, לוגריתמי (אקספוננציאלי), נייח ומוות.
- תאים ניתן לספור על ידי ספירת תאים קיימא ישירה. צלחת המזיגה ושיטות צלחת הפיזור משמשות לצלחת דילולים סדרתיים לתוך או על אגר, בהתאמה, כדי לאפשר ספירה של תאים ברי קיימא המולידים יחידות יוצרות מושבה. סינון ממברנה משמש לספירת תאים חיים בתמיסות מדוללות. שיטת מספר התא הסביר ביותר (MPN) מאפשרת הערכה של מספרי תאים בתרבויות ללא שימוש במדיה מוצקה.
- ניתן להשתמש בשיטות עקיפות להערכת צפיפות התרבות על ידי מדידת עכירות של תרבית או צפיפות תאים חיים על ידי מדידת פעילות מטבולית.
- דפוסים אחרים של חלוקת תאים כוללים היווצרות נוקלאואידית מרובה בתאים; חלוקה א-סימטרית, כמו בניצנים; ויצירת היפות ונבגים סופניים.
- ביופילמים הם קהילות של מיקרואורגניזמים הכרוכים במטריצה של חומר פולימרי חוץ תאי. היווצרות ביופילם מתרחשת כאשר תאים פלנקטוניים מתחברים למצע והופכים לנייחים. תאים בביופילמים מתאמים את פעילותם על ידי תקשורת באמצעות חישת מניין.
- ביופילמים נמצאים בדרך כלל על משטחים בטבע ובגוף האדם, שם הם עשויים להועיל או לגרום לזיהומים חמורים. פתוגנים הקשורים לביופילמים הם לרוב עמידים יותר לאנטיביוטיקה וחומרי חיטוי.