
11.4: ייחוס
- Page ID
- 208735

ההגדרה הביולוגית של מינים, הפועלת עבור אורגניזמים המתרבים מינית, היא קבוצה של פרטים בפועל או פוטנציאליים להתרבות. על פי הגדרה זו, מין אחד נבדל ממין אחר על ידי האפשרות להזדווג בין פרטים מכל מין לייצר צאצאים פוריים. יש יוצאים מן הכלל לכלל זה. מינים רבים דומים מספיק כדי שצאצאים היברידיים אפשריים ועשויים להופיע לעתים קרובות בטבע, אך עבור רוב המינים כלל זה מתקיים בדרך כלל. למעשה, הימצאותם של כלאיים בין מינים דומים מעידה על כך שהם עשויים לנבוע ממין רבייה יחיד וכי ייתכן שתהליך הייחוד עדיין לא הושלם.
בהתחשב במגוון החיים יוצא הדופן על פני כדור הארץ חייבים להיות מנגנונים לייחוס: היווצרותם של שני מינים ממין מקורי אחד. דרווין ראה בעיני רוחו את התהליך הזה כאירוע מסועף ותרגם את התהליך באיור היחיד שנמצא ב על מוצא המינים (איור א). \(\PageIndex{1}\) כדי שיתרחש התגלמות, יש ליצור שתי אוכלוסיות חדשות מאוכלוסייה מקורית אחת, והן חייבות להתפתח בצורה כזו שלא יהיה אפשרי עבור פרטים משתי האוכלוסיות החדשות להתרבות. ביולוגים הציעו מנגנונים שבאמצעותם זה יכול להתרחש המתחלקים לשתי קטגוריות רחבות. ייחוס אלופטרי, כלומר ייחוס ב"ארצות מולדת אחרות ", כרוך בהפרדה גיאוגרפית של אוכלוסיות ממין הורה והתפתחות שלאחר מכן. ייחוס סימפטרי, כלומר ייחוס ב"אותה מולדת ", כרוך בהתייחסות המתרחשת בתוך מין הורה תוך הישארות במקום אחד.
ביולוגים חושבים על אירועי ייחוס כעל פיצול של מין אבות אחד לשני מינים צאצאים. אין שום סיבה שלא ייווצרו יותר משני מינים בו זמנית, אלא שזה פחות סביר וניתן להמשיג אירועים מרובים כאלה גם כפיצולים בודדים המתרחשים קרוב בזמן.

ייחוס באמצעות הפרדה גיאוגרפית
לאוכלוסייה רציפה גיאוגרפית יש מאגר גנים שהוא הומוגני יחסית. זרימת הגנים, תנועת האללים על פני טווח המינים, חופשית יחסית מכיוון שפרטים יכולים לנוע ואז להזדווג עם פרטים במיקומם החדש. לפיכך, תדירות האלל בקצה אחד של התפלגות תהיה דומה לתדירות האלל בקצה השני. כאשר אוכלוסיות הופכות לבלתי רציפות גיאוגרפית, נמנעת זרימה חופשית של אללים. כאשר הפרדה זו נמשכת פרק זמן, שתי האוכלוסיות מסוגלות להתפתח לאורך מסלולים שונים. לפיכך, תדרי האללים שלהם בלוקוסים גנטיים רבים הופכים בהדרגה שונים יותר ויותר ככל שאללים חדשים מתעוררים באופן עצמאי על ידי מוטציה בכל אוכלוסייה. בדרך כלל, תנאי הסביבה, כגון אקלים, משאבים, טורפים ומתחרים, עבור שתי האוכלוסיות יהיו שונים וגורמים לברירה הטבעית להעדיף התאמות שונות בכל קבוצה. היסטוריות שונות של סחף גנטי, המשופרות מכיוון שהאוכלוסיות קטנות יותר מאוכלוסיית ההורים, יובילו גם הן להתבדלות.
