
23.1: מקורות אוקריוטיים
- Page ID
- 206552

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)מיומנויות לפיתוח
- רשום את המאפיינים המאחדים של אוקריוטים
- תאר את מה שמדענים יודעים על מקורות האיקריוטים בהתבסס על האב הקדמון המשותף האחרון
- הסבר את התיאוריה האנדוסימביוטית
יצורים חיים מתחלקים לשלוש קבוצות גדולות: ארכאה, חיידקים ואוקריה. לשניים הראשונים יש תאים פרוקריוטים, והשלישי מכיל את כל האיקריוטים. קיים תיעוד מאובנים דליל יחסית שיעזור להבחין כיצד נראו החברים הראשונים בכל אחת מהשושלות הללו, כך שייתכן שכל האירועים שהובילו לאב הקדמון המשותף האחרון של האוקריוטים הקיימים יישארו לא ידועים. עם זאת, ביולוגיה השוואתית של אורגניזמים קיימים ותיעוד המאובנים המצומצם מספקים תובנה מסוימת על ההיסטוריה של אוקריה.
נראה כי המאובנים המוקדמים ביותר שנמצאו הם חיידקים, ככל הנראה ציאנובקטריה. הם בני כ-3.5 מיליארד שנים וניתנים לזיהוי בגלל המבנה המורכב יחסית שלהם, ועבור פרוקריוטים, תאים גדולים יחסית. לרוב הפרוקריוטים האחרים יש תאים קטנים, בגודל של 1 או 2 מיקרומטר, ויהיה קשה לבחור אותם כמאובנים. לרוב האיקריוטים החיים יש תאים בגודל 10 מיקרומטר ומעלה. מבנים בגודל זה, שעשויים להיות מאובנים, מופיעים בתיעוד הגיאולוגי לפני כ -2.1 מיליארד שנה.
מאפייני האוקריוטים
נתונים ממאובנים אלה הובילו ביולוגים השוואתיים למסקנה שאאוקריוטים חיים הם כולם צאצאים של אב קדמון משותף אחד. מיפוי המאפיינים שנמצאו בכל קבוצות האיקריוטים העיקריות מגלה כי המאפיינים הבאים בוודאי היו קיימים באב הקדמון המשותף האחרון, מכיוון שמאפיינים אלה קיימים לפחות בחלק מחברי כל שושלת מרכזית.
- תאים עם גרעינים מוקפים במעטפת גרעינית עם נקבוביות גרעיניות. זהו המאפיין היחיד שהוא הכרחי ומספיק כדי להגדיר אורגניזם כאאוקריוט. לכל האיקריוטים הקיימים יש תאים עם גרעינים.
- מיטוכונדריה. לחלק מהאוקריוטים הקיימים יש שרידים מופחתים מאוד של המיטוכונדריה בתאים שלהם, בעוד שלחברים אחרים בשושלתם יש מיטוכונדריה "טיפוסית".
- שלד המכיל את המרכיבים המבניים והתנועתיים הנקראים מיקרופילמנטים אקטין ומיקרוטובולים. לכל האיקריוטים הקיימים יש את היסודות הציטו-שלדיים הללו.
- Flagella ו cilia, אברונים הקשורים לתנועתיות התא. לחלק מהאוקריוטים הקיימים חסרים דגלים ו/או ריסים, אך הם צאצאים מאבות קדומים שהיו ברשותם.
- כרומוזומים, שכל אחד מהם מורכב ממולקולת DNA ליניארית המפותלת סביב חלבונים בסיסיים (אלקליין) הנקראים היסטונים. האיקריוטים המעטים עם כרומוזומים חסרי היסטונים התפתחו בבירור מאבות קדומים שהיו להם.
- מיטוזיס, תהליך של חלוקה גרעינית שבו כרומוזומים משוכפלים מחולקים ומופרדים באמצעות אלמנטים של שלד הציטוס. מיטוזיס קיימת באופן אוניברסלי באוקריוטים.
- מין, תהליך של רקומבינציה גנטית ייחודית לאאוקריוטים שבו גרעינים דיפלואידים בשלב אחד של מחזור החיים עוברים מיוזה כדי להניב גרעינים הפלואידים וקריוגמיה שלאחר מכן, שלב שבו שני גרעינים הפלואידים מתמזגים יחד ליצירת גרעין זיגוטה דיפלואידי.
- לחברים בכל השושלות הגדולות יש קירות תאים, וייתכן שיהיה סביר להסיק שהאב הקדמון המשותף האחרון יכול ליצור קירות תאים בשלב כלשהו של מחזור חייו. עם זאת, לא ידוע מספיק על דפנות התא של האוקריוטים והתפתחותם כדי לדעת כמה הומולוגיה קיימת ביניהם. אם האב הקדמון המשותף האחרון יכול היה ליצור קירות תאים, ברור שיכולת זו בוודאי אבדה בקבוצות רבות.
אנדוסימביוזה והתפתחות האיקריוטים
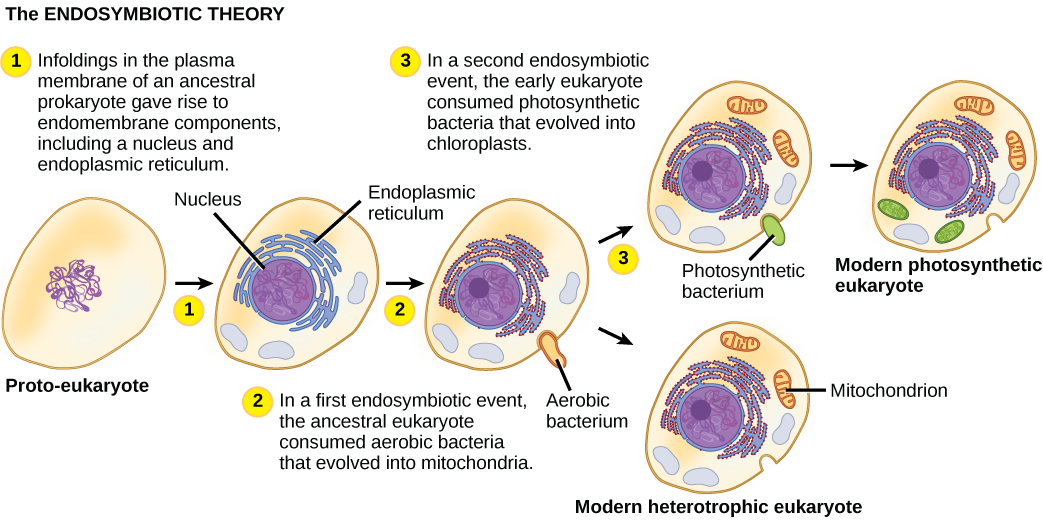
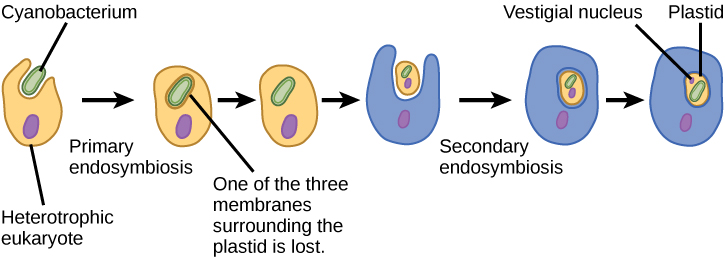
על מנת להבין אורגניזמים אוקריוטיים במלואם, יש להבין שכל האיקריוטים הקיימים הם צאצאים של אורגניזם כימרי שהיה מורכב של תא מארח והתאים של אלפא-פרוטאובקטריום ש"התגורר "בתוכו. נושא מרכזי זה במקור האיקריוטים ידוע בשם אנדוסימביוזה, תא אחד בולע אחר כך שהתא הנבלע שורד ושני התאים מרוויחים. במשך דורות רבים, מערכת יחסים סימביוטית יכולה לגרום לשני אורגניזמים התלויים זה בזה בצורה כה מוחלטת שאף אחד מהם לא יכול לשרוד בכוחות עצמו. אירועים אנדוסימביוטיים תרמו ככל הנראה למקורו של האב הקדמון המשותף האחרון של האיקריוטים של ימינו ולגיוון מאוחר יותר בשושלות מסוימות של אוקריוטים (איור). \(\PageIndex{4}\) לפני שמסבירים זאת עוד יותר, יש לקחת בחשבון את חילוף החומרים בפרוקריוטים.
