
9.2: שכפול DNA
- Page ID
- 208870

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)כאשר תא מתחלק, חשוב שכל תא בת יקבל עותק זהה של ה- DNA. זה מושג על ידי תהליך שכפול ה- DNA. שכפול ה- DNA מתרחש בשלב הסינתזה, או שלב S, של מחזור התא, לפני שהתא נכנס למיטוזה או למיוזה.
הבהרת מבנה הסליל הכפול סיפקה רמז לאופן העתקת ה- DNA. נזכיר כי נוקלאוטידים אדנין מזדווגים עם נוקלאוטידים של תימין, וציטוסין עם גואנין. המשמעות היא ששני הגדילים משלימים זה את זה. לדוגמה, לגדיל DNA עם רצף נוקלאוטידים של AGTCATGA יהיה גדיל משלים עם הרצף TCAGTACT (איור). \(\PageIndex{1}\)
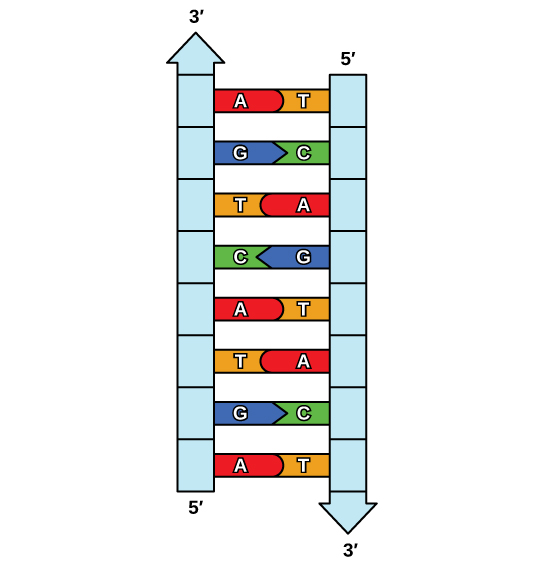
בגלל ההשלמה של שני הגדילים, בעל גדיל אחד פירושו שניתן ליצור מחדש את הגדיל השני. מודל זה לשכפול מציע ששני הגדילים של הסליל הכפול נפרדים במהלך השכפול, וכל גדיל משמש כתבנית שממנה מועתק הגדיל המשלים החדש (איור\(\PageIndex{2}\)).
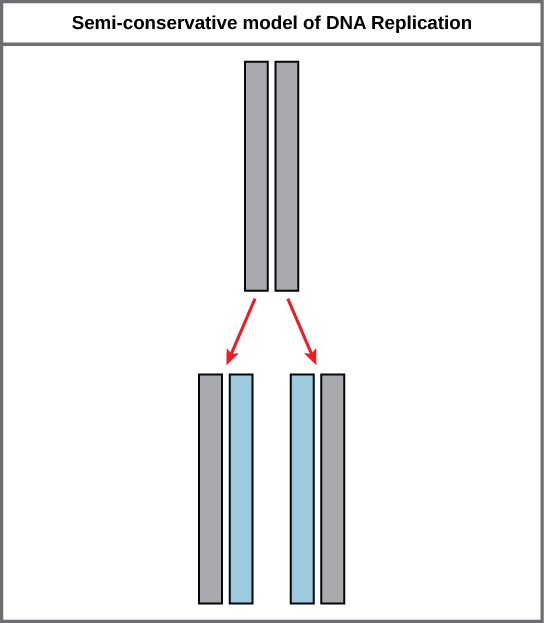
במהלך שכפול ה- DNA, כל אחד משני הגדילים המרכיבים את הסליל הכפול משמש כתבנית שממנה מועתקים גדילים חדשים. הגדיל החדש יהיה משלים לחוט ההורי או "הישן". כל גדיל כפול חדש מורכב מחוט הורי אחד וחוט בת חדש אחד. זה ידוע בשם שכפול שמרני למחצה. כאשר נוצרים שני עותקי DNA, יש להם רצף זהה של בסיסי נוקלאוטידים והם מחולקים באופן שווה לשני תאי בת.
שכפול DNA באיקריוטים
מכיוון שגנום אוקריוטי מורכב מאוד, שכפול ה- DNA הוא תהליך מסובך מאוד הכולל מספר אנזימים וחלבונים אחרים. זה מתרחש בשלושה שלבים עיקריים: התחלה, התארכות וסיום.
נזכיר כי DNA אוקריוטי קשור לחלבונים המכונים היסטונים ליצירת מבנים הנקראים נוקלאוזומים. במהלך ההתחלה, ה-DNA נגיש לחלבונים ולאנזימים המעורבים בתהליך השכפול. כיצד מכונות השכפול יודעות היכן להתחיל בסליל הכפול של ה-DNA? מסתבר שיש רצפי נוקלאוטידים ספציפיים הנקראים מקורות שכפול שבהם מתחיל השכפול. חלבונים מסוימים נקשרים למקור השכפול בעוד אנזים הנקרא helicase מתפרק ופותח את סליל ה- DNA. כאשר ה-DNA נפתח, נוצרים מבנים בצורת Y הנקראים מזלגות שכפול (איור\(\PageIndex{3}\)). שני מזלגות שכפול נוצרים במקור השכפול, ואלה מורחבים לשני הכיוונים ככל שהשכפול מתקדם. ישנם מקורות שכפול מרובים בכרומוזום האוקריוטי, כך ששכפול יכול להתרחש בו זמנית מכמה מקומות בגנום.
במהלך התארכות, אנזים הנקרא DNA פולימראז מוסיף נוקלאוטידים של DNA לקצה 3' של התבנית. מכיוון ש- DNA פולימראז יכול להוסיף נוקלאוטידים חדשים רק בקצה עמוד השדרה, רצף פריימר, המספק נקודת התחלה זו, מתווסף עם נוקלאוטידים משלימים של RNA. פריימר זה מוסר מאוחר יותר, והנוקלאוטידים מוחלפים בנוקלאוטידים של DNA. גדיל אחד, המשלים את גדיל ה-DNA ההורי, מסונתז ברציפות לכיוון מזלג השכפול כך שהפולימראז יכול להוסיף נוקלאוטידים בכיוון זה. גדיל מסונתז זה ברציפות ידוע כגדיל המוביל. מכיוון ש- DNA פולימראז יכול לסנתז DNA רק בכיוון 5 'עד 3', הגדיל החדש השני מורכב בחלקים קצרים הנקראים שברי Okazaki. שברי Okazaki דורשים כל אחד פריימר עשוי RNA כדי להתחיל את הסינתזה. הגדיל עם שברי אוקאזאקי ידוע כגדיל הפיגור. ככל שהסינתזה מתקדמת, אנזים מסיר את פריימר ה-RNA, אשר מוחלף לאחר מכן בנוקלאוטידים של DNA, והפערים בין השברים נאטמים על ידי אנזים הנקרא DNA ligase.
ניתן לסכם את תהליך שכפול ה- DNA באופן הבא:
- ה- DNA מתפרק במקור השכפול.
- בסיסים חדשים מתווספים לחוטי ההורים המשלימים. גדיל חדש אחד מיוצר ברציפות, ואילו הגדיל השני מיוצר בחתיכות.
- פריימרים מוסרים, נוקלאוטידים חדשים של DNA מונחים במקום הפריימרים ועמוד השדרה אטום על ידי ליגאז DNA.