בהינתן מספיק זמן, ההתבדלות הגנטית והפנוטיפית בין אוכלוסיות תשפיע ככל הנראה על דמויות המשפיעות על הרבייה מספיק, כך שאילו יחידים משתי האוכלוסיות יובאו יחד, ההזדווגות תהיה פחות סבירה, או אם תתרחש הזדווגות, הצאצאים יהיו בלתי קיימא או עקרים. סוגים רבים של דמויות שונות עשויים להשפיע על בידוד הרבייה (חוסר יכולת להתרבות) של שתי האוכלוסיות. ניתן לחלק מנגנונים אלה של בידוד רבייה למנגנונים פרזיגוטיים (אלה הפועלים לפני ההפריה) ומנגנונים פוסט-זיגוטיים (אלה הפועלים לאחר ההפריה). מנגנונים פרה -זיגוטיים כוללים תכונות המאפשרות לאנשים למצוא זה את זה, כגון תזמון ההזדווגות, רגישות לפרומונים או בחירת אתרי הזדווגות. אם אנשים מסוגלים להיתקל זה בזה, סטיית אופי עשויה למנוע מטקסי חיזור להוביל להזדווגות בין אם בגלל שההעדפות הנשיות השתנו או שההתנהגויות הגבריות השתנו. שינויים פיזיולוגיים עלולים להפריע להפריה מוצלחת אם ההזדווגות מסוגלת להתרחש. מנגנונים פוסט-זיגוטיים כוללים חוסר תאימות גנטית המונעת התפתחות תקינה של הצאצאים, או אם הצאצאים חיים, ייתכן שהם אינם מסוגלים לייצר גמטות קיימא בעצמם כמו בדוגמה של הפרד, הצאצאים העקרים של סוס נקבה וחמור זכר.
אם שתי האוכלוסיות המבודדות יוחזרו יחד ולצאצאים ההיברידיים שנוצרו מהזדווגות בין פרטים משתי האוכלוסיות יש הישרדות נמוכה יותר או פוריות מופחתת, אז הברירה תעדיף פרטים המסוגלים להבחין בין בני זוג פוטנציאליים של האוכלוסייה שלהם לבין האוכלוסייה האחרת.. בחירה זו תשפר את בידוד הרבייה.
בידוד אוכלוסיות המובילות להתייחסות אלופטרית יכול להתרחש במגוון דרכים: מנהר היוצר ענף חדש, שחיקה היוצרת עמק חדש, או קבוצת אורגניזמים הנוסעים למקום חדש ללא יכולת לחזור, כגון זרעים שצפים מעל האוקיינוס לאי. אופי ההפרדה הגיאוגרפית הדרושה לבידוד אוכלוסיות תלוי לחלוטין בביולוגיה של האורגניזם ובפוטנציאל הפיזור שלו. אם שתי אוכלוסיות חרקים מעופפים יתגוררו בעמקים סמוכים נפרדים, רוב הסיכויים שאנשים מכל אוכלוסייה יעופו הלוך ושוב, וימשיכו בזרימת הגנים. עם זאת, אם שתי אוכלוסיות מכרסמים יחולקו על ידי היווצרות אגם חדש, המשך זרימת הגנים לא יהיה סביר; לכן, סביר יותר שההתארגנות תהיה סבירה יותר.
ביולוגים מקבצים תהליכים אלופטריים לשתי קטגוריות. אם כמה מבני מין עוברים לאזור גיאוגרפי חדש, זה נקרא פיזור. אם נוצר מצב טבעי לחלוקה פיזית של אורגניזמים, זה נקרא עקיפה.
