
7.4: זרחון חמצוני
- Page ID
- 205696

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)מיומנויות לפיתוח
- תאר כיצד אלקטרונים נעים בשרשרת הובלת האלקטרונים ומה קורה לרמות האנרגיה שלהם
- הסבר כיצד שיפוע פרוטון (H +) נוצר ומתוחזק על ידי שרשרת הובלת האלקטרונים
קראת זה עתה על שני מסלולים בקטבוליזם של גלוקוז - גליקוליזה ומחזור חומצת לימון - היוצרים ATP. רוב ה- ATP שנוצר במהלך הקטבוליזם האירובי של גלוקוז, לעומת זאת, אינו נוצר ישירות ממסלולים אלה. במקום זאת, הוא נגזר מתהליך שמתחיל בהעברת אלקטרונים דרך סדרה של מובילי אלקטרונים שעוברים תגובות חיזור. זה גורם ליוני מימן להצטבר בתוך חלל המטריצה. לכן נוצר שיפוע ריכוז שבו יוני מימן מתפזרים מחוץ לחלל המטריצה על ידי מעבר דרך סינתאז ATP. הזרם של יוני מימן מניע את הפעולה הקטליטית של סינתאז ATP, אשר מזרחן ADP, ומייצר ATP.
שרשרת הובלת אלקטרונים
שרשרת הובלת האלקטרונים (איור\(\PageIndex{1}\)) היא המרכיב האחרון בנשימה אירובית והיא החלק היחיד בחילוף החומרים של הגלוקוז המשתמש בחמצן אטמוספרי. חמצן מתפזר ברציפות לצמחים; אצל בעלי חיים הוא נכנס לגוף דרך מערכת הנשימה. הובלת אלקטרונים היא סדרה של תגובות חיזור הדומות למירוץ ממסר או חטיבת דלי בכך שהאלקטרונים מועברים במהירות מרכיב אחד למשנהו, לנקודת הקצה של השרשרת שבה האלקטרונים מפחיתים חמצן מולקולרי, ומייצרים מים. ישנם ארבעה קומפלקסים המורכבים מחלבונים, המסומנים I עד IV באיור\(\PageIndex{1}\), והצבירה של ארבעת המתחמים הללו, יחד עם נשאי אלקטרונים ניידים נלווים, נקראת שרשרת הובלת האלקטרונים. שרשרת הובלת האלקטרונים קיימת במספר עותקים בקרום המיטוכונדריאלי הפנימי של האאוקריוטים ובממברנת הפלזמה של הפרוקריוטים.
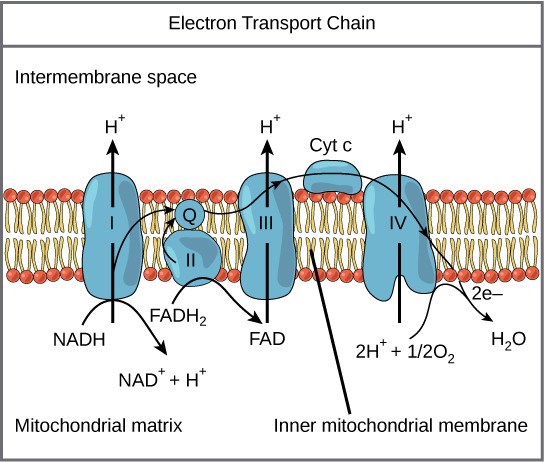
קומפלקס I
כדי להתחיל, שני אלקטרונים מועברים למתחם הראשון על סיפון NADH. קומפלקס זה, שכותרתו I, מורכב ממונונוקלאוטיד פלבין (FMN) וחלבון המכיל ברזל-גופרית (Fe-S). FMN, שמקורו בוויטמין B 2, הנקרא גם ריבופלבין, הוא אחת מכמה קבוצות תותבות או גורמים משותפים בשרשרת הובלת האלקטרונים. קבוצה תותבת היא מולקולה שאינה חלבונית הנדרשת לפעילות של חלבון. קבוצות תותבות הן מולקולות אורגניות או אנאורגניות, שאינן פפטידיות הקשורות לחלבון המקלות על תפקודו; קבוצות תותבות כוללות אנזימים משותפים, שהם קבוצות האנזימים התותבות. האנזים במתחם I הוא NADH dehydrogenase והוא חלבון גדול מאוד, המכיל 45 שרשראות חומצות אמינו. קומפלקס I יכול לשאוב ארבעה יוני מימן על פני הממברנה מהמטריצה לחלל הבין-ממברני, ובדרך זו מתבסס ומתוחזק שיפוע יוני המימן בין שני התאים המופרדים על ידי הממברנה המיטוכונדריאלית הפנימית.