מטבוליזם פרוקריוטי
תהליכים מטבוליים חשובים רבים התעוררו בפרוקריוטים, וחלקם, כגון קיבוע חנקן, לעולם אינם נמצאים באיקריוטים. תהליך הנשימה האירובית נמצא בכל השושלות העיקריות של האאוקריוטים, והוא מקומי במיטוכונדריה. נשימה אירובית נמצאת גם בשושלות רבות של פרוקריוטים, אך היא אינה קיימת בכולן, וצורות רבות של עדויות מצביעות על כך שפרוקריוטים אנאירוביים כאלה מעולם לא ביצעו נשימה אירובית וגם לא אבותיהם.
בעוד שהאטמוספירה של היום היא כחמישית חמצן מולקולרי (O 2), עדויות גיאולוגיות מראות כי במקור היה חסר O 2. ללא חמצן, נשימה אירובית לא הייתה צפויה, ויצורים חיים היו מסתמכים על תסיסה במקום זאת. בשלב מסוים לפני כן, לפני כ -3.5 מיליארד שנים, כמה פרוקריוטים החלו להשתמש באנרגיה מאור השמש כדי להניע תהליכים אנבוליים המפחיתים פחמן דו חמצני ליצירת תרכובות אורגניות. כלומר, הם פיתחו את היכולת לפוטוסינתזה. מימן, שמקורו במקורות שונים, נלכד באמצעות תגובות המופעלות על ידי אור להפחתת פחמן דו חמצני קבוע במחזור קלווין. קבוצת החיידקים הגראם-שליליים שהולידו ציאנובקטריה השתמשה במים כמקור המימן ושחררה את O 2 כתוצר פסולת.
בסופו של דבר, כמות החמצן הפוטוסינתטי שהצטברה בסביבות מסוימות לרמות שהיוו סיכון לאורגניזמים חיים, מכיוון שהיא עלולה לפגוע בתרכובות אורגניות רבות. התפתחו תהליכים מטבוליים שונים שהגנו על אורגניזמים מפני חמצן, שאחד מהם, נשימה אירובית, יצר גם רמות גבוהות של ATP. הוא הפך להיות נוכח בקרב פרוקריוטים, כולל בקבוצה שאנו מכנים כיום אלפא-פרוטאובקטריה. אורגניזמים שלא רכשו נשימה אירובית נאלצו להישאר בסביבות נטולות חמצן. במקור, סביר להניח שסביבות עשירות בחמצן היו ממוקמות סביב מקומות שבהם ציאנובקטריה הייתה פעילה, אך לפני כ -2 מיליארד שנה, עדויות גיאולוגיות מראות כי חמצן הצטבר לריכוזים גבוהים יותר באטמוספירה. רמות החמצן הדומות לרמות של ימינו עלו רק במהלך 700 מיליון השנים האחרונות.
נזכיר כי המאובנים הראשונים שלדעתנו הם אוקריוטים מתוארכים לגיל של כ -2 מיליארד שנה, ולכן הם הופיעו ככל שרמות החמצן עלו. כמו כן, זכור כי כל האיקריוטים הקיימים צאצאים מאב קדמון עם מיטוכונדריה. אברונים אלה נצפו לראשונה על ידי מיקרוסקופי אור בסוף המאה ה -19, שם נראה שהם מבנים בצורת תולעת משהו שנראו כאילו הם מסתובבים בתא. כמה משקיפים מוקדמים הציעו שהם עשויים להיות חיידקים החיים בתוך תאי מארח, אך השערות אלה נותרו לא ידועות או נדחות ברוב הקהילות המדעיות.