מדענים תיעדו מקרים רבים של התגלמות אלופטרית המתרחשת. לדוגמה, לאורך החוף המערבי של ארצות הברית, קיימים שני תת-מינים נפרדים של ינשופים מנומרים. לינשוף המנומר הצפוני יש הבדלים גנטיים ופנוטיפיים מקרובו הקרוב, הינשוף המנומר המקסיקני, שחי בדרום (איור\(\PageIndex{2}\)). הסיבה להפרדתם הראשונית אינה ברורה, אך יתכן שהיא נגרמה על ידי הקרחונים של עידן הקרח שחילקו אוכלוסייה ראשונית לשניים. 1
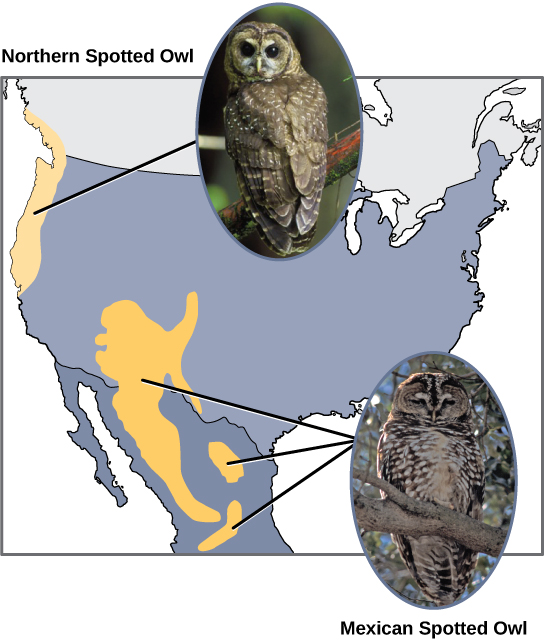
בנוסף, מדענים גילו שככל שהמרחק בין שתי קבוצות שפעם היו אותו מין, כך גדל הסיכוי להתרחשות התגלמות. זה נראה הגיוני מכיוון שככל שהמרחק גדל, סביר להניח שלגורמים הסביבתיים השונים יהיה פחות משותף מאשר מיקומים בסמיכות. קחו בחשבון את שני הינשופים; בצפון האקלים קריר יותר מאשר בדרום; סוגי האורגניזמים האחרים בכל מערכת אקולוגית נבדלים זה מזה, וכך גם התנהגויותיהם והרגליהם; כמו כן, הרגלי הציד ובחירות הטרף של הינשופים בדרום משתנים מהצפוניים. שונות זו יכולה להוביל להבדלים מתפתחים בינשופים, ועם הזמן סביר להניח שיתרחשו התגלמות אלא אם כן זרימת הגנים בין האוכלוסיות תשוחזר.
במקרים מסוימים, אוכלוסייה של מין אחד מתפזרת בכל אזור, וכל אחד מוצא נישה או בית גידול מבודד. עם הזמן, הדרישות המגוונות של אורח חייהם החדש מובילות לאירועי ייחוס מרובים שמקורם במין יחיד, הנקרא קרינה אדפטיבית. מנקודת מוצא אחת מתפתחות התאמות רבות הגורמות למין להקרין למספר חדשים. ארכיפלג איים כמו איי הוואי מספקים הקשר אידיאלי לאירועי קרינה אדפטיביים מכיוון שמים מקיפים כל אי, מה שמוביל לבידוד גיאוגרפי של אורגניזמים רבים (איור\(\PageIndex{3}\)). זוחל הדבש ההוואי ממחיש דוגמה אחת לקרינה אדפטיבית. ממין בודד, הנקרא המין המייסד, התפתחו מינים רבים, כולל השמונה המוצגים באיור\(\PageIndex{3}\).
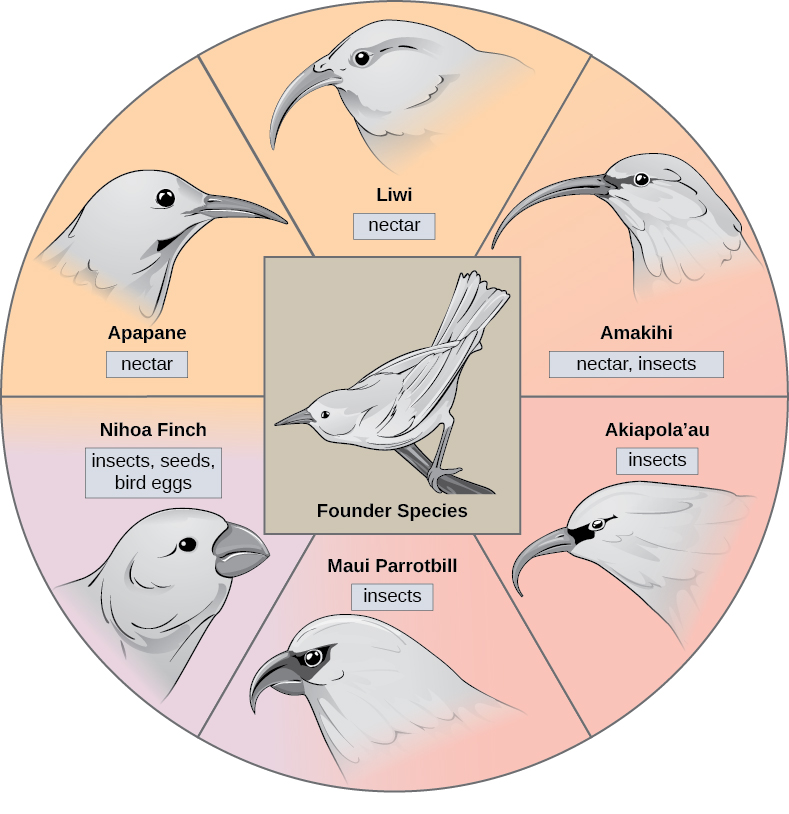
שימו לב להבדלים במקור המינים באיור. \(\PageIndex{3}\) שינוי בשונות הגנטית של מקור בתגובה לברירה הטבעית המבוססת על מקורות מזון ספציפיים בכל בית גידול חדש הוביל לאבולוציה של מקור אחר המתאים למקור המזון הספציפי. לציפורים אוכלות הפירות והזרעים יש מקור עבה וחזק יותר המתאים לשבירת אגוזים קשים. לציפורים אוכלות הצוף יש מקור ארוך לטבול בפרחים כדי להגיע לצוף שלהן. לציפורים אוכלות החרקים יש מקור כמו חרבות, המתאים לדקירת חרקים ושידודם. החוחיות של דרווין הן דוגמה נוספת שנחקרה היטב לקרינה אדפטיבית בארכיפלג.