Q ומתחם II
מתחם II מקבל ישירות FADH 2, שאינו עובר במתחם I. המתחם המחבר את המתחם הראשון והשני לשלישי הוא ubiquinone (Q). מולקולת Q מסיסה בשומנים ונעה בחופשיות דרך הליבה ההידרופובית של הממברנה. ברגע שהוא מצטמצם, (QH 2), ubiquinone מעביר את האלקטרונים שלו למתחם הבא בשרשרת הובלת האלקטרונים. Q מקבל את האלקטרונים שמקורם ב- NADH ממתחם I ואת האלקטרונים שמקורם ב- FADH 2 ממתחם II, כולל succinate dehydrogenase. אנזים זה ו- FADH 2 יוצרים קומפלקס קטן המספק אלקטרונים ישירות לשרשרת הובלת האלקטרונים, ועוקף את המתחם הראשון. מכיוון שאלקטרונים אלה עוקפים ובכך אינם ממריצים את משאבת הפרוטון בקומפלקס הראשון, פחות מולקולות ATP מיוצרות מהאלקטרונים FADH 2. מספר מולקולות ה- ATP המתקבלות בסופו של דבר עומד ביחס ישר למספר הפרוטונים הנשאבים על פני הממברנה המיטוכונדריאלית הפנימית.
מתחם III
המתחם השלישי מורכב מציטוכרום b, חלבון Fe-S נוסף, מרכז ריסקה (מרכז 2Fe-2S) וחלבוני ציטוכרום c; קומפלקס זה נקרא גם ציטוכרום אוקסידורדוקטאז. לחלבוני ציטוכרום יש קבוצה תותבת של heme. מולקולת ההמה דומה להאם בהמוגלובין, אך היא נושאת אלקטרונים, לא חמצן. כתוצאה מכך, יון הברזל שבליבתו מצטמצם ומתחמצן כאשר הוא עובר את האלקטרונים, משתנה בין מצבי חמצון שונים: Fe ++ (מופחת) ו- Fe +++ (מחומצן). למולקולות ההמה בציטוכרומים יש מאפיינים מעט שונים בשל השפעות החלבונים השונים הקושרים אותם, ומעניקים מאפיינים מעט שונים לכל קומפלקס. קומפלקס III שואב פרוטונים דרך הממברנה ומעביר את האלקטרונים שלו לציטוכרום c להובלה לקומפלקס הרביעי של חלבונים ואנזימים (ציטוכרום c הוא מקבל האלקטרונים מ- Q; עם זאת, בעוד ש- Q נושא זוגות אלקטרונים, ציטוכרום c יכול לקבל רק אחד בכל פעם).
קומפלקס IV
הקומפלקס הרביעי מורכב מחלבוני ציטוכרום c, a ו-a 3. קומפלקס זה מכיל שתי קבוצות heme (אחת בכל אחת משני הציטוכרומים, a ו- a 3) ושלושה יוני נחושת (זוג Cu A ואחד Cu B בציטוכרום a 3). הציטוכרומים מחזיקים מולקולת חמצן בחוזקה רבה בין יוני הברזל והנחושת עד שהחמצן מצטמצם לחלוטין. החמצן המופחת קולט אז שני יוני מימן מהמדיום שמסביב ליצירת מים (H 2 O). הוצאת יוני המימן מהמערכת תורמת לשיפוע היונים המשמש בתהליך הכימיוזמוזה.
