
10.1: מקרה מבחן - נספחים אקטודרמליים
- Page ID
- 207621

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)תוספות אקטודרמליות הן איברים הקשורים לעור כמו נוצות ושיער. בדרך כלל אנו מכנים רק הרחבות עור חוליות "תוספות חוץ רחמיות" למרות שלקבוצות בעלי חיים אחרות יש גם איברי עור (כמו בלוטות). בחלק זה נתמקד רק בתוספות החוליות עם דגש מיוחד על בלוטות החלב. פרטים על כך וכן סיכום של התפתחות איברי עור אחרים ניתן למצוא כאן: מנגנונים של אורגנוגנזה חוץ רחמית.
לתוספות (או איברים) אקטודרמליים יש דפוס התפתחותי משותף ונחשבים שיש להם גם איבר אבולוציוני משותף, כאשר אופציה משותפת של אלמנטים מסוגי תוספת שונים מתרחשת לעתים קרובות. נספחים אלה ברורים תחילה כעיבוי האפידרמיס ליצירת "פלאקוד". לאחר מכן התפשטות תאית של האפידרמיס ועיבוי של תאים מזנכימליים הבסיסיים של הדרמיס ליצירת ניצן עם בסיס מזנכימלי עבה. הניצן הנגזר חוץ רחמי יכול כעת לעבור אחד ממסלולים מרובים, אותו הוא בוחר בהתבסס על אותות מהמזודרם - המזנכיים של הדרמיס וכן המזודרם הסומיטי (איור 1). אנו מסווגים בערך את הנספחים הללו לשתי קבוצות עיקריות: תוספות הפה, כולל שיניים ובלוטות הרוק, ותוספות העור, כולל נוצות, קשקשים, שיער, בלוטות חלב, בלוטות זיעה ובלוטות שמן.
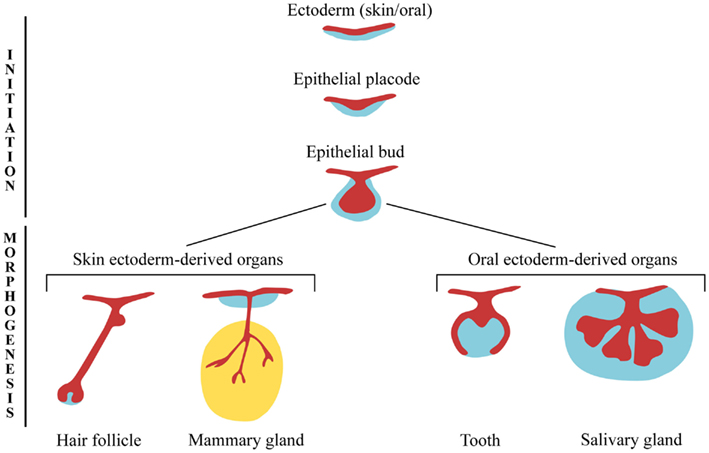
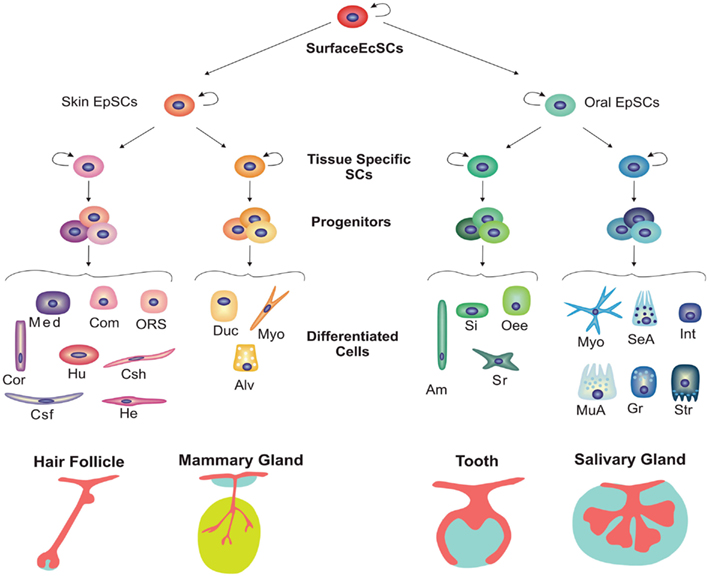
בשלב מוקדם, מדענים הבחינו במשהו מעניין - ברגע שהיה במקום פלאקוד חוץ רחמי, ניתן היה לגרום לו ליצור אחת מרקמות רבות. לדוגמה, מזנכימה של ניצן החלב יכולה לעורר היווצרות בלוטות חלב באפיתל הגבי 2. ניסוי מעניין אחד הראה שמזנכימה דנטלית של עכברים יכולה אפילו לגרום לשיניים באפיתל הפה של האפרוח, למרות העובדה שציפורים בדרך כלל לא מגדלות שיניים 3. כפי שנדון קודם לכן, התפתחות תוספת חוץ רחמית עוקבת אחר המסלול של כללי לספציפי (איור 2) כאשר הפלקודים המוקדמים מוגדרים פחות מניצנים מאוחרים יותר.