ייחוס ללא הפרדה גיאוגרפית
האם יכולה להתרחש סטייה אם אין מחסומים פיזיים להפרדת פרטים הממשיכים לחיות ולהתרבות באותו בית גידול? הוצעו ונחקרו מספר מנגנונים לייחוס סימפטרי.
צורה אחת של ייחוס סימפטרי יכולה להתחיל בשגיאה כרומוזומלית במהלך מיוזה או היווצרות של אדם היברידי עם יותר מדי כרומוזומים. פוליפלואידיה היא מצב שבו לתא, או לאורגניזם, יש קבוצה נוספת, או קבוצות, של כרומוזומים. מדענים זיהו שני סוגים עיקריים של פוליפלואידיות שיכולים להוביל לבידוד הרבייה של אדם במצב פוליפלואידי. במקרים מסוימים לאדם פוליפלואידי יהיו שתי קבוצות שלמות או יותר של כרומוזומים מהמין שלו במצב הנקרא אוטופוליפלואידיה (איור). \(\PageIndex{4}\) הקידומת "אוטומטי" פירושה עצמי, ולכן המונח פירושו כרומוזומים מרובים מהמין של עצמו. פוליפלואידיה נובעת משגיאה במיוזה שבה כל הכרומוזומים עוברים לתא אחד במקום להיפרד.
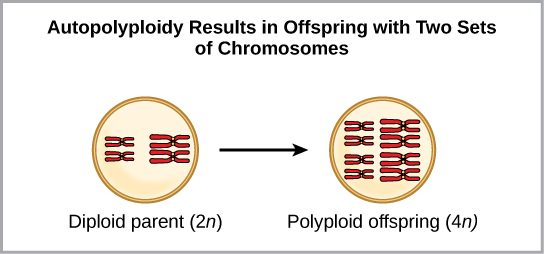
לדוגמה, אם מין צמחי עם 2 נ = 6 מייצר גמטות אוטופוליפלואידיות שהן גם דיפלואידיות (2 נ = 6, כשהן צריכות להיות נ = 3), לגמטות יש כעת כפליים כרומוזומים ממה שהיו צריכים להיות. גמטות חדשות אלה לא יהיו תואמות את הגמטות הרגילות המיוצרות על ידי מין צמחי זה. אבל הם יכלו להאביק את עצמם או להתרבות עם צמחים אוטופוליפלואידים אחרים עם גמטות בעלות אותו מספר דיפלואידי. בדרך זו, התגלמות סימפטרית יכולה להתרחש במהירות על ידי יצירת צאצאים עם 4 n הנקראים טטרפלואיד. אנשים אלה יוכלו מיד להתרבות רק עם אלה מהסוג החדש הזה ולא עם אלה של המין הקדמוני. הצורה האחרת של פוליפלואידיה מתרחשת כאשר פרטים משני מינים שונים מתרבים ויוצרים צאצא בר-קיימא הנקרא אלופוליפלואיד. הקידומת "allo" פירושה "אחר" (נזכר מאלופטרי); לפיכך, אלופוליפלואיד מתרחש כאשר גמטות משני מינים שונים מתאחדים. איור \(\PageIndex{5}\) ממחיש דרך אפשרית אחת שבה יכולה להיווצר אלופוליפלואידיה. שימו לב איך זה לוקח שני דורות, או שני פעולות רבייה, לפני התוצאות ההיברידיות הפוריות הקיימות.