כימיוזמוזה
בכימיוזמוזה, האנרגיה החופשית מסדרת תגובות החיזור שתוארו זה עתה משמשת לשאיבת יוני מימן (פרוטונים) על פני הממברנה. ההתפלגות הלא אחידה של יוני H+ על פני הממברנה קובעת הן ריכוז והן שיפועים חשמליים (לפיכך, שיפוע אלקטרוכימי), בשל המטען החיובי של יוני המימן והצטברותם בצד אחד של הממברנה.
אם הממברנה הייתה פתוחה לדיפוזיה על ידי יוני המימן, היונים היו נוטים להתפזר חזרה אל המטריצה, מונעים על ידי שיפוע האלקטרוכימי שלהם. נזכיר כי יונים רבים אינם יכולים להתפזר דרך האזורים הלא קוטביים של ממברנות הפוספוליפידים ללא עזרת תעלות יונים. באופן דומה, יוני מימן בחלל המטריצה יכולים לעבור רק דרך הממברנה המיטוכונדריאלית הפנימית דרך חלבון ממברנה אינטגרלי הנקרא ATP synthase (איור\(\PageIndex{2}\)). חלבון מורכב זה פועל כמחולל זעיר, המופנה בכוח יוני המימן המתפזרים דרכו, במורד השיפוע האלקטרוכימי שלהם. הפיכת חלקים ממכונה מולקולרית זו מקלה על הוספת פוספט ל- ADP, ויוצר ATP, תוך שימוש באנרגיה הפוטנציאלית של שיפוע יון המימן.
חיבור אמנות
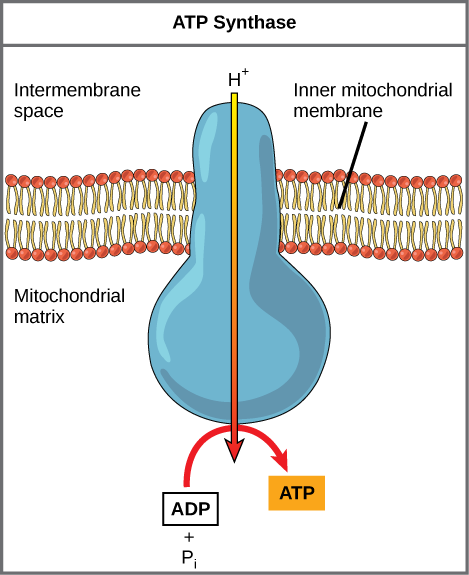
דיניטרופנול (DNP) הוא מפרק שגורם לקרום המיטוכונדריאלי הפנימי לדלוף לפרוטונים. הוא שימש עד 1938 כתרופה לירידה במשקל. איזו השפעה היית מצפה שתהיה ל-DNP על השינוי ב-pH על פני הממברנה המיטוכונדריאלית הפנימית? מדוע לדעתך זו יכולה להיות תרופה יעילה לירידה במשקל?
כימיוזמוזה (איור\(\PageIndex{3}\)) משמשת ליצירת 90 אחוז מה-ATP שנוצר במהלך קטבוליזם גלוקוז אירובי; זוהי גם השיטה המשמשת בתגובות האור של הפוטוסינתזה כדי לרתום את אנרגיית אור השמש בתהליך הפוטופוספורילציה. נזכיר כי ייצור ATP באמצעות תהליך הכימיוזמוזה במיטוכונדריה נקרא זרחון חמצוני. התוצאה הכוללת של תגובות אלו היא ייצור ATP מהאנרגיה של האלקטרונים שהוסרו מאטומי מימן. אטומים אלה היו במקור חלק ממולקולת גלוקוז. בסוף המסלול, האלקטרונים משמשים להפחתת מולקולת חמצן ליוני חמצן. האלקטרונים הנוספים על החמצן מושכים יוני מימן (פרוטונים) מהמדיום שמסביב, ונוצרים מים.