אבולוציה וגיוון של נספחי עור טטרפודים
לדגים יש תוספות עור מדהימות ומגוונות משלהם, כולל איברי "הקו הרוחבי" המזהים שדות חשמליים, לחץ ושינויי זרימת מים 4. עם זאת, כאן נתמקד בתוספות האקטודרמליות של הטטרפודים תוך התמקדות בבלוטות החלב של ממליה. ההשערה היא כי לטטרפוד הקדמוני היו בלוטות ריר בעורו, בדומה לאלה של דו-חיים חיים. בלוטות ריר אלה שומרות על לחות העור ומונעות את התייבשות החלקים הפנימיים של החיה. בנוסף, בלוטות ריר ובלוטות עור אחרות משמשות את הדו-חיים המודרניים הדוהרים כדי למנוע מהביצים נטולות קליפת הביצים שלהם להתייבש במערכות אקולוגיות יבשתיות וייתכן שמילאו תפקיד דומה אצל טטרפודים מוקדמים (איור 3).

לפני כ -350 מיליון שנה, קבוצה אחת של טטרפודים, אמניוטה, פיתחה סוג חדש של חלבון עור שמנע את התייבשות עורם: קרטין. לכל מי השפיר (כולל יונקים, ציפורים, לטאות וכו ') יש אלפא-קרטין ולסורופסידים (ציפורים וזוחלים אחרים) יש בנוסף בטא-קרטין, בו הם משתמשים לבניית נוצות וקשקשים. בניגוד ליונקים המטילים ביצים, לסורופסידים יש קליפות ביצים מסוידות המפחיתות את איבוד המים. ליונקים המטילים ביצים (המונוטרמים כמו פלטיפוס), יש ביצים מקולפות קלף שיכולות להתייבש במהירות. זה נחשב כתנאי האבות לקבוצה המכילה את היונקים, הסינפסידים והפרשות העור הבלוטות מההורה הדוהר (כמו הפרשות ריר בסלמנדרות) נחשבים כסייעו במניעת התייבשות ביציות בסינפסידים אבותיים כולל יונקים 5.
היונק הקדמון ירש אפוא עור בלוטות שהיה רב תכליתי: סביר להניח שהוא הפריש שמן, זיעה והפרשות מדוללות אחרות (אולי סוג של זיעה) כדי לשמור על התייבשות הביציות שלו. אנו יכולים לקבל כמה רמזים כיצד בלוטות העור הללו התגוונו לתוספות אקטודרמליות מרובות על ידי השוואת שלושת החלקים החיים של המאמלים, המונוטרמים המטילים ביצים, חיות הכיס הטריות והשליות התריות (הידועות גם בשם האתריות). ליונקים ארבעה סוגים עיקריים של בלוטות עור, אלה בלוטות האוזן המייצרות שעווה (מהן נתעלם לעת עתה, מבלוטות החלב המייצרות שמן, בלוטות הזיעה ובלוטות החלב. לשלושת הבלוטות האחרונות יש קשרים אנטומיים מיוחדים זה עם זה. זקיק שיער טיפוסי קשור לבלוטת חלב ויכול להיות קשור גם לבלוטת זיעה אפוקרינית. בלוטות החלב במינים מסוימים קשורות לזקיק שיער כמו גם לבלוטת חלב. זה הוביל להשערה שבלוטות החלב נגזרות לפחות בחלקן מבלוטות זיעה אפוקריניות (איור 4).
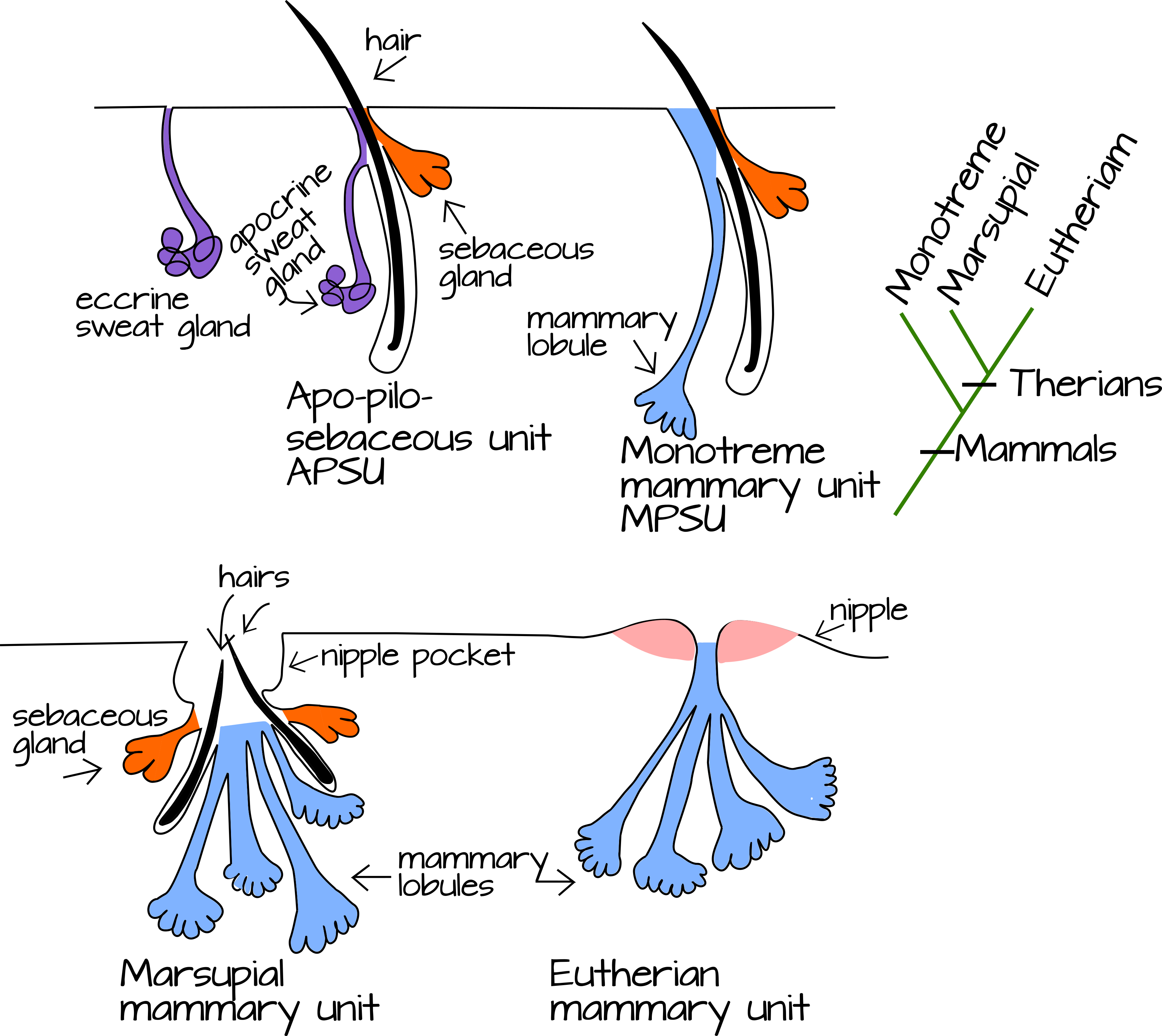 |
איור 4: השוואה בין בלוטות עור יונקים. ה- APSU (יחידת apo-pilosebaceous) הוא סוג נפוץ של בלוטת עור הנמצאת בבית השחי ובמפשעה של בני אדם. הוא מורכב מבלוטת חלב, זקיק שיער ובלוטת זיעה אפוקרינית. בלוטות אפוקריניות מפרישות על ידי צביטה של חלק מתא הפרשה ושחרורו לצינור. זה שונה מאוד מבלוטת זיעה טיפוסית (אקרינית) המפרישה על ידי אקסוציטוזיס ונמצאת בכל עורם של בני אדם. ה- MPSU (יחידה ממולובולרית-פילוסבאסית) נחשב ל- APSU נגזר עם תאי הפרשה ממולובולריים אפוקריניים המחליפים את תאי בלוטת הזיעה. בלוטת חלב מכילה MPSU אחד או יותר ופטמה (ברוב המקרים, אם כי לחלק מהיונקים כמו מונוטרמים חסרים פטמות). |
סקירה כללית של התפתחות בלוטת החלב
למרות המגוון בצורותיהן הסופיות, בלוטות החלב מתפתחות לאורך מסלולים דומים במידה רבה על פני יונקים. כפי שמוצג באיור 1, הם נגרמים על ידי מזנכיים בסיסיים ואותות אחרים ליצירת שני קווי חלב מקבילים, אשר ככל הנראה נגרמים על ידי הפרשת FGF10 המושרה על ידי GLI3 על ידי קבוצת משנה של המזודרם הסומיטי, המניע את הביטוי של WNT10b באקטודרם מעל וציון גורל תאי החלב 6. מונוטרמים שומרים על קווי החלב ככתמים של תאים המפרישים חלב. דיבור צולב בין האפידרמיס לדרמיס (mesenchymal) יוצר ניצן חלב, הצומח כלפי מטה לתוך המזנכיים ליצירת נבט חלב. אצל תריאנים, ניצני החלב והנבטים מקבלים איתות BMP4 מהמזנכימה הבסיסית כדי להמשיך ולמנוע מהם לקחת את גורל ברירת המחדל של זקיק השיער 6. בחלקים אחרים של העור, Noggin ו- Shh מעכבים את BMP4 ומאפשרים את גורל השיער המוגדר כברירת מחדל 6. ניצני החלב האפיתליים מבטאים pTHRP (פפטיד הקשור להורמון פארתירואיד), המפעיל את הקולטן שלו במזנכיים כדי לגרום לעיבוי והתמיינות של הנבט הראשוני החלב הגדל. לפיכך יש דיבור צולב בין האפידרמיס לדרמיס (mesenchyme) כדי ליצור את ניצן החלב ולגרום לצמיחה לתוך הנבט הראשוני. בחלק מיונקי השליה כמו עכברים הנבט הראשוני גדל עמוק לתוך רקמת השומן ההיפודרמלית, שם הוא מתחיל להסתעף לרקמת בלוטות מכתשית. אצל יונקים אחרים הוא מסתעף בצורה רדודה יותר לנבטים משניים, שבעצמם מסתעפים לרקמת בלוטות מכתשית 8.
לאן נעלם השיער?
אם בלוטות החלב התפתחו מזקיקי השיער והבלוטות הקשורות אליהם, מה קרה לשיער? בעוד ליונקים שליה רבים יש שיער ארולרי (איור 5), לרובם אין שיער הקשור לחלק הפטמה המפריש חלב. בנוסף, אף יונקי שליה שנבדקו לא מפתחים זקיק שיער במהלך או לפני היווצרות נבט חלב. אני אחזור לנושא יונקי השליה בעוד רגע, אבל קודם אני רוצה להקדיש קצת זמן להסתכל על בני הדודים השעירים שלנו.
מונוטרמים, אותם יונקים המסתעפים באופן בסיסי, אכן שומרים על קשר שיער עם בלוטות החלב (איור 4). אופטדל מציע שזהו מקרה היונקים הקדמונים וכי לשערות החלב, ולא לפטמה, יש שימוש כפול: הן לא רק מנפות חלב מבלוטות העור לפיו של תינוק, אלא הן גם מנפות הפרשות לחות לביצים מועדות להתייבשות. האבולוציה של התפתחות עוברית פנימית ולידה חיה אצל היונקים הטריים שינו את הלחץ האבולוציוני על הבלוטות המייצרות חלב. הם איבדו את הלחץ הסלקטיבי לשמור על לחות ביציות ובמקום זאת נבחרו אך ורק להזנת תינוקות. זה אולי הניע את צינורות החלב להתעבות ולזרום לתוך פטמה אחת לכל בלוטת החלב, ולא כתם שיער מפוזר (איור 6) 5,8.
עדויות מהתפתחות בלוטות החלב אצל יונקים תריאנים נותנות תמיכה מסוימת להשערה זו. אצל חיות כיס, התפתחות מוקדמת של בלוטת החלב מתרחשת כגרסה מרוכזת פיזית של התפתחות מונוטרמה. במונוטרמים, נבט ראשוני של החלב מתפתח לזקיק שיער, לבלוטת חלב ולתאי הפרשת החלב והצינור (MPSU הרב-רכיבי). בחיות כיס, נבט ראשוני יחיד יוצר MPSU מרובים, כל אחד עם זקיק השיער שלו, בלוטת החלב ותאי החלב. עם זאת, זקיקי השיער בסופו של דבר מתנוונים ומתים, ומשאירים אחריהם כיס גדול שאליו חלב וחלב יכולים להתרוקן (איור 4) 8. יונקים שליה (eutherians), כמונו, משתנים יותר. האותרים יכולים לייצר נבט ראשוני אחד או רבים לכל נורת חלב (וכאמור לעיל) עשויים ליצור או לחסר נבטים משניים. למינים ללא נבטים משניים אין זקיקי שיער וחלב הקשורים לבלוטות החלב הגדלות, אך למינים עם נבטים משניים יש. זה כולל בני אדם וסוסים. זקיקי השיער של MPSU ובלוטות החלב שלנו מתנוונים ומתים במהלך האמבריוגנזה ולכן נשארנו עם MPSU העשוי מרכיב יחיד: alveoli החלב וצינורות 8.
על ידי השוואת התפתחות בלוטות החלב של קבוצות שונות אלה, Oftedal ו- Dhouailly הציעו תרחיש להתפתחות בלוטות החלב על ידי אופציה משותפת והטרוכרוניה. בקצרה, בלוטות החלב משתמשות במנגנוני התפתחות קיימים של פלקודה בעור כדי להתחיל להיווצר, בפרט בדפוסים הקיימים המשמשים את בלוטות האפו-פילוסבאביס, ואז עוברות לתוכנית של מורפוגנזה מסתעפת (מנגנון דפוס התפתחותי המצוי בכליות, בריאות ובבלוטות הרוק בטטרפודים אך גם בנוצות ציפורים והתפתחות קנה הנשימה של חרקים). זה יוצר את העץ הצינורי של בלוטת החלב כחלק האפוקריני של היחידה האפו-פילוסבוסית, במקום בלוטת זיעה 8. מקור הפטמה שמעליה עדיין לא ברור אך נגרם התפתחותית על ידי בלוטת החלב המתפתחת.
הגנטיקה של התפתחות בלוטות החלב: אפשרות משותפת וחידוש
מוקדם יותר, דנתי בקצרה בבחירת הגורל שהעור עושה בין שיער לבלוטת החלב - הפעלת איתות BMP4 בתאים מזנכימליים באמצעות איתות FGF10 מחלק מהמזודרם הסומיטי מכבה את תוכנית השיער המוגדרת כברירת מחדל ומפעילה את תוכנית החלב. אינטראקציות FGF/Wnt/BMP משמשות ליצירת עור אחר באקטודרם מעל משמש בדפוסי תוספות עור אחרות ועשוי להיות קלטת כללית לדפוס אקטודרמי 9. בדרך זו, בלוטות החלב placode והיווצרות ניצנים מונעות ככל הנראה על ידי איתות דומה לתוספות אקטודרמליות אחרות עם כמה שינויים. ברגע שהשלייה/ניצן החלב מתחיל לצמוח, הוא מבטא גן איבר אפידרמיס נפוץ: pTHRP. ל- PthRP יש תפקיד אבות יותר בחולייתנים ביצירת עצם אנדוכונדרית והתפרצות שיניים. מכיוון ששיניים הן מבנה אפיתל חוץ רחמי, סביר שתפקידו של PthRP בהתפתחות בלוטות החלב נבע מהתפתחות השיניים. עדויות לאופציה משותפת של גנים בשיניים מגיעות ממפתח אחר של משפחת גנים בהתפתחות השיניים - חלבון קושר הסידן ODAM. ODAM שוכפל במהלך התפתחות הטטרפודים ליצירת הקזאינים, חלבוני הובלת הסידן בחלב 10.