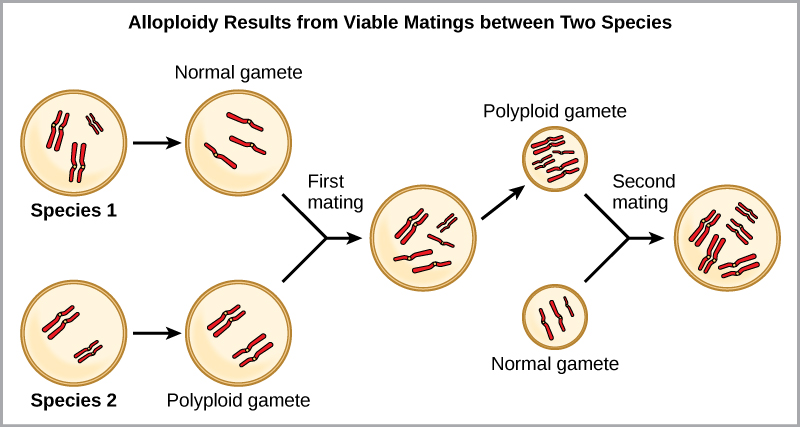
הצורות המעובדות של צמחי חיטה, כותנה וטבק הן כולן אלופוליפלואידים. למרות שפוליפלואידיה מתרחשת מדי פעם בבעלי חיים, רוב החריגות הכרומוזומליות בבעלי חיים הן קטלניות; זה מתרחש לרוב בצמחים. מדענים גילו שיותר מ -1/2 מכל מיני הצמחים שנחקרו מתייחסים למין שהתפתח באמצעות פוליפלואידיות.
ייחוס סימפטרי עשוי להתרחש גם בדרכים אחרות מלבד פוליפלואידיות. לדוגמה, דמיין מין דגים שחי באגם. ככל שהאוכלוסייה גדלה, התחרות על מזון גדלה גם היא. בלחץ למצוא מזון, נניח שלקבוצה של דגים אלה הייתה הגמישות הגנטית לגלות ולהאכיל משאב אחר שלא היה בשימוש על ידי הדגים האחרים. מה אם מקור המזון החדש הזה נמצא בעומק אחר של האגם? עם הזמן, אלה הניזונים ממקור המזון השני היו מתקשרים יותר זה עם זה מאשר הדגים האחרים; לכן הם היו מתרבים גם יחד. צאצאים של דגים אלה יתנהגו ככל הנראה כהוריהם ויאכילו ויחיו באותו אזור, וישאירו אותם נפרדים מהאוכלוסייה המקורית. אם קבוצת דגים זו תמשיך להישאר נפרדת מהאוכלוסייה הראשונה, בסופו של דבר עלולה להתרחש התגלמות סימפטרית ככל שיצטברו הבדלים גנטיים נוספים ביניהם.
תרחיש זה אכן מתרחש בטבע, וכך גם אחרים המובילים לבידוד רבייה. מקום אחד כזה הוא אגם ויקטוריה באפריקה, המפורסם בזכות ייחודו הסימפטרי של דגי ציקלידים. חוקרים מצאו מאות אירועי ייחוס סימפטריים בדגים אלה, אשר לא התרחשו רק במספר רב, אלא גם לאורך זמן קצר. איור \(\PageIndex{6}\) מציג סוג זה של ייחוס בקרב אוכלוסיית דגי ציקלידים בניקרגואה. באזור זה, שני סוגים של ציקלידים חיים באותו מיקום גיאוגרפי; עם זאת, יש להם מורפולוגיות שונות המאפשרות להם לאכול מקורות מזון שונים.