חיבור אמנות
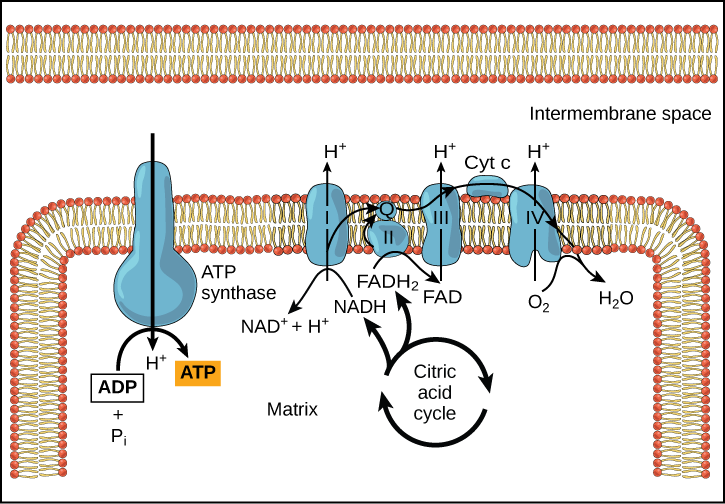
ציאניד מעכב ציטוכרום c אוקסידאז, מרכיב בשרשרת הובלת האלקטרונים. אם מתרחשת הרעלת ציאניד, האם היית מצפה שה-pH של החלל הבין-ממברני יגדל או יירד? איזו השפעה תהיה לציאניד על סינתזת ATP?
תשואת ATP
מספר מולקולות ה- ATP הנוצרות מקטבוליזם של גלוקוז משתנה. לדוגמה, מספר יוני המימן שמתחמי שרשרת הובלת האלקטרונים יכולים לשאוב דרך הממברנה משתנה בין המינים. מקור נוסף לשונות נובע ממעבורת האלקטרונים על פני ממברנות המיטוכונדריה. (ה-NADH שנוצר מגליקוליזה לא יכול להיכנס בקלות למיטוכונדריה.) לפיכך, אלקטרונים נאספים בחלק הפנימי של המיטוכונדריה על ידי NAD + או FAD +. כפי שלמדת קודם, מולקולות FAD+אלה יכולות להעביר פחות יונים; כתוצאה מכך, פחות מולקולות ATP נוצרות כאשר FAD+פועל כנשא. NAD + משמש כמעביר האלקטרונים בכבד ו- FAD+פועל במוח.
גורם נוסף המשפיע על התשואה של מולקולות ATP הנוצרות מגלוקוז הוא העובדה שתרכובות ביניים במסלולים אלה משמשות למטרות אחרות. קטבוליזם של גלוקוז מתחבר למסלולים הבונים או מפרקים את כל התרכובות הביוכימיות האחרות בתאים, והתוצאה קצת יותר מבולגנת מהמצבים האידיאליים שתוארו עד כה. לדוגמה, סוכרים שאינם גלוקוז מוזנים למסלול הגליקוליטי להפקת אנרגיה. יתר על כן, הסוכרים בעלי חמישה פחמנים היוצרים חומצות גרעין עשויים מחומרי ביניים בגליקוליזה. ניתן לייצר חומצות אמינו לא חיוניות מסוימות מתוצרי ביניים הן של הגליקוליזה והן של מחזור חומצת הלימון. ליפידים, כמו כולסטרול וטריגליצרידים, עשויים גם הם מחומרי ביניים במסלולים אלה, וגם חומצות אמינו וגם טריגליצרידים מתפרקים לאנרגיה דרך מסלולים אלה. בסך הכל, במערכות חיות, מסלולים אלה של קטבוליזם של גלוקוז מפיקים כ -34 אחוז מהאנרגיה הכלולה בגלוקוז.
