
13.3: פרוטיסטים
- Page ID
- 208923

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)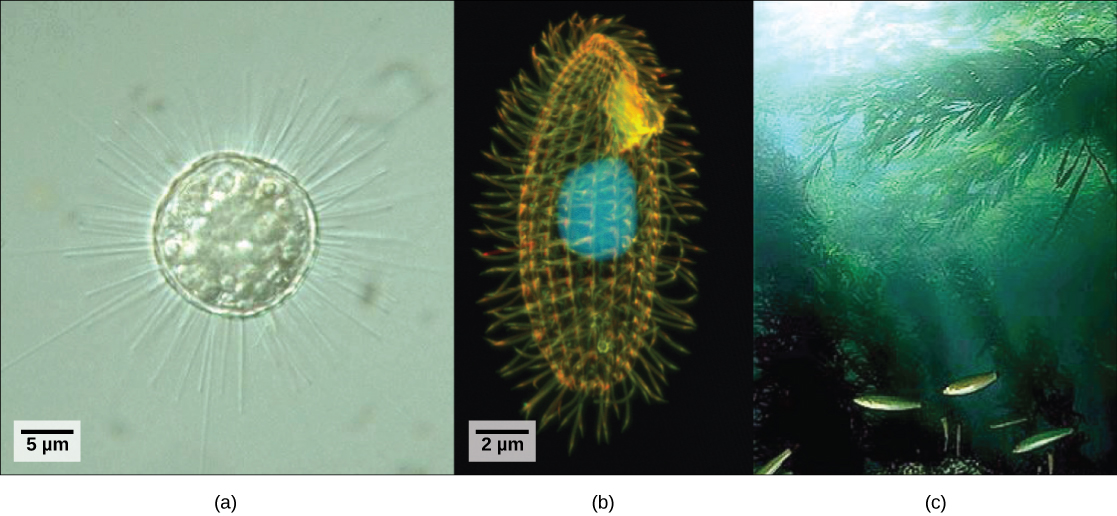
אורגניזמים אוקריוטיים שלא התאימו לקריטריונים של הממלכות Animalia, Fungi או Plantae מבחינה היסטורית נקראו פרוטיסטים וסווגו לממלכה פרוטיסטה. הפרוטיסטים כוללים את האאוקריוטים החד-תאיים החיים במי בריכה (איור\(\PageIndex{1}\)), אם כי מינים פרוטיסטים חיים במגוון סביבות מימיות ויבשתיות אחרות, ותופסים נישות רבות ושונות. לא כל הפרוטיסטים הם מיקרוסקופיים וחד-תאיים; קיימים כמה מינים רב-תאיים גדולים מאוד, כמו האצות. במהלך שני העשורים האחרונים, תחום הגנטיקה המולקולרית הוכיח שחלק מהפרוטיסטים קשורים יותר לבעלי חיים, צמחים או פטריות מאשר לפרוטיסטים אחרים. מסיבה זו, שושלות פרוטיסטיות שסווגו במקור לממלכה פרוטיסטה הוקצו מחדש לממלכות חדשות או לממלכות קיימות אחרות. השושלות האבולוציוניות של הפרוטיסטים ממשיכות להיבחן ולהתלבט. בינתיים, המונח "פרוטיסט" עדיין משמש באופן לא פורמלי לתיאור קבוצת האוקריוטים המגוונת להפליא הזו. כקבוצה קולקטיבית, פרוטיסטים מציגים מגוון מדהים של מורפולוגיות, פיזיולוגיות ואקולוגיות.
מאפייני הפרוטיסטים
ישנם למעלה מ-100,000 מינים חיים מתוארים של פרוטיסטים, ולא ברור כמה מינים לא מתוארים עשויים להתקיים. מכיוון שפרוטיסטים רבים חיים במערכות יחסים סימביוטיות עם אורגניזמים אחרים ויחסים אלה הם לרוב ספציפיים למינים, קיים פוטנציאל עצום למגוון פרוטיסטי לא מתואר התואם את מגוון המארחים. כמונח המפתח לאורגניזמים אוקריוטיים שאינם בעלי חיים, צמחים, פטריות או כל קבוצה אחת הקשורה לפילוגנטית, אין זה מפתיע שמעט מאפיינים משותפים לכל הפרוטיסטים.
כמעט כל הפרוטיסטים קיימים בסביבה מימית כלשהי, כולל סביבות מים מתוקים וימיים, אדמה לחה ואפילו שלג. כמה מינים פרוטיסטיים הם טפילים המדביקים בעלי חיים או צמחים. טפיל הוא אורגניזם שחי על אורגניזם אחר או בתוכו וניזון ממנו, לעתים קרובות מבלי להרוג אותו. כמה מינים פרוטיסטים חיים על אורגניזמים מתים או על הפסולת שלהם, ותורמים לריקבון שלהם.
מבנה פרוטיסטי
התאים של הפרוטיסטים הם בין התאים המשוכללים ביותר מכל התאים. רוב הפרוטיסטים הם מיקרוסקופיים וחד-תאיים, אך קיימות כמה צורות רב-תאיות אמיתיות. כמה פרוטיסטים חיים כמושבות שמתנהגות במובנים מסוימים כקבוצה של תאים חיים חופשיים ובדרכים אחרות כאורגניזם רב תאי. פרוטיסטים אחרים מורכבים מתאי יחיד עצומים, רב-גרעיניים, שנראים כמו כתמי רפש אמורפיים או, במקרים אחרים, כמו שרכים. למעשה, תאים פרוטיסטיים רבים הם רב-גרעיניים; במינים מסוימים הגרעינים בגדלים שונים ויש להם תפקידים מובחנים בתפקוד התא הפרוטיסטי.
תאים פרוטיסטים בודדים נעים בגודלם בין פחות ממיקרומטר לאורכים של 3 מטרים של התאים הרב-גרעיניים של האצה Caulerpa. תאים פרוטיסטים עשויים להיות עטופים בממברנות תאים דמויי בעלי חיים או בדפנות תאים דמויי צמחים. אחרים עטופים בקליפות מבוססות סיליקה מזוגגות או פצועות בכדורים של רצועות חלבון משתלבות. הכדור מתפקד כמו מעיל שריון גמיש, ומונע מהפרוטיסט להיקרע או לנקב מבלי לפגוע בטווח התנועה שלו.
רוב הפרוטיסטים הם תנועתיים, אך סוגים שונים של פרוטיסטים פיתחו אופני תנועה מגוונים. לחלק מהפרוטיסטים יש דגל אחד או יותר, שהם מסובבים או מקציפים. אחרים מכוסים בשורות או בציציות של ציציות זעירות שהם מכים בתיאום לשחייה. אחרים שולחים פסאודופודיה דמוית אונה מכל מקום בתא, מעגנים את הפסאודופודיום למצע ומושכים את שאר התא לכיוון נקודת העיגון. חלק מהפרוטיסטים יכולים לנוע לעבר האור על ידי צימוד אסטרטגיית התנועה שלהם לאיבר חישה אור.
כיצד פרוטיסטים משיגים אנרגיה
פרוטיסטים מפגינים צורות תזונה רבות ועשויים להיות אירוביים או אנאירוביים. פרוטיסטים פוטוסינתטיים (פוטואוטוטרופים) מאופיינים בנוכחות כלורופלסטים. פרוטיסטים אחרים הם הטרוטרופים וצורכים חומרים אורגניים (כמו אורגניזמים אחרים) כדי להשיג תזונה. אמבות וכמה מינים פרוטיסטיים הטרוטרופיים אחרים בולעים חלקיקים בתהליך הנקרא פגוציטוזיס, שבו קרום התא בולע חלקיק מזון ומכניס אותו פנימה, צובט שק קרומי תוך תאי, או שלפוחית, הנקראת ואקום מזון (איור). \(\PageIndex{2}\) שלפוחית זו מתמזגת לאחר מכן עם ליזוזום, וחלקיק המזון מתפרק למולקולות קטנות שיכולות להתפזר לתוך הציטופלזמה ולהשתמש בהן בחילוף החומרים התאי. שרידים לא מעוכלים גורשים בסופו של דבר מהתא באמצעות אקסוציטוזיס.
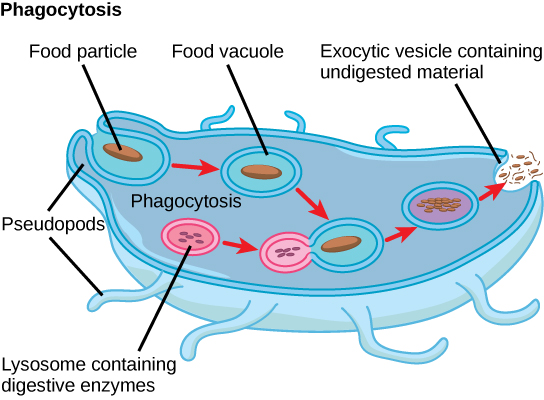
חלק מההטרוטרופים סופגים חומרים מזינים מאורגניזמים מתים או מהפסולת האורגנית שלהם, ואחרים מסוגלים להשתמש בפוטוסינתזה או להאכיל מחומרים אורגניים, בהתאם לתנאים.
רפרודוקציה
פרוטיסטים מתרבים על ידי מגוון מנגנונים. רובם מסוגלים צורה כלשהי של רבייה א -מינית, כגון ביקוע בינארי לייצר שני תאי בת, או ביקוע מרובה להתחלק בו זמנית לתאי בת רבים. אחרים מייצרים ניצנים זעירים שממשיכים להתחלק וגדלים לגודל הפרוטיסט ההורי. רבייה מינית, הכוללת מיוזה והפריה, נפוצה בקרב פרוטיסטים, ומינים פרוטיסטים רבים יכולים לעבור מרבייה א-מינית למינית בעת הצורך. רבייה מינית קשורה לעיתים קרובות לתקופות בהן חומרים מזינים מתרוקנים או מתרחשים שינויים סביבתיים. רבייה מינית עשויה לאפשר לפרוטיסט לשלב מחדש גנים ולייצר וריאציות חדשות של צאצאים שעשויות להתאים יותר לשרוד בסביבה החדשה. עם זאת, רבייה מינית קשורה לעיתים קרובות גם לציסטות המהוות שלב מגן ומנוחה. בהתאם לבית הגידול שלהם, הציסטות עשויות להיות עמידות במיוחד בפני טמפרטורות קיצוניות, התייבשות או pH נמוך. אסטרטגיה זו מאפשרת גם לפרוטיסטים מסוימים "להמתין" לגורמי לחץ עד שסביבתם תהיה נוחה יותר להישרדות או עד שהם נישאים (כגון על ידי רוח, מים או הובלה על אורגניזם גדול יותר) לסביבה אחרת מכיוון שציסטות אינן מציגות כמעט תא חילוף חומרים.
גיוון פרוטיסטי
עם הופעת רצף ה- DNA, היחסים בין קבוצות פרוטיסטים ובין קבוצות פרוטיסטיות לאיקריוטים אחרים מתחילים להתבהר. מערכות יחסים רבות שהתבססו על קווי דמיון מורפולוגיים מוחלפות במערכות יחסים חדשות המבוססות על קווי דמיון גנטיים. פרוטיסטים המציגים מאפיינים מורפולוגיים דומים עשויים להתפתח מבנים אנלוגיים בגלל לחצים סלקטיביים דומים - ולא בגלל מוצא משותף לאחרונה. תופעה זו נקראת אבולוציה מתכנסת. זו אחת הסיבות לכך שהסיווג הפרוטיסטי כל כך מאתגר. תכנית הסיווג המתהווה מקבצת את כל התחום אוקריוטה לשש "קבוצות-על" המכילות את כל הפרוטיסטים וכן בעלי חיים, צמחים ופטריות (איור\(\PageIndex{3}\)); אלה כוללים את Excavata, Chromalveolata, Rhizaria, Archeaplastida, Amoebozoa ו-Opisthokonta. מאמינים כי קבוצות העל הן מונופילטיות; כל האורגניזמים בתוך כל קבוצת-על מאמינים שהתפתחו מאב קדמון משותף אחד, ולכן כל החברים קשורים זה לזה באופן הדוק ביותר מאשר לאורגניזמים מחוץ לקבוצה זו. עדיין חסרות עדויות למונופיליה של קבוצות מסוימות.