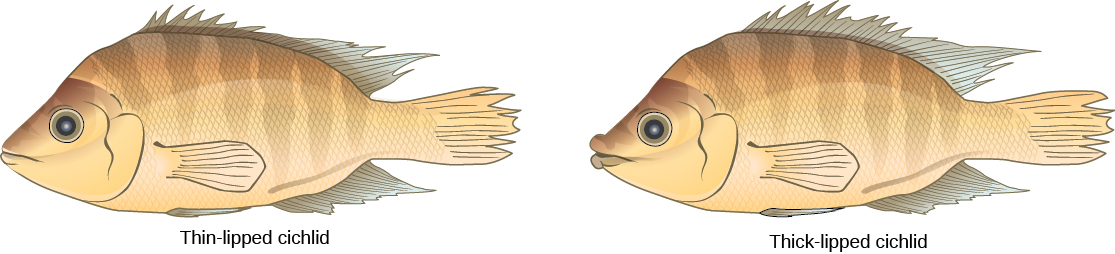
לבסוף, דוגמה מתועדת היטב להתייחסות סימפטרית מתמשכת התרחשה בזבוב רימת התפוחים, Rhagoletis pomonella, שהתעוררה כאוכלוסייה מבודדת מתישהו לאחר הכנסת התפוח לצפון אמריקה. אוכלוסיית הילידים של זבובים הניזונים ממיני עוזרד והיא ספציפית למארח: היא פוגעת רק בעצי עוזרד. חשוב לציין, הוא גם משתמש בעצים כמיקום להיפגש להזדווגות. ההשערה היא כי באמצעות מוטציה או טעות התנהגותית, זבובים קפצו מארחים ונפגשו והזדווגו בעצי תפוח, ולאחר מכן הטילו את ביציהם בפרי תפוח. הצאצאים התבגרו ושמרו על העדפתם לעצי התפוח וחילקו למעשה את האוכלוסייה המקורית לשתי אוכלוסיות חדשות המופרדות על ידי מינים מארחים, לא לפי גיאוגרפיה. הקפיצה המארחת התרחשה במאה התשע עשרה, אך כיום ישנם הבדלים ניתנים למדידה בין שתי אוכלוסיות הזבובים. סביר להניח כי סגוליות המארח של טפילים באופן כללי היא גורם שכיח לייחוס סימפטרי.
סיכום המדור
התגלמות מתרחשת בשני מסלולים עיקריים: הפרדה גיאוגרפית (ייחוס אלופטרי) ובאמצעות מנגנונים המתרחשים בתוך בית גידול משותף (מין סימפטרי). שני המסלולים מכריחים בידוד רבייה בין אוכלוסיות. ייחוס סימפטרי יכול להתרחש באמצעות טעויות במיוזה היוצרות גמטות עם כרומוזומים נוספים, הנקראים פוליפלואידיה. אוטופוליפלואידיה מתרחשת בתוך מין בודד, ואילו אלופוליפלואידיה מתרחשת בגלל הזדווגות בין מינים קרובים. ברגע שהאוכלוסיות מבודדות, יכולה להתרחש סטייה אבולוציונית המובילה לאבולוציה של תכונות בידוד רבייה המונעות הכלאה אם שתי האוכלוסיות יתאחדו שוב. הכדאיות המופחתת של צאצאים היברידיים לאחר תקופת בידוד צפויה לבחור במנגנוני בידוד מובנים חזקים יותר.
הערות שוליים
- 1 Courtney, S.P., et al, "הערכה מדעית של מעמדו של הינשוף המנוקד הצפוני", המכון למערכות אקולוגיות בר קיימא (2004), פורטלנד, OR.
רשימת מילים
- קרינה אדפטיבית
- מין כאשר מין אחד מקרין החוצה ויוצר כמה מינים אחרים
- ייחוס אלופטרי
- סוג המתרחש באמצעות הפרדה גיאוגרפית
- פיזור
- סוג אלופטרי המתרחש כאשר כמה מבני מין עוברים לאזור גיאוגרפי חדש
- ייחוס
- היווצרות של מין חדש
- ייחוס סימפטרי
- סוג המתרחש באותו מרחב גיאוגרפי
- משנה
- סוג אלופטרי המתרחש כאשר משהו בסביבה מפריד אורגניזמים מאותו מין לקבוצות נפרדות