תיאוריה אנדוסימביוטית
ככל שהתפתחה ביולוגיה של התא במאה העשרים, התברר כי המיטוכונדריה הן האברונים האחראים לייצור ATP באמצעות נשימה אירובית. בשנות השישים פיתחה הביולוגית האמריקאית לין מרגוליס תיאוריה אנדוסימביוטית, הקובעת כי ייתכן שאוקריוטים היו תוצר של תא אחד הבולע אחר, אחד חי בתוך אחר, ומתפתח עם הזמן עד שהתאים הנפרדים כבר לא היו ניתנים לזיהוי ככאלה. בשנת 1967 הציגה מרגוליס עבודה חדשה על התיאוריה וביססה את ממצאיה באמצעות עדויות מיקרוביולוגיות. למרות שעבודתו של מרגוליס נתקלה בתחילה בהתנגדות, השערה זו של פעם מהפכנית מתקבלת כיום באופן נרחב (אך לא לגמרי), כאשר העבודה מתקדמת בחשיפת השלבים הכרוכים בתהליך אבולוציוני זה ושחקני המפתח המעורבים. עדיין נותר לגלות הרבה על מקורות התאים המרכיבים כעת את התאים בכל האיקריוטים החיים.
באופן כללי, התברר שרבים מהגנים הגרעיניים שלנו והמנגנון המולקולרי האחראי לשכפול וביטוי נראים קשורים קשר הדוק לאלה שבארכאה. מצד שני, האברונים המטבוליים והגנים האחראים לתהליכי קצירת אנרגיה רבים מקורם בחיידקים. נותר הרבה להבהיר כיצד התרחשה מערכת יחסים זו; זה ממשיך להיות תחום גילוי מרגש בביולוגיה. לדוגמה, לא ידוע אם האירוע האנדוסימביוטי שהוביל למיטוכונדריה התרחש לפני או אחרי שלתא המארח היה גרעין. אורגניזמים כאלה יהיו בין המבשרים שנכחדו של האב הקדמון המשותף האחרון של האיקריוטים.
מיטוכונדריה
אחד המאפיינים העיקריים המבדילים בין פרוקריוטים לאאוקריוטים הוא נוכחות המיטוכונדריה. תאים אוקריוטיים עשויים להכיל בין אחד לכמה אלפי מיטוכונדריה, בהתאם לרמת צריכת האנרגיה של התא. כל מיטוכונדריון באורך של 1 עד 10 מיקרומטר ומעלה וקיים בתא כאברון שיכול להיות בצורת ביצה עד תולעת עד מסועף מורכב (איור). \(\PageIndex{1}\) מיטוכונדריה נובעות מחלוקת המיטוכונדריה הקיימת; הם עשויים להתמזג יחד; והם עשויים להיות מועברים בתוך התא על ידי אינטראקציות עם השלד. עם זאת, המיטוכונדריה אינה יכולה לשרוד מחוץ לתא. מכיוון שהאטמוספירה התחמצנה על ידי פוטוסינתזה, וככל שהתפתחו פרוקריוטים אירוביים מוצלחים, עדויות מצביעות על כך שתא אבות עם מידור ממברנה כלשהו בלע פרוקריוט אירובי חי חופשי, במיוחד אלפא-פרוטאובקטריום, ובכך נתן לתא המארח את היכולת להשתמש בחמצן לשחרר אנרגיה המאוחסנת בחומרים מזינים. אלפא-פרוטאובקטריה הם קבוצה גדולה של חיידקים הכוללת מינים סימביוטיים עם צמחים, אורגניזמים של מחלות שיכולים להדביק בני אדם באמצעות קרציות, ומינים רבים החיים חופשיים המשתמשים באור לאנרגיה. מספר קווי ראיות תומכים בכך שהמיטוכונדריה נגזרת מאירוע אנדוסימביוטי זה. רוב המיטוכונדריה מעוצבות כמו אלפא-פרוטאובקטריה ומוקפות בשתי ממברנות, מה שייווצר כאשר אורגניזם אחד הקשור לממברנה נבלע בוואקום על ידי אורגניזם אחר הקשור לממברנה. הממברנה הפנימית המיטוכונדריאלית נרחבת וכוללת פיתולים משמעותיים הנקראים cristae הדומים למשטח החיצוני המרקם של אלפא-פרוטאובקטריה. המטריצה והקרום הפנימי עשירים באנזימים הדרושים לנשימה אירובית.