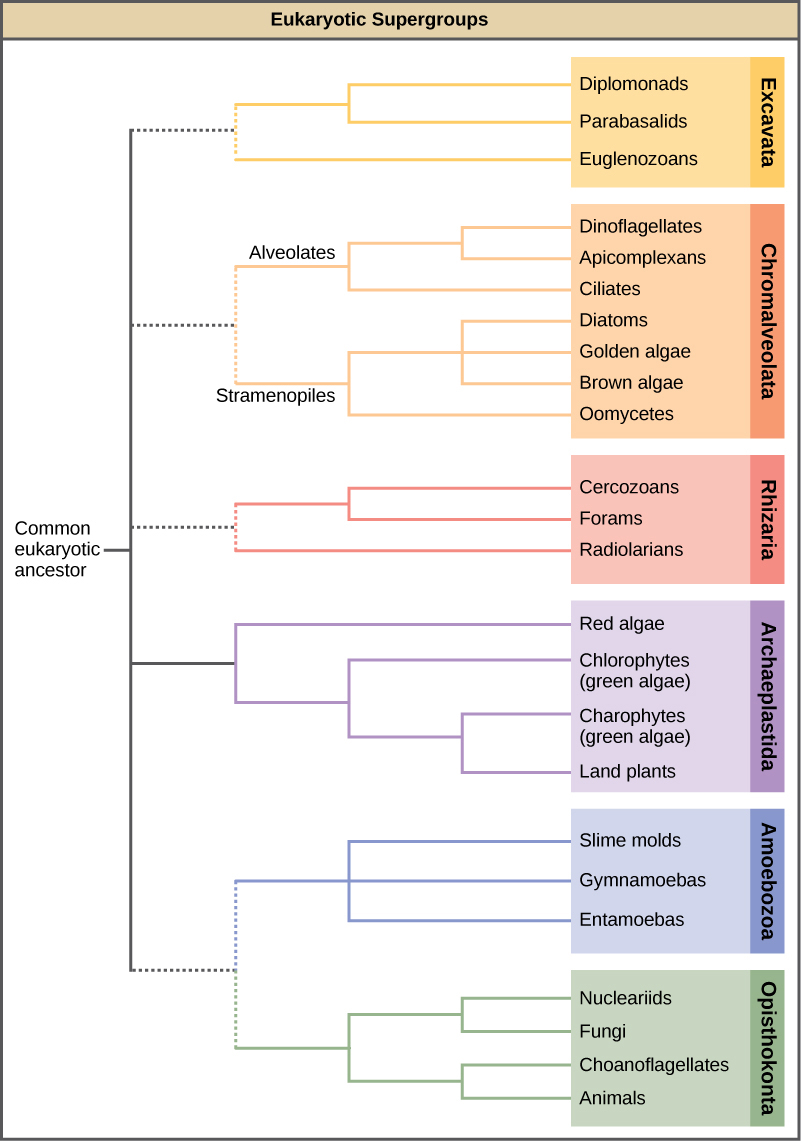
פתוגנים אנושיים
פרוטיסטים רבים הם טפילים פתוגניים שחייבים להדביק אורגניזמים אחרים כדי לשרוד ולהתפשט. טפילים פרוטיסטים כוללים את הגורמים הסיבתיים למלריה, מחלת שינה אפריקאית וגסטרואנטריטיס הנישאת במים בבני אדם. פתוגנים פרוטיסטים אחרים טורפים צמחים, וגורמים להרס מאסיבי של גידולי מזון.