חיבור אמנות
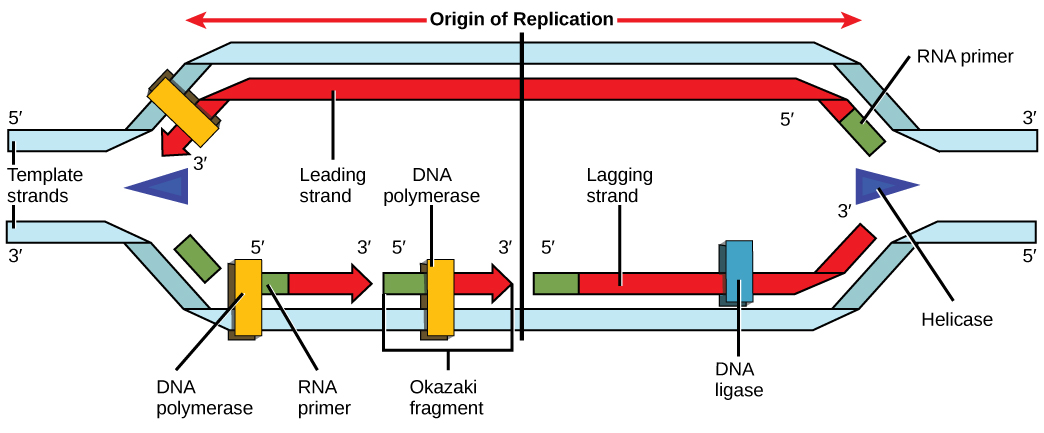
אתה מבודד זן תאים שבו החיבור של שברי Okazaki נפגע וחושד שהתרחשה מוטציה באנזים שנמצא במזלג השכפול. איזה אנזים צפוי לעבור מוטציה?
שכפול טלומרים
מכיוון שכרומוזומים אוקריוטיים הם ליניאריים, שכפול ה- DNA מגיע לסוף קו בכרומוזומים אוקריוטיים. כפי שלמדת, האנזים DNA פולימראז יכול להוסיף נוקלאוטידים בכיוון אחד בלבד. בגדיל המוביל הסינתזה נמשכת עד שמגיעים לסוף הכרומוזום; עם זאת, בגדיל הפיגור אין מקום לייצור פריימר להעתקת שבר ה- DNA בסוף הכרומוזום. זה מהווה בעיה עבור התא מכיוון שהקצוות נשארים לא מזווגים, ועם הזמן הקצוות הללו מתקצרים בהדרגה ככל שהתאים ממשיכים להתחלק. קצות הכרומוזומים הליניאריים ידועים בשם טלומרים, שיש להם רצפים חוזרים שאינם מקודדים לגן מסוים. כתוצאה מכך, הטלומרים מתקצרים בכל סיבוב של שכפול DNA במקום גנים. לדוגמה, בבני אדם, רצף של שישה זוגות בסיסים, TTAGGG, חוזר על עצמו 100 עד 1000 פעמים. גילוי האנזים טלומראז (איור\(\PageIndex{4}\)) עזר בהבנת האופן שבו נשמרים קצוות הכרומוזומים. הטלומראז מתחבר לקצה הכרומוזום, ובסיסים משלימים לתבנית ה- RNA מתווספים בקצה גדיל ה- DNA. ברגע שתבנית הגדיל המפגר מוארכת מספיק, DNA פולימראז יכול כעת להוסיף נוקלאוטידים המשלימים את קצות הכרומוזומים. לפיכך, קצות הכרומוזומים משוכפלים.
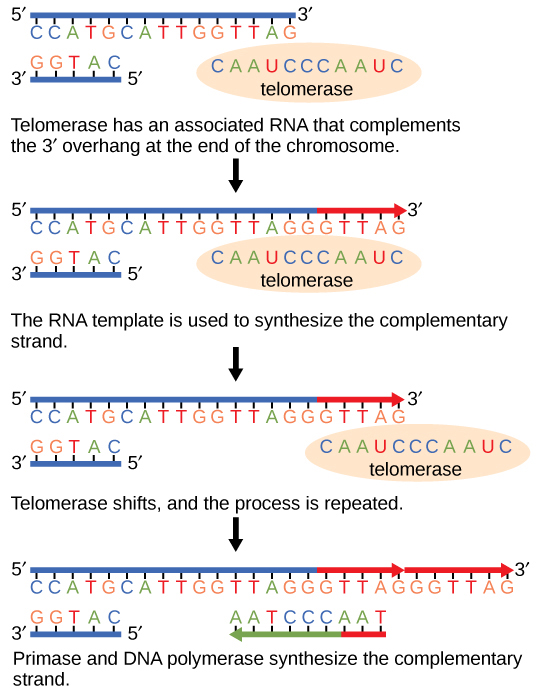
טלומראז נמצא בדרך כלל פעיל בתאי נבט, בתאי גזע בוגרים ובכמה תאים סרטניים. על גילוי הטלומראז ופעולתו, אליזבת בלקברן (איור\(\PageIndex{5}\)) קיבלה את פרס נובל לרפואה ופיזיולוגיה בשנת 2009.