FGF10 ו- Wnt10b אינם השחקנים היחידים במפרט החלב. מספר גנים של Hox מעורבים במפרט איברים חוץ רחמיים, כאשר Hoxc8 יוזם התפתחות פלאקוד החלב של העכבר ו- Hoxb3, b6, b9, d9, d10 ו- d8, בין היתר, באו לידי ביטוי במהלך התפתחות מאוחרת יותר, כולל במהלך התפתחות החלב לאחר הלידה 11,12. מתוכם, Hoxd9 ו- d8 מתבטאים לרוב בנורות החלב של העכבר. שני הגנים הללו מעורבים גם בהתפתחות הגפיים והשיער בעכברים.
בעוד Hoxd9 ו-d8 ממוקמים זה ליד זה בכרומוזום ומווסתים במידה רבה על ידי שני אלמנטים ענקיים cis-רגולטוריים Hoxd-t-dom ו-Hoxd-C-dom, הם אינם מציגים דפוסי ביטוי זהים לחלוטין, מה שמצביע על כך שיש רגולטורים ספציפיים הפועלים באופן דיפרנציאלי על היזמים האישיים שלהם. לאחרונה, מדענים במעבדת בקארי בז'נבה ניתחו את האלמנטים הרגולטוריים המניעים את ביטוי Hox d8 ו- d9 ומצאו כיצד הם הצליחו לעבור מביטוי בגפה ובשיער לביטוי בנורת החלב. בעוד שביטוי נורת החלב של Hoxd8 היה תלוי באזור משפר מקומי של 13kb, ביטוי נורת החלב Hoxd9 היה תלוי באזורים רגולטוריים מרוחקים של cis ב-T-DOM 12.
האלמנט הרגולטורי של נורת החלב Hoxd9 cis מסקרן במיוחד מכיוון שנראה שהוא מסתמך בחלקו על האלמנטים הרגולטוריים הקיימים של cis המניעים את ביטוי הגפיים של Hoxd9. ביטוי הגפיים מונע על ידי אלמנטים רגולטוריים cis בתוך T-DOM ובמהלך התפתחות הגפיים, אזור ענק של 1500 קילו-בייט של הכרומוזום מתקפל ליצירת קשר עם Hoxd9. משפר ניצני החלב עבור Hoxd9 נמצא גם באזור זה של 1500 kb, וסבורים כי משפר זה מנצל את המשיכה בין משפר ניצן הגפיים לאשכול Hox כדי לקבל גישה ל- Hoxd9 במהלך התפתחות נורת החלב (איור 7) 12. ביחד, זה מתחיל לצייר תמונה של האבולוציה של פלקודים ונורות החלב - הם משתמשים בגורם שעתוק קיים ובגנים דפוסים בשילובים חדשים. לפחות אחד מהגנים המעורבים מתבטא ברקמת החלב המתפתחת על ידי שיתוף ושינוי אלמנטים משפרים קיימים.
לאחר בניית בלוטות החלב העובריות, הן עדיין צריכות להפריש חלב. בעוד שהדמיון בין בלוטות המייצרות חלב לבלוטות זיעה אפוקריניות ברור ברמות אנטומיות והתפתחותיות, התוכן של שני החומרים המיוצרים שונה באופן מהותי. מוצר חלב אחד, הקזאינים, עשוי מחלבון שיניים ODAM מוטציה (ראה לעיל). החלבון העיקרי החיוני בעיבוד טיפות השומן המצויות בחלב, XOR, הוא בדרך כלל חלבון אנטי-מיקרוביאלי אך ממלא תפקיד שני בביוסינתזה של חלב. זהו מקרה ברור של הטרוטופיה (ביטוי של גן בתחום חדש). ליזוזים לקח את שני הנתיבים - הוא הוצא ממערכת החיסון כחלבון אנטי-מיקרוביאלי בבלוטות החלב, והוא גם עבר שכפול גנים ליצירת חלבון מי גבינה המשמש ללקטוז 13. למרות שהרכב החלב מורכב ומשתנה ממין למין (אפילו בתוך מינים ככל שהתינוק מתבגר), אנו יכולים להתחקות אחר ההיסטוריה האבולוציונית של רכיבים אלה ולראות כיצד האבולוציה הייתה יכולה לפעול לייצור הפרשה זו.
