
14.5: שכפול DNA באיקריוטים
- Page ID
- 205826

מיומנויות לפיתוח
- דון בדמיון ובהבדלים בין שכפול DNA באאוקריוטים ופרוקריוטים
- ציין את תפקיד הטלומראז בשכפול ה- DNA
הגנום האוקריוטי מורכב וגדול בהרבה מגנום פרוקריוטי. לגנום האנושי יש שלושה מיליארד זוגות בסיסים לכל קבוצה הפלואידית של כרומוזומים, ו-6 מיליארד זוגות בסיסים משוכפלים במהלך שלב S של מחזור התא. ישנם מקורות שכפול מרובים בכרומוזום האוקריוטי; לבני אדם יכולים להיות עד 100,000 מקורות שכפול. קצב השכפול הוא כ-100 נוקלאוטידים בשנייה, איטי בהרבה משכפול פרוקריוטי. בשמרים, שהוא אוקריוט, נמצאים על הכרומוזומים רצפים מיוחדים המכונים רצפים משכפלים אוטונומיים (ARS). אלה שקולים למקור השכפול ב- E. coli.
מספר פולימראזות ה-DNA באיקריוטים הוא הרבה יותר מפרוקריוטים: ידועים 14, מתוכם חמישה ידועים כבעלי תפקידים מרכזיים במהלך השכפול ונחקרו היטב. הם ידועים בשם פול α, פול β, פול γ, פול δ ופול ε.
שלבי השכפול החיוניים זהים לפרוקריוטים. לפני שהשכפול יכול להתחיל, ה-DNA צריך להיות זמין כתבנית. DNA אוקריוטי קשור לחלבונים בסיסיים המכונים היסטונים ליצירת מבנים הנקראים נוקלאוזומים. הכרומטין (הקומפלקס בין DNA לחלבונים) עשוי לעבור כמה שינויים כימיים, כך שה-DNA יוכל להחליק מהחלבונים או להיות נגיש לאנזימים של מכונות שכפול ה-DNA. במקור השכפול, נוצר קומפלקס טרום-שכפול עם חלבוני יוזם אחרים. לאחר מכן מגויסים חלבונים אחרים כדי להתחיל בתהליך השכפול (טבלה\(\PageIndex{1}\)).
הליקאז המשתמש באנרגיה מהידרוליזה של ATP פותח את סליל ה-DNA. מזלגות שכפול נוצרים בכל מקור שכפול כאשר ה-DNA מתפרק. פתיחת הסליל הכפול גורמת לפיתול יתר, או סליל-על, ב-DNA לפני מזלג השכפול. אלה נפתרים בפעולה של טופואיזומראזות. פריימרים נוצרים על ידי האנזים פרימאז, ובאמצעות הפריימר, DNA pol יכול להתחיל סינתזה. בעוד שהגדיל המוביל מסונתז ברציפות על ידי האנזים pol δ, הגדיל המפגר מסונתז על ידי pol ε. חלבון מהדק הזזה המכונה PCNA (אנטיגן גרעיני תאים מתרבים) מחזיק את פול ה- DNA במקומו כך שהוא לא יחליק מה- DNA. RNase H מסיר את פריימר ה- RNA, אשר מוחלף לאחר מכן בנוקלאוטידים של DNA. שברי Okazaki בגדיל הפיגור מחוברים יחד לאחר החלפת פריימרים RNA עם DNA. הפערים שנותרו אטומים על ידי ליגאז DNA, היוצר את הקשר הפוספודיסטר.
שכפול טלומרים
בניגוד לכרומוזומים פרוקריוטיים, הכרומוזומים האוקריוטיים הם ליניאריים. כפי שלמדת, האנזים DNA pol יכול להוסיף נוקלאוטידים רק בכיוון 5' עד 3'. בגדיל המוביל הסינתזה נמשכת עד שמגיעים לסוף הכרומוזום. על גדיל הפיגור, DNA מסונתז במתיחות קצרות, שכל אחת מהן יוזמת על ידי פריימר נפרד. כאשר מזלג השכפול מגיע לקצה הכרומוזום הליניארי, אין מקום ליצירת פריימר להעתקת שבר ה- DNA בסוף הכרומוזום. קצוות אלה נותרים אפוא ללא זיווג, ועם הזמן קצוות אלה עשויים להתקצר בהדרגה ככל שהתאים ממשיכים להתחלק.
קצות הכרומוזומים הליניאריים ידועים בשם טלומרים, שיש להם רצפים חוזרים שאינם מקודדים לגן מסוים. במובן מסוים, הטלומרים הללו מגנים על הגנים מפני מחיקה כאשר התאים ממשיכים להתחלק. בבני אדם, רצף של שישה זוגות בסיסים, TTAGGG, חוזר על עצמו 100 עד 1000 פעמים. גילוי האנזים טלומראז (איור\(\PageIndex{1}\)) עזר בהבנת האופן שבו נשמרים קצוות הכרומוזומים. האנזים טלומראז מכיל חלק קטליטי ותבנית RNA מובנית. הוא מתחבר לקצה הכרומוזום, ובסיסים משלימים לתבנית ה-RNA מתווספים בקצה 3' של גדיל ה-DNA. ברגע שקצה 3' של תבנית הגדיל המפגר מוארך מספיק, פולימראז DNA יכול להוסיף את הנוקלאוטידים המשלימים לקצוות הכרומוזומים. לפיכך, קצות הכרומוזומים משוכפלים.