טלומראז אינו פעיל בתאים סומטיים בוגרים. תאים סומטיים בוגרים שעוברים חלוקת תאים ממשיכים לקצר את הטלומרים שלהם. זה בעצם אומר שקיצור הטלומרים קשור להזדקנות. בשנת 2010, מדענים גילו כי טלומראז יכול להפוך כמה מצבים הקשורים לגיל בעכברים, וייתכן שיש לכך פוטנציאל ברפואה רגנרטיבית. 1 עכברים חסרי טלומראז שימשו במחקרים אלה; לעכברים אלה יש ניוון רקמות, דלדול תאי גזע, כשל במערכת האיברים ותגובות פגיעה ברקמות לקויות. הפעלה מחדש של טלומראז בעכברים אלה גרמה להארכת הטלומרים, להפחתת נזק ל- DNA, ניוון עצבי הפוך ושיפור תפקוד האשכים, הטחול והמעיים. לפיכך, להפעלה מחדש של הטלומרים עשוי להיות פוטנציאל לטיפול במחלות הקשורות לגיל בבני אדם.
שכפול DNA בפרוקריוטים
נזכיר כי הכרומוזום הפרוקריוטי הוא מולקולה מעגלית בעלת מבנה סליל פחות נרחב מאשר כרומוזומים אוקריוטיים. הכרומוזום האוקריוטי הוא ליניארי ומפותל מאוד סביב חלבונים. אמנם ישנם קווי דמיון רבים בתהליך שכפול ה- DNA, אך הבדלים מבניים אלה מחייבים הבדלים מסוימים בתהליך שכפול ה- DNA בשתי צורות החיים הללו.
שכפול ה- DNA נחקר היטב בפרוקריוטים, בעיקר בגלל גודלו הקטן של הגנום ומספר רב של גרסאות זמינות. ל-Escherichia coli יש 4.6 מיליון זוגות בסיסים בכרומוזום מעגלי יחיד, וכל זה משתכפל תוך כ-42 דקות, החל ממקור שכפול יחיד וממשיך סביב הכרומוזום לשני הכיוונים. משמעות הדבר היא כי כ 1000 נוקלאוטידים מתווספים לשנייה. התהליך הוא הרבה יותר מהיר מאשר באוקריוטים. הטבלה \(\PageIndex{1}\) מסכמת את ההבדלים בין שכפולים פרוקריוטיים לאוקריוטיים.
| רכוש | פרוקריוטים | אוקריוטים |
|---|---|---|
| מקור השכפול | רווק | מרובה |
| קצב השכפול | 1000 נוקלאוטידים/שניות | 50 עד 100 נוקלאוטידים/שניות |
| מבנה הכרומוזום | מחזורי | ליניארי |
| טלומראז | לא נוכח | הווה |
מושג בפעולה
לחץ על הדרכה בנושא שכפול DNA.
תיקון DNA
פולימראז DNA יכול לעשות טעויות בעת הוספת נוקלאוטידים. הוא עורך את ה- DNA על ידי הגהה של כל בסיס שנוסף לאחרונה. בסיסים שגויים מוסרים ומוחלפים על ידי הבסיס הנכון, ולאחר מכן פילמור ממשיך (איור \(\PageIndex{6}\) א). רוב הטעויות מתוקנות במהלך השכפול, אם כי כאשר זה לא קורה, מנגנון תיקון חוסר ההתאמה מופעל. אנזימי תיקון חוסר התאמה מזהים את הבסיס המשולב בצורה לא נכונה ומוציאים אותו מה-DNA, ומחליפים אותו בבסיס הנכון (איור \(\PageIndex{6}\) ב). בסוג אחר של תיקון, תיקון כריתת נוקלאוטידים, הגדיל הכפול של ה-DNA מתפרק ומופרד, הבסיסים השגויים מוסרים יחד עם כמה בסיסים בקצה 5' ו-3', ואלה מוחלפים בהעתקת התבנית בעזרת DNA פולימראז (איור ג). \(\PageIndex{6}\) תיקון כריתת נוקלאוטידים חשוב במיוחד בתיקון דימרים של תימין, הנגרמים בעיקר מאור אולטרה סגול. בדימר תימין, שני נוקלאוטידים של תימין הסמוכים זה לזה על גדיל אחד קשורים זה לזה באופן קוולנטי ולא הבסיסים המשלימים שלהם. אם הדימר לא יוסר ומתוקן הוא יוביל למוטציה. אנשים עם פגמים בגנים לתיקון כריתת הנוקלאוטידים שלהם מראים רגישות קיצונית לאור השמש ומפתחים סרטן עור בשלב מוקדם של חייהם.