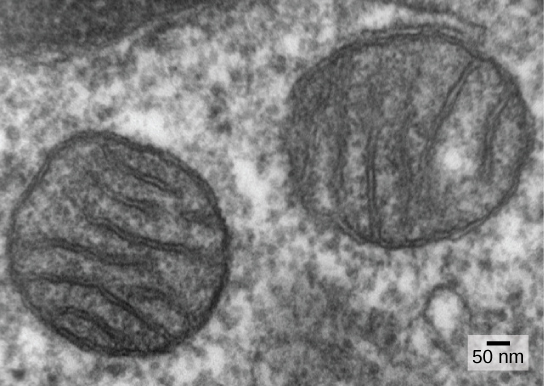
המיטוכונדריה מתחלקת באופן עצמאי בתהליך הדומה לביקוע בינארי בפרוקריוטים. באופן ספציפי, המיטוכונדריה אינן נוצרות מאפס (דה נובו) על ידי התא האוקריוטי; הם מתרבים בתוכו ומופצים עם הציטופלזמה כאשר תא מתחלק או שני תאים מתמזגים. לכן, למרות שאברונים אלה משולבים מאוד בתא האוקריוטי, הם עדיין מתרבים כאילו הם אורגניזמים עצמאיים בתוך התא. עם זאת, הרבייה שלהם מסונכרנת עם הפעילות והחלוקה של התא. למיטוכונדריה יש כרומוזום DNA מעגלי משלהם (בדרך כלל) שמתייצב על ידי התקשרות לקרום הפנימי ונושא גנים הדומים לגנים המתבטאים על ידי אלפא-פרוטאובקטריה. למיטוכונדריה יש גם ריבוזומים מיוחדים ו-RNA העברה הדומים לרכיבים אלה בפרוקריוטים. כל התכונות הללו תומכות בכך שהמיטוכונדריה היו פעם פרוקריוטים חיים חופשיים.
למיטוכונדריה המבצעות נשימה אירובית יש גנום משלהן, עם גנים דומים לאלה באלפא-פרוטאובקטריה. עם זאת, רבים מהגנים לחלבוני נשימה ממוקמים בגרעין. כאשר משווים גנים אלה לאלה של אורגניזמים אחרים, נראה שהם ממוצא אלפא-פרוטובקטריאלי. בנוסף, בחלק מהקבוצות האיקריוטיות, גנים כאלה נמצאים במיטוכונדריה, בעוד שבקבוצות אחרות הם נמצאים בגרעין. זה התפרש כראיה לכך שגנים הועברו מכרומוזום האנדוסימביונט לגנום המארח. אובדן גנים זה על ידי האנדוסימביונט הוא כנראה הסבר אחד מדוע המיטוכונדריה אינה יכולה לחיות ללא מארח.
חלק מהאוקריוטים החיים הם אנאירוביים ואינם יכולים לשרוד בנוכחות יותר מדי חמצן. נראה כי לחלקם חסרים אברונים שניתן לזהות אותם כמיטוכונדריה. בשנות ה-70 עד תחילת שנות ה-90, ביולוגים רבים הציעו שחלק מהאיקריוטים הללו היו צאצאים מאבות קדמונים ששושלתם התפצלה משושלת האיקריוטים המכילים מיטוכונדריה לפני שהתרחשה אנדוסימביוזה. עם זאת, ממצאים מאוחרים יותר מצביעים על כך שאברונים מופחתים נמצאים ברוב האאוקריוטים האנאירוביים, אם לא בכולם, וכי נראה כי כל האיקריוטים נושאים כמה גנים בגרעינים שלהם שמקורם במיטוכונדריה. בנוסף לדור האירובי של ATP, למיטוכונדריה יש מספר פונקציות מטבוליות אחרות. אחת הפונקציות הללו היא ליצור אשכולות של ברזל וגופרית שהם קופקטורים חשובים של אנזימים רבים. פונקציות כאלה קשורות לעתים קרובות לאברונים המופחתים שמקורם במיטוכונדריה של אוקריוטים אנאירוביים. לכן, רוב הביולוגים מקבלים שלאב הקדמון המשותף האחרון של האאוקריוטים היו מיטוכונדריה.