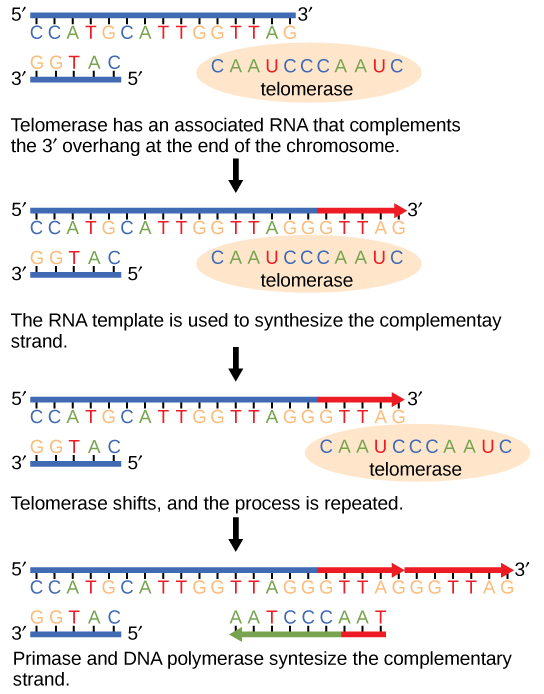
טלומראז פעיל בדרך כלל בתאי נבט ובתאי גזע בוגרים. הוא אינו פעיל בתאים סומטיים בוגרים. על גילוי הטלומראז ופעולתו, אליזבת בלקברן (איור\(\PageIndex{2}\)) קיבלה את פרס נובל לרפואה ופיזיולוגיה בשנת 2009.

טלומראז והזדקנות
תאים שעוברים חלוקת תאים ממשיכים לקצר את הטלומרים שלהם מכיוון שרוב התאים הסומטיים אינם מייצרים טלומראז. זה בעצם אומר שקיצור הטלומרים קשור להזדקנות. עם הופעת הרפואה המודרנית, טיפול מונע ואורח חיים בריא יותר, תוחלת החיים האנושית גדלה, ויש ביקוש הולך וגובר לאנשים להיראות צעירים יותר ואיכות חיים טובה יותר ככל שהם מתבגרים.
בשנת 2010, מדענים גילו כי טלומראז יכול להפוך כמה מצבים הקשורים לגיל בעכברים. זה עשוי להיות בעל פוטנציאל ברפואה רגנרטיבית. 1 עכברים חסרי טלומראז שימשו במחקרים אלה; לעכברים אלה יש ניוון רקמות, דלדול תאי גזע, כשל במערכת האיברים ותגובות פגיעה ברקמות לקויות. הפעלה מחדש של טלומראז בעכברים אלה גרמה להארכת הטלומרים, להפחתת נזק ל- DNA, ניוון עצבי הפוך ושיפרה את תפקוד האשכים, הטחול והמעיים. לפיכך, להפעלה מחדש של הטלומרים עשוי להיות פוטנציאל לטיפול במחלות הקשורות לגיל בבני אדם.
סרטן מאופיין בחלוקת תאים בלתי מבוקרת של תאים לא תקינים. התאים צוברים מוטציות, מתרבים ללא שליטה ויכולים לנדוד לאזורים שונים בגוף באמצעות תהליך הנקרא גרורות. מדענים הבחינו כי תאים סרטניים קיצרו במידה ניכרת את הטלומרים וכי הטלומראז פעיל בתאים אלה. מעניין שרק לאחר שהתקצרו הטלומרים בתאי הסרטן, הטלומראז הפך לפעיל. אם ניתן לעכב את פעולת הטלומראז בתאים אלה על ידי תרופות במהלך טיפול בסרטן, אזי ניתן היה לעצור את התאים הסרטניים מחלוקה נוספת.
| רכוש | פרוקריוטים | אוקריוטים |
|---|---|---|
| מקור השכפול | יחיד | מרובה |
| קצב השכפול | 1000 נוקלאוטידים/שניות | 50 עד 100 נוקלאוטידים/שניות |
| סוגי פולימראז DNA | 5 | 14 |
| טלומראז | לא נוכח | הווה |
| הסרת פריימר RNA | DNA פול I | RNase H |
| התארכות סטרנד | פול DNA III | פול δ, פול ε |
| מהדק הזזה | מהדק הזזה | PCNA |
סיכום
שכפול באאוקריוטים מתחיל במספר מקורות שכפול. המנגנון דומה למדי לפרוקריוטים. פריימר נדרש ליזום סינתזה, המורחבת לאחר מכן על ידי פולימראז DNA מכיוון שהיא מוסיפה נוקלאוטידים בזה אחר זה לשרשרת הגדלה. הגדיל המוביל מסונתז ברציפות, ואילו הגדיל המפגר מסונתז במתיחות קצרות הנקראות שברי Okazaki. הפריימרים של ה- RNA מוחלפים בנוקלאוטידים של DNA; ה- DNA נשאר גדיל רציף אחד על ידי קישור שברי ה- DNA עם ליגאז DNA. קצות הכרומוזומים מהווים בעיה מכיוון שפולימראז אינו מסוגל להאריך אותם ללא פריימר. טלומראז, אנזים בעל תבנית RNA מובנית, מרחיב את הקצוות על ידי העתקת תבנית ה- RNA והארכת קצה אחד של הכרומוזום. לאחר מכן DNA פולימראז יכול להאריך את ה-DNA באמצעות הפריימר. באופן זה מוגנים קצות הכרומוזומים.
הערות שוליים
- 1 Jaskelioff et al., "הפעלה מחדש של טלומראז הופכת את התנוונות הרקמות בעכברים חסרי טלומראז מיושנים", Nature 469 (2011): 102-7.
רשימת מילים
- טלומראז
- אנזים המכיל חלק קטליטי ותבנית RNA מובנית; הוא מתפקד לשמירה על טלומרים בקצות הכרומוזומים
- טלומרים
- DNA בקצה הכרומוזומים הליניאריים