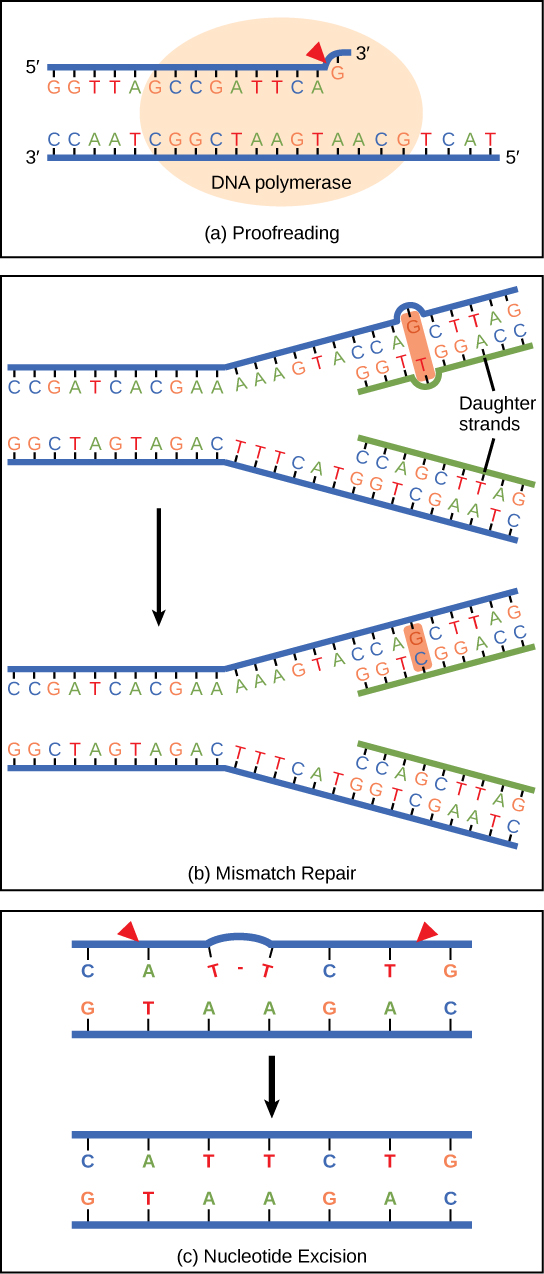
רוב הטעויות מתוקנות; אם הן לא, הן עלולות לגרום למוטציה - המוגדרת כשינוי קבוע ברצף ה- DNA. מוטציות בגנים לתיקון עלולות להוביל לתוצאות חמורות כמו סרטן.
סיכום
DNA משוכפל בשיטה שמרנית למחצה שבה כל אחד משני גדילי ה-DNA ההוריים משמש כתבנית לסינתזה של DNA חדש. לאחר השכפול, לכל DNA יש גדיל הורי או "ישן" אחד, ובת אחת או גדיל "חדש".
שכפול באאוקריוטים מתחיל במספר מקורות שכפול, בעוד שכפול בפרוקריוטים מתחיל ממקור שכפול יחיד. ה- DNA נפתח באנזימים, וכתוצאה מכך נוצר מזלג השכפול. Primase מסנתז פריימר RNA ליזום סינתזה על ידי DNA פולימראז, שיכול להוסיף נוקלאוטידים בכיוון אחד בלבד. גדיל אחד מסונתז ברציפות לכיוון מזלג השכפול; זה נקרא הגדיל המוביל. הגדיל השני מסונתז בכיוון הרחק ממזלג השכפול, בקטעי DNA קצרים המכונים שברי אוקזאקי. גדיל זה ידוע כגדיל הפיגור. לאחר השלמת השכפול, הפריימרים של ה- RNA מוחלפים בנוקלאוטידים של DNA וה- DNA אטום בליגאז DNA.