פלסטידים
כמה קבוצות של אוקריוטים הן פוטוסינתטיות. התאים שלהם מכילים, בנוסף לאברונים האוקריוטיים הסטנדרטיים, סוג אחר של אברון הנקרא פלסטיד. כאשר תאים כאלה מבצעים פוטוסינתזה, הפלסטידים שלהם עשירים בפיגמנט כלורופיל a ובמגוון פיגמנטים אחרים, הנקראים פיגמנטים נלווים, המעורבים בקצירת אנרגיה מאור. פלסטידים פוטוסינתטיים נקראים כלורופלסטים (איור). \(\PageIndex{2}\)
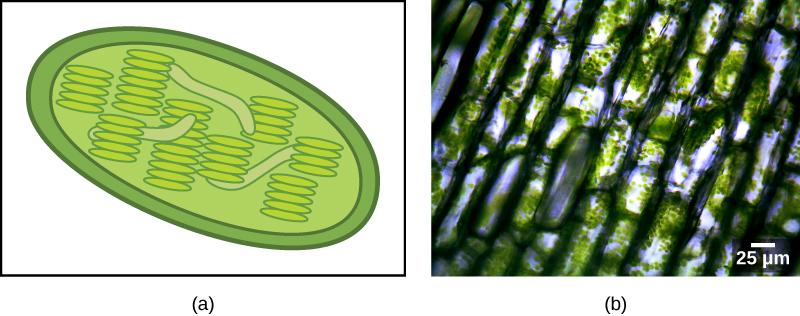
בדומה למיטוכונדריה, נראה כי לפלסטידים יש מקור אנדוסימביוטי. השערה זו ניצחה גם על ידי לין מרגוליס. פלסטידים נגזרים מציאנובקטריה שחיה בתוך תאים של אוקריוט קדמון, אירובי, הטרוטרופי. זה נקרא אנדוזימביוזה ראשונית, ופלסטידים ממוצא ראשוני מוקפים בשני ממברנות. העדות הטובה ביותר היא שזה קרה פעמיים בהיסטוריה של האוקריוטים. במקרה אחד, האב הקדמון המשותף של השושלה/קבוצת העל העיקרית Archaeplastida לקח על עצמו אנדוסימביונט ציאנובקטריאלי; בשני, האב הקדמון של הטקסון הריזארי האמבואידי הקטן, פאולינלה, לקח על עצמו אנדוסימביונט ציאנובקטריאלי אחר. כמעט כל האיקריוטים הפוטוסינתטיים הם צאצאים מהאירוע הראשון, ורק כמה מינים נגזרים מהאחר.
ציאנובקטריה הם קבוצה של חיידקים גראם שליליים עם כל המבנים המקובלים של הקבוצה. עם זאת, בניגוד לרוב הפרוקריוטים, יש להם שקיות נרחבות ופנימיות הקשורות לממברנה הנקראות תילקואידים. כלורופיל הוא מרכיב בממברנות אלו, וכך גם רבים מחלבוני תגובות האור של הפוטוסינתזה. לציאנובקטריה יש גם את דופן הפפטידוגליקן ושכבת הליפופוליסכריד הקשורים לחיידקים גראם שליליים.
לכלורופלסטים ממוצא ראשוני יש תילקואידים, כרומוזום DNA מעגלי וריבוזומים הדומים לאלה של ציאנובקטריה. כל כלורופלסט מוקף בשתי ממברנות. בקבוצה של Archaeplastida הנקראת glaucophytes ובפאולינלה קיימת שכבה פפטידוגליקנית דקה בין הממברנות הפלסטידיות החיצוניות והפנימיות. כל שאר הפלסטידים חסרים את הקיר הציאנובקטריאלי הרליקטואלי הזה. סבורים כי הממברנה החיצונית המקיפה את הפלסטיד נגזרת מהוואקום במארח, וחושבים שהממברנה הפנימית נגזרת מממברנת הפלזמה של הסימביונט.
יש גם, כמו במקרה של מיטוכונדריה, עדויות חזקות לכך שרבים מהגנים של האנדוסימביונט הועברו לגרעין. פלסטידים, כמו מיטוכונדריה, אינם יכולים לחיות באופן עצמאי מחוץ למארח. בנוסף, בדומה למיטוכונדריה, פלסטידים נגזרים מחלוקת פלסטידים אחרים ומעולם לא נבנים מאפס. חוקרים הציעו כי האירוע האנדוסימביוטי שהוביל לארכיפלסטידה התרחש לפני 1 עד 1.5 מיליארד שנה, לפחות 5 מאות מיליון שנים לאחר שהתיעוד המאובנים מצביע על כך שהיו איקריוטים.