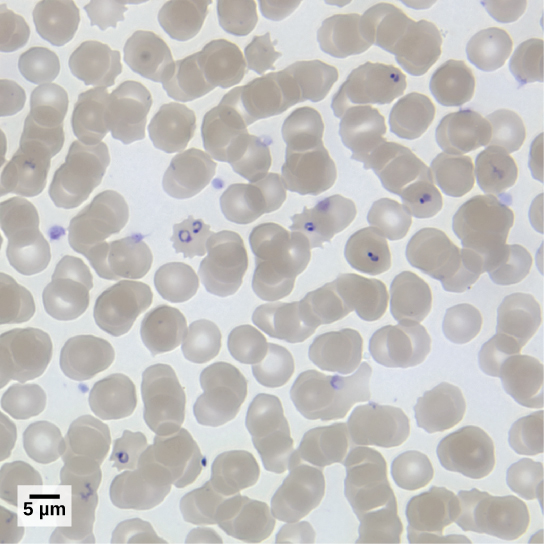
מינים פלסמודיום
חברי הסוג פלסמודיום חייבים להדביק יתוש וחוליות כדי להשלים את מחזור חייהם. אצל בעלי חוליות, הטפיל מתפתח בתאי כבד וממשיך להדביק תאי דם אדומים, פורצים מתאי הדם ומשמידים אותם בכל מחזור שכפול א-מיני (איור\(\PageIndex{4}\)). מבין ארבעת מיני הפלסמודיום הידועים כמדביקים בני אדם, P. falciparum מהווה 50 אחוז מכלל מקרי המלריה והוא הגורם העיקרי לתמותה הקשורה למחלות באזורים טרופיים בעולם. בשנת 2010 הוערך כי מלריה גרמה בין 0.5 למיליון מקרי מוות, בעיקר בילדים אפריקאים. במהלך המלריה, P. falciparum יכול להדביק ולהשמיד יותר ממחצית מתאי הדם המסתובבים של האדם, מה שמוביל לאנמיה קשה. בתגובה למוצרי פסולת המשתחררים כשהטפילים מתפרצים מתאי דם נגועים, מערכת החיסון המארחת מגבירה תגובה דלקתית מסיבית עם פרקי חום מעוררי הזיות, כאשר טפילים הורסים תאי דם אדומים, ושופכים פסולת טפילים לזרם הדם. P. falciparum מועבר לבני אדם על ידי יתוש המלריה האפריקאי, אנופלס גמביה. טכניקות להרוג, לעקר או להימנע מחשיפה למין יתושים אגרסיבי מאוד זה חיוניות לשליטה במלריה.
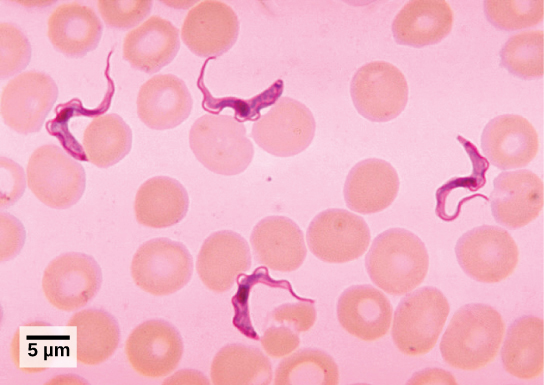
טריפנוזומים
T. brucei, הטפיל שאחראי על מחלת השינה האפריקאית, מבלבל את מערכת החיסון האנושית על ידי שינוי השכבה העבה של גליקופרוטאינים על פני השטח עם כל מחזור זיהומי (איור). \(\PageIndex{5}\) הגליקופרוטאינים מזוהים על ידי מערכת החיסון כחומר זר, והגנה ספציפית של נוגדנים מותקנת כנגד הטפיל. עם זאת, T. לברוסיי יש אלפי אנטיגנים אפשריים, ועם כל דור שלאחר מכן, הפרוטיסט עובר לציפוי גליקופרוטאין בעל מבנה מולקולרי שונה. בדרך זו, T. brucei מסוגל לשכפל ברציפות מבלי שמערכת החיסון תצליח אי פעם לנקות את הטפיל. ללא טיפול, מחלת שינה אפריקאית מובילה תמיד למוות בגלל נזק שהיא גורמת למערכת העצבים. בתקופות מגיפה, התמותה מהמחלה יכולה להיות גבוהה. אמצעי מעקב ובקרה גדולים יותר הובילו לצמצום המקרים המדווחים; חלק מהמספרים הנמוכים ביותר שדווחו מזה 50 שנה (פחות מ -10,000 מקרים בכל אפריקה שמדרום לסהרה) התרחשו מאז 2009.