סיכום
שרשרת הובלת האלקטרונים היא החלק של הנשימה האירובית המשתמשת בחמצן חופשי כמקבל האלקטרונים הסופי של האלקטרונים שהוסרו מתרכובות הביניים בקטבוליזם של גלוקוז. שרשרת הובלת האלקטרונים מורכבת מארבעה קומפלקסים גדולים מרובי חלבונים המוטבעים בקרום המיטוכונדריאלי הפנימי ושני נשאי אלקטרונים קטנים הניתנים לפיזור המעבירים אלקטרונים ביניהם. האלקטרונים מועברים דרך סדרה של תגובות חיזור, כאשר כמות קטנה של אנרגיה חופשית משמשת בשלוש נקודות להובלת יוני מימן על פני קרום. תהליך זה תורם לשיפוע המשמש בכימיוזמוזה. האלקטרונים העוברים בשרשרת הובלת האלקטרונים מאבדים בהדרגה אנרגיה, אלקטרונים בעלי אנרגיה גבוהה שנתרמו לשרשרת על ידי NADH או FADH 2 משלימים את השרשרת, שכן אלקטרונים בעלי אנרגיה נמוכה מפחיתים מולקולות חמצן ויוצרים מים. רמת האנרגיה החופשית של האלקטרונים יורדת מכ-60 קק"ל/מול ב-NADH או 45 קק"ל/מול ב-FADH 2 לכ-0 קק"ל/מול במים. התוצרים הסופיים של שרשרת הובלת האלקטרונים הם מים ו-ATP. ניתן להפנות מספר תרכובות ביניים של מחזור חומצת הלימון לאנבוליזם של מולקולות ביוכימיות אחרות, כגון חומצות אמינו לא חיוניות, סוכרים ושומנים. אותן מולקולות יכולות לשמש כמקורות אנרגיה למסלולי הגלוקוז.
חיבורי אמנות
איור\(\PageIndex{2}\): דיניטרופנול (DNP) הוא מפרק שגורם לקרום המיטוכונדריאלי הפנימי לדלוף לפרוטונים. הוא שימש עד 1938 כתרופה לירידה במשקל. איזו השפעה היית מצפה שתהיה ל-DNP על השינוי ב-pH על פני הממברנה המיטוכונדריאלית הפנימית? מדוע לדעתך זו יכולה להיות תרופה יעילה לירידה במשקל?
- תשובה
-
לאחר הרעלת DNP, שרשרת הובלת האלקטרונים כבר לא יכולה ליצור שיפוע פרוטונים, וסינתאז ATP כבר לא יכול ליצור ATP. DNP היא תרופת דיאטה יעילה מכיוון שהיא מנתקת את סינתזת ה- ATP; במילים אחרות, לאחר נטילתו, אדם משיג פחות אנרגיה מהמזון שהוא אוכל. מעניין, אחת מתופעות הלוואי הגרועות ביותר של התרופה היא היפרתרמיה, או התחממות יתר של הגוף. מכיוון שלא ניתן ליצור ATP, האנרגיה מהובלת אלקטרונים הולכת לאיבוד כחום.
איור\(\PageIndex{3}\): ציאניד מעכב ציטוכרום c אוקסידאז, מרכיב בשרשרת הובלת האלקטרונים. אם מתרחשת הרעלת ציאניד, האם היית מצפה שה-pH של החלל הבין-ממברני יגדל או יירד? איזו השפעה תהיה לציאניד על סינתזת ATP?
- תשובה
-
לאחר הרעלת ציאניד, שרשרת הובלת האלקטרונים אינה יכולה עוד לשאוב אלקטרונים לחלל הבין-ממברני. ה- pH של החלל הבין -ממברני יעלה, שיפוע ה- pH יירד וסינתזת ATP תיפסק.
רשימת מילים
- סינתאז ATP
- (כמו כן, F1F0 ATP synthase) קומפלקס חלבון מוטבע ממברנה המוסיף פוספט ל-ADP עם אנרגיה מפרוטונים שמתפזרים דרכו
- קבוצה תותבת
- (גם, קופקטור תותב) מולקולה הקשורה לחלבון שמקל על תפקוד החלבון
- יוביקינון
- טרנספורטר אלקטרונים מסיס בשרשרת הובלת האלקטרונים המחבר את המתחם הראשון או השני לשלישי