קצות הכרומוזומים האוקריוטיים מהווים בעיה, מכיוון שפולימראז אינו מסוגל להאריך אותם ללא פריימר. טלומראז, אנזים בעל תבנית RNA מובנית, מרחיב את הקצוות על ידי העתקת תבנית ה- RNA והארכת קצה אחד של הכרומוזום. לאחר מכן DNA פולימראז יכול להאריך את ה- DNA באמצעות הפריימר. באופן זה, קצות הכרומוזומים מוגנים. לתאים יש מנגנונים לתיקון DNA כאשר הוא ניזוק או נעשות שגיאות בשכפול. מנגנונים אלה כוללים תיקון חוסר התאמה להחלפת נוקלאוטידים המשויכים לבסיס לא משלים ותיקון כריתת נוקלאוטידים, המסיר בסיסים שניזוקו כגון דימרים של תימין.
חיבורי אמנות
איור\(\PageIndex{3}\): אתה מבודד זן תאים שבו החיבור של שברי אוקאזאקי נפגע וחושד שהתרחשה מוטציה באנזים שנמצא במזלג השכפול. איזה אנזים צפוי לעבור מוטציה?
- תשובה
-
ליגאז, כאשר אנזים זה מחבר יחד שברי אוקזאקי.
הערות שוליים
- 1 מריאלה ג'סקליוף, et al., "הפעלה מחדש של טלומראז הופכת את התנוונות הרקמות בעכברים חסרי טלומראז מיושנים", Nature, 469 (2011): 102-7.
רשימת מילים
- ליגאז DNA
- האנזים המזרז את החיבור של שברי DNA יחד
- פולימראז DNA
- אנזים המסנתז גדיל חדש של DNA המשלים לגדיל תבנית
- הליקאז
- אנזים שעוזר לפתוח את סליל ה-DNA במהלך שכפול ה-DNA על ידי שבירת קשרי המימן
- גדיל בפיגור
- במהלך שכפול של גדיל 3' עד 5', הגדיל המשוכפל בשברים קצרים והרחק ממזלג השכפול
- גדיל מוביל
- הגדיל המסונתז ברציפות בכיוון 5 'עד 3' המסונתז לכיוון מזלג השכפול
- תיקון אי התאמה
- צורה של תיקון DNA שבו נוקלאוטידים לא משלימים מזוהים, נכרתים ומוחלפים בנוקלאוטידים נכונים
- מוטציה
- וריאציה קבועה ברצף הנוקלאוטידים של גנום
- תיקון כריתת נוקלאוטיד
- צורה של תיקון DNA שבו מולקולת ה-DNA נפרשת ומופרדת באזור הנזק לנוקלאוטיד, הנוקלאוטידים הפגועים מוסרים ומוחלפים בנוקלאוטידים חדשים באמצעות הגדיל המשלים, וגדיל ה-DNA נסגר מחדש ומאפשר להצטרף מחדש להשלמתו
- שברי אוקזאקי
- שברי ה- DNA המסונתזים במתיחות קצרות על הגדיל המפגר
- צבע יסוד
- קטע קצר של נוקלאוטידים של RNA הנדרש ליזום שכפול ולאפשר ל-DNA פולימראז להיקשר ולהתחיל בשכפול
- מזלג שכפול
- המבנה בצורת Y שנוצר במהלך תחילת השכפול
- שכפול חצי שמרני
- השיטה המשמשת לשכפול DNA שבה מופרדת המולקולה הדו-גדילית וכל גדיל משמש כתבנית לסינתזה של גדיל חדש, כך שמולקולות ה-DNA המתקבלות מורכבות מחוט חדש אחד של נוקלאוטידים וגדיל ישן אחד של נוקלאוטידים
- טלומראז
- אנזים המכיל חלק קטליטי ותבנית RNA מובנית; הוא מתפקד לשמירה על טלומרים בקצות הכרומוזומים
- טלומרים
- ה- DNA בקצה הכרומוזומים הליניאריים