באמריקה הלטינית, מין אחר בסוג, T. קרוזי, אחראי למחלת צ'אגאס. T. דלקות קרוזי נגרמות בעיקר על ידי באג מוצץ דם. הטפיל מאכלס רקמות לב ומערכת העיכול בשלב הכרוני של ההדבקה, מה שמוביל לתת תזונה ואי ספיקת לב הנגרמת על ידי קצב לב לא תקין. על פי ההערכות, 10 מיליון בני אדם נדבקים במחלת צ'אגאס, שגרמה ל -10,000 מקרי מוות בשנת 2008.
טפילי צמחים
טפילים פרוטסטיים של צמחים יבשתיים כוללים סוכנים המשמידים גידולי מזון. ה-oomycete Plasmopara viticola מטפיל צמחי ענבים וגורם למחלה הנקראת טחב פלומתי (איור א). \(\PageIndex{6}\) צמחי ענבים הנגועים ב- P. ויטיקולה נראית מעוכבת ויש לה עלים קמלים דהויים. התפשטות הטחב הפלומתי גרמה לקריסה הקרובה של תעשיית היין הצרפתית במאה התשע עשרה.

Phytophthora infestans הוא oomycete האחראי לדלקת מאוחרת של תפוחי אדמה, הגורמת לגבעולי תפוחי אדמה וגבעולים להתפורר לרפש שחור (איור ב). \(\PageIndex{6}\) דלקת תפוחי אדמה נרחבת הנגרמת על ידי P. אינפסטנים גרמו לרעב תפוחי האדמה האירי הידוע במאה התשע עשרה שגבה את חייהם של כמיליון בני אדם והוביל להגירה מאירלנד של לפחות מיליון נוספים. הדלקת המאוחרת ממשיכה לפגוע בגידולי תפוחי אדמה בחלקים מסוימים של ארצות הברית ורוסיה, ומחסלת עד 70 אחוז מהיבולים כאשר לא מורחים חומרי הדברה.
פרוטיסטים מועילים
פרוטיסטים ממלאים תפקידים אקולוגיים חשובים ביותר כיצרנים במיוחד באוקיינוסים בעולם. הם חשובים באותה מידה בקצה השני של קורי המזון כמו מפרקים.
פרוטיסטים כמקורות מזון
פרוטיסטים הם מקורות תזונה חיוניים לאורגניזמים רבים אחרים. במקרים מסוימים, כמו בפלנקטון, פרוטיסטים נצרכים ישירות. לחלופין, פרוטיסטים פוטוסינתטיים משמשים כמפיקים של תזונה לאורגניזמים אחרים על ידי קיבוע פחמן. לדוגמה, דינופלגלטים פוטוסינתטיים הנקראים zooxanthellae מעבירים את רוב האנרגיה שלהם לפוליפים האלמוגים המאכלסים אותם (איור). \(\PageIndex{7}\) במערכת יחסים מועילה הדדית זו, הפוליפים מספקים סביבה מגנה וחומרים מזינים לזואוקסנתלים. הפוליפים מפרישים את הסידן פחמתי הבונה שוניות אלמוגים. ללא סימביונים דינופלגלטים, האלמוגים מאבדים פיגמנטים של אצות בתהליך הנקרא הלבנת אלמוגים, ובסופו של דבר הם מתים. זה מסביר מדוע אלמוגים בוני שונית אינם שוכנים במים עמוקים מ -20 מטר: לא מספיק אור מגיע לעומקים אלה כדי שדינופלגלטים יוכלו לבצע פוטוסינתזה.

הפרוטיסטים עצמם ותוצרי הפוטוסינתזה שלהם חיוניים - במישרין או בעקיפין - להישרדותם של אורגניזמים החל מחיידקים ועד יונקים. כיצרנים ראשוניים, פרוטיסטים מאכילים חלק גדול מהמינים המימיים בעולם. (ביבשה, צמחים יבשתיים משמשים כיצרנים ראשוניים.) למעשה, כרבע מהפוטוסינתזה בעולם מתבצעת על ידי פרוטיסטים, במיוחד דינופלגלטים, דיאטומים ואצות רב תאיות.