לא כל הפלסטידים באאוקריוטים נגזרים ישירות מאנדוסימביוזה ראשונית. חלק מהקבוצות העיקריות של אצות הפכו לפוטוסינתטיות על ידי אנדוסימביוזה משנית, כלומר על ידי נטילת אצות ירוקות או אצות אדומות (שניהם מ- Archaeplastida) כאנדוסימביונטים (איור). \(\PageIndex{3}\) מחקרים מיקרוסקופיים וגנטיים רבים תמכו במסקנה זו. פלסטידים משניים מוקפים בשלושה ממברנות או יותר, ולחלק מהפלסטידים המשניים יש אפילו שרידים ברורים של גרעין האצה האנדוסימביוטית. אחרים לא "שמרו" שרידים. ישנם מקרים בהם אירועים אנדוסימביוטיים שלישוניים או מסדר גבוה יותר הם ההסברים הטובים ביותר לפלסטידים בחלק מהאיקריוטים.
חיבור אמנות
אילו עדויות יש לכך שהמיטוכונדריה שולבו בתא האוקריוטי הקדמוני לפני כלורופלסטים?
חיבור אבולוציה: אנדוסימביוזה משנית בכלורארכניופיטים
אנדוסימביוזה כרוכה בתא אחד הבולע תא אחר כדי לייצר, לאורך זמן, מערכת יחסים מתפתחת שבה אף תא לא יכול לשרוד לבד. הכלורופלסטים של אצות אדומות וירוקות, למשל, נגזרים מבליעה של ציאנובקטריה פוטוסינתטית על ידי פרוקריוט מוקדם.
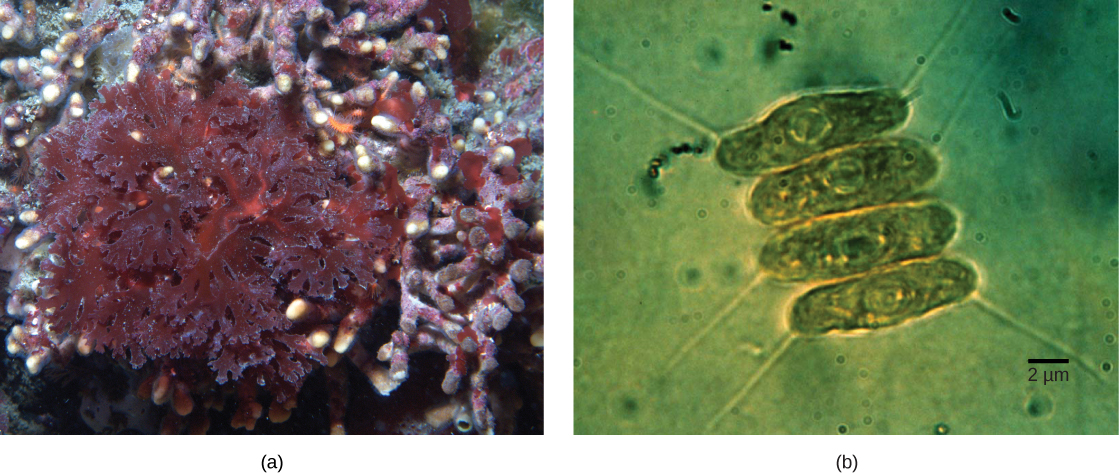
זה מוביל לשאלת האפשרות שתא המכיל אנדוסימביונט לעצמו ייבלע, וכתוצאה מכך אנדוסימביוזה משנית. עדויות מולקולריות ומורפולוגיות מצביעות על כך שהפרוטיסטים של כלורארכניופיטים נגזרים מאירוע אנדוסימביוטי משני. כלורארכניופיטים הם אצות נדירות שמקורן בים טרופי וחול שניתן לסווג לקבוצת העל הריזרית. כלורארכניופיטים מרחיבים גדילים ציטופלסמיים דקים, ומחברים את עצמם עם כלורארכניופיטים אחרים, ברשת ציטופלזמית. סבורים שמקורם של פרוטיסטים אלה כאשר אוקריוט בלע אצה ירוקה, שהאחרונה שלה כבר יצרה קשר אנדוסימביוטי עם ציאנובקטריה פוטוסינתטית (איור). \(\PageIndex{5}\)
מספר שורות ראיות תומכות בכך שכלורארכניופיטים התפתחו מאנדוסימביוזה משנית. הכלורופלסטים הכלולים בתוך האנדוסימביונטים של האצות הירוקות עדיין מסוגלים לפוטוסינתזה, מה שהופך את הכלורארכניופיטים לפוטוסינתטיים. האנדוסימביונט של האצות הירוקות מציג גם גרעין שריד מעוכב. למעשה, נראה כי כלורארכניופיטים הם תוצרים של אירוע אנדוסימביוטי משני אחרון מבחינה אבולוציונית. הפלסטידים של כלורארכניופיטים מוקפים בארבע ממברנות: השניים הראשונים תואמים את הממברנות