הפניות
- "קביעת גורל תאי גזע במהלך התפתחות והתחדשות של איברים חוץ רחמיים." (2012) גבולות בפיזיולוגיה, לוסיה ג'ימנז-רוג'ו, זורייד גרנצ'י, דניאל גראף ותימיוס א 'מיציאדיס, https://doi.org/10.3389/fphys.2012.00107.
- "ביטוי פנוטיפי של החלב המושרה בתאי אפידרמיס על ידי מזנכיים של החלב העוברי" (1995) ג'רלד ר 'קונה, פיטר יאנג, קונסטנטין טי כריסטוב, ר סי גוזמן, ס 'בננדי, פרנק ג'יי טלמנטס, גודמונדור ת'ורדרסון, PMID: 7572029.
- "התפתחות שיניים בעוברי אפרוחים לאחר השתלות ציצה עצבית של עכברים", (2003), PNAS, תימיוס א 'מיטסיאדיס, איבונניק צ'ראוד, פול שארפ וג'וזיאן פונטן-פרוס, https://doi.org/10.1073/pnas.1137104100.
- "הקו הרוחבי יכול לתווך ריאוטקסיס בדגים" (1997), טֶבַע, ג'ון סי מונטגומרי, סינדי פ בייקר ואלכסנדר ג'י קרטון, https://doi.org/10.1038/40135.
- "מקור ההנקה כמקור מים לביצים מופגזות קלף" (2002) כתב העת לביולוגיה וניאופלזיה של בלוטות החלב, אולב טי אופטדל, https://doi.org/10.1023/A:1022848632125. .
- "תרחיש חדש למקור האבולוציוני של קשקשי שיער, נוצות ועופות", (2009) כתב העת לאנטומיה, דניאלה דהוילי, דוי: 10.1111/j.1469-7580.2008.01041.x.
- "חלבון הקשור להורמון פארתירואיד שומר על גורל אפיתל החלב ומעורר התמיינות עור הפטמה במהלך התפתחות השד העוברי." (2001) פיתוח, ג'יי פולי, פ 'דן, ג'יי הונג, ג'יי קוסגרוב, ב' דרייר, ד 'רים, מ' דונבר, וו פילבריק, ג'יי וויסולמרסקי, PMID: 11171335.
- "Evo-devo של בלוטת החלב", (2013), כתב העת לביולוגיה של בלוטות החלב וניאופלזיה, אולב ט 'אופטדל ודניאלה דואילי, דוי: 10.1007/s10911-013-9290-8.
- "איתות FGF נדרש לתחילת פיתוח פלוקוד נוצות" (2004) פיתוח, מרקוס מנדלר, אנט נוביזר, דוי: 10.1242/dev.01203.
- "האבולוציה של גנים של קזאין חלב מגנים של שיניים לפני מקורם של יונקים" (2012), קזוהיקו קוואסאקי, אן-גאל לאפונט, ז'אן איב אדון, ביולוגיה מולקולרית ואבולוציה, https://doi.org/10.1093/molbev/msr020.
- "HOXC8 יוזם תוכנית חלב חוץ רחמית על ידי ויסות ביטוי Fgf10 ו- Tbx3, ואיתות Wnt/β-catenin" (2015) פיתוח, לארה ס 'קרול, מריו ר קפצ'י, דוי: 10.1242/dev.128298.
- "שליטה בתעתיק גנים של הוקסד בניצן החלב על ידי חטיפת נוף רגולטורי קיים" (2016), PNAS, רובן שפ, אנמריה נקסולאה, אדי רודריגז-קרבלו, איזבל גרריירו, גיום אנדריי, ת'י האן נגוין הוינה, וירג'יני מרסט, יוזף זקני, דניס דובוין לה, ולאונרדו בקארי, https://doi.org/10.1073/pnas.1617141113.
- "אבולוציה של בלוטת החלב ממערכת החיסון המולדת?" (2006) מסות ביולוגיות, קלאודיה וורבך מריו ר 'קאפצ'י יוסף מ' פנינגר, doi.org/10.1002/bies.20423.
תמונה ממוזערת: תמונת תקריב של נוצות טווס, שהיא דוגמה אחת לתוספות אקטודרמליות. (נחלת הכלל).