פרוטיסטים אינם יוצרים מקורות מזון רק לאורגניזמים השוכנים בים. לדוגמה, מינים אנאירוביים מסוימים קיימים בדרכי העיכול של טרמיטים וג'וקים אוכלי עצים, שם הם תורמים לעיכול תאית הנבלעת על ידי חרקים אלה כשהם נודדים בעץ. האנזים בפועל המשמש לעיכול התאית מיוצר למעשה על ידי חיידקים החיים בתוך התאים הפרוטסטיים. הטרמיט מספק את מקור המזון לפרוטיסט ולחיידקים שלו, והפרוטיסט והחיידקים מספקים חומרים מזינים לטרמיט על ידי פירוק התאית.
סוכני פירוק
פרוטיסטים רבים דמויי פטריות הם ספרובים, אורגניזמים הניזונים מאורגניזמים מתים או מחומר הפסולת המיוצר על ידי אורגניזמים (ספרופיט הוא מונח שווה ערך), ומתמחים לספיגת חומרים מזינים מחומרים אורגניים שאינם חיים. לדוגמה, סוגים רבים של oomycetes גדלים על בעלי חיים מתים או אצות. לפרוטיסטים ספרוביים יש את התפקיד החיוני של החזרת חומרים מזינים אנאורגניים לאדמה ולמים. תהליך זה מאפשר גידול צמחים חדש, אשר בתורו מייצר מזון לאורגניזמים אחרים לאורך שרשרת המזון. ואכן, ללא מינים ספרוביים, כגון פרוטיסטים, פטריות וחיידקים, החיים יפסיקו להתקיים כאשר כל הפחמן האורגני "נקשר" באורגניזמים מתים.
סיכום מדור
הפרוטיסטים מגוונים ביותר מבחינת מאפיינים ביולוגיים ואקולוגיים, בעיקר בשל העובדה שהם מכלול מלאכותי של קבוצות שאינן קשורות פילוגנטית. פרוטיסטים מציגים מבני תאים מגוונים ביותר, מספר סוגים של אסטרטגיות רבייה, כמעט כל סוג תזונה אפשרי ובתי גידול מגוונים. רוב הפרוטיסטים החד-תאיים הם תנועתיים, אך אורגניזמים אלה משתמשים במבנים מגוונים לתחבורה.
תהליך סיווג הפרוטיסטים לקבוצות משמעותיות נמשך, אך נתונים גנטיים ב -20 השנים האחרונות הבהירו קשרים רבים שבעבר לא היו ברורים או שגויים. השקפת הרוב כיום היא להזמין את כל האיקריוטים לשש קבוצות-על. מטרת תכנית סיווג זו היא ליצור אשכולות של מינים שכולם נגזרים מאב קדמון משותף.
רשימת מילים
- אמבוזואה
- קבוצת העל האוקריוטית המכילה את האמבות ותבניות הרפש
- ארכפלסטידה
- קבוצת העל האוקריוטית המכילה צמחי יבשה, אצות ירוקות ואצות אדומות
- כרומלבולטה
- קבוצת העל האוקריוטית המכילה את הדינופלגלטים, הריסים, האצות החומות, הדיאטומים ותבניות המים
- מחפר
- קבוצת העל האוקריוטית המכילה אורגניזמים חד-תאיים מסומנים עם חריץ האכלה
- אופיסטוקונטה
- קבוצת העל האוקריוטית המכילה את הפטריות, בעלי החיים והצ'אנופלגלטים
- טפיל
- אורגניזם שחי על אורגניזם אחר או בתוכו וניזון ממנו, לעתים קרובות מבלי להרוג אותו
- כדורית
- כיסוי תא חיצוני המורכב מרצועות חלבון משתלבות המתפקדות כמו שכבת שריון גמישה, המונעת מהתאים להיקרע או לנקב מבלי לפגוע בטווח התנועה שלהם
- ריזריה
- קבוצת העל האוקריוטית המכילה אורגניזמים הנעים בתנועה אמבואידית
- ספרוב
- אורגניזם הניזון מחומר אורגני מת