הפנימיות והחיצוניות של הציאנובקטריה הפוטוסינתטית, השלישית תואמת את האצה הירוקה, והרביעית תואמת את הוואקוול שהקיף את האצה הירוקה כשהוא נבלע על ידי האב הקדמון של כלורארכניופיט. בשושלות אחרות שכללו אנדוסימביוזה משנית, ניתן לזהות רק שלוש ממברנות סביב פלסטידים. זה מתוקן כרגע כאובדן רציף של קרום במהלך האבולוציה.
תהליך האנדוסימביוזה המשנית אינו ייחודי לכלורארכניופיטים. למעשה, אנדוסימביוזה משנית של אצות ירוקות הובילה גם לפרוטיסטים אאוגלנידים, ואילו אנדוסימביוזה משנית של אצות אדומות הובילה לאבולוציה של דינופלגלטים, אפיקומפלקסנים וסטרמנופילים.
סיכום
העדות המאובנת העתיקה ביותר לאאוקריוטים היא בת כ-2 מיליארד שנה. מאובנים מבוגרים מזה נראים כולם פרוקריוטים. סביר להניח שהאוקריוטים של ימינו הם צאצאים מאב קדמון שהיה לו ארגון פרוקריוטי. לאב הקדמון המשותף האחרון של Eukarya של ימינו היו מספר מאפיינים, כולל תאים עם גרעינים שהתחלקו מיטוטית והכילו כרומוזומים ליניאריים שבהם ה-DNA היה קשור להיסטונים, מערכת שלד ואנדוממברנה, והיכולת ליצור ריסים/דגלים במהלך לפחות חלק ממחזור חייו. זה היה אירובי מכיוון שהיו לו מיטוכונדריה שהיו תוצאה של אלפא-פרוטאובקטריום אירובי שחי בתוך תא מארח. אם למארח זה היה גרעין בזמן הסימביוזה הראשונית עדיין לא ידוע. לאב הקדמון המשותף האחרון אולי היה דופן תא לפחות בחלק ממחזור חייו, אך יש צורך בנתונים נוספים כדי לאשר השערה זו. האיקריוטים של ימינו מגוונים מאוד בצורותיהם, בארגון, במחזורי החיים ובמספר התאים לאדם.
חיבורי אמנות
איור\(\PageIndex{4}\): אילו עדויות קיימות לכך שהמיטוכונדריה שולבו בתא האוקריוטי הקדמוני לפני כלורופלסטים?
- תשובה
-
לכל התאים האיקריוטים יש מיטוכונדריה, אך לא לכל התאים האיקריוטים יש כלורופלסטים.
רשימת מילים
- אנדוסימביוזה
- בליעת תא אחד בתוך תא אחר כך שהתא הנבלע שורד, ושני התאים מרוויחים; התהליך האחראי להתפתחות המיטוכונדריה והכלורופלסטים באיקריוטים
- תיאוריה אנדוסימביוטית
- תיאוריה הקובעת כי ייתכן שאאוקריוטים היו תוצר של תא אחד בולע אחר, אחד חי בתוך אחר, ומתפתח עם הזמן עד שהתאים הנפרדים כבר לא היו ניתנים לזיהוי ככאלה
- פלסטיד
- אחד מקבוצת אברונים קשורים בתאי צמחים המעורבים באחסון עמילנים, שומנים, חלבונים ופיגמנטים