4.2: איתות והגבלת גורל: תקשורת תא-תא - בתוך התא
- Page ID
- 207723

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)תקשורת תא-תאים: מסלולי העברת אותות
מסלולי העברת אותות מצמידים אותות חיצוניים לשינויים בביטוי הגנים בתוך תא. כלומר, אות חיצוני מעורר מפל של שינויים ביוכימיים בתא וכתוצאה מכך שעתוק גבוה או נמוך יותר של קבוצת גנים. פחות מתריסר מסלולי העברת אותות עיקריים מסדירים בדרך כלל את התפתחות בעלי החיים 5,6. כאן, נתמקד בארבעה כאלה הנקראים על שם הקולטן או הליגנד שלהם: Notch, קיפוד (Hh), TGF-b (נקרא גם Dpp או BMP) ו-Wnt (איור 4). מסלולי העברת אותות אלה פועלים כמתגים עבור רשתות רגולטוריות של גנים (נדון להלן) אותם הם מפעילים באמצעות מנגנונים שונים. שני גורמים עיקריים משפיעים על סוגי תהליכי הדפוס והמפרט שמסלול העברת אותות מסוים מתאים להם.
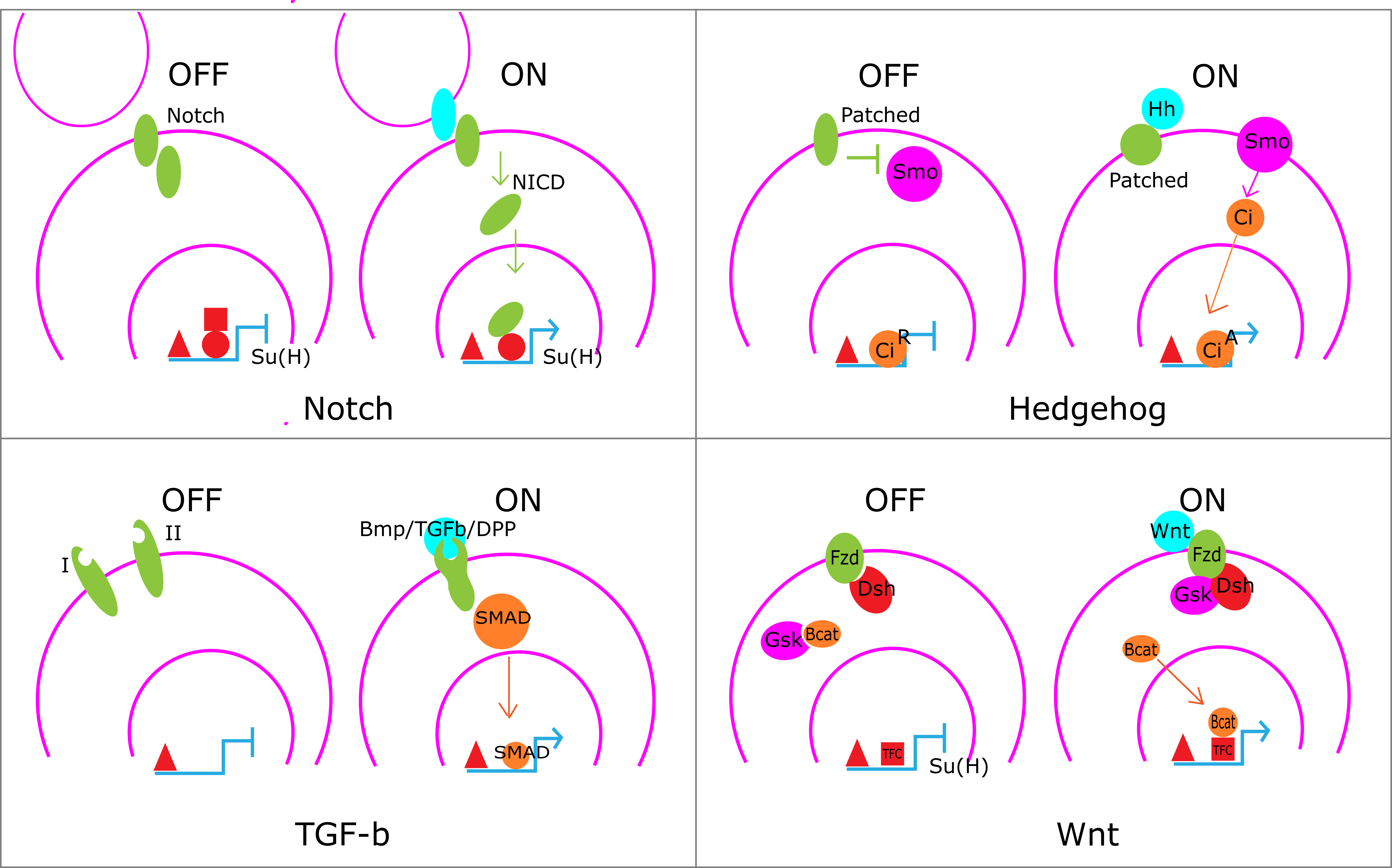
ראשית, כיצד מועבר האות? האם הוא משתמש בליגנד לטווח ארוך (איתות פאראקריני), האם הוא דורש מגע בין תא לתא (איתות juxtacrine), או שהליגנד פועל על אותו תא ממנו הופרש (איתות אוטוקריני)? מסלולי פראקרין ארוכי טווח, כמו TGF-b ו-Wnt, מצוינים ביצירת שיפועים על פני רקמה גדולה ומשמשים לעתים קרובות כמורפוגנים מוקדמים המעוצבים צירי גוף או סוגי תאים מרובים על פני רקמה. מסלולי פראקרין לטווח קצר, כמו Hh, פועלים לעתים קרובות כמורפוגנים בקנה מידה קטן יותר - כוונון עדין של דפוסי אזוריות. גורמים אחרים משנים את טווח האיתות של ליגנדים אלה, למשל כמות הליגנד המופרש, נטרול על ידי חלבוני מטריצה חוץ-תאית, מספר התאים המגיבים והאם תאים מגיבים מבטאים מעכבים או קואקטיבטורים של גני המטרה 6. אותות Juxtacrine, כמו מסלול Notch, כרוכים בקשר של שני קולטנים הקשורים לממברנה. במקרה של Notch, זהו בדרך כלל ליגנד מסוג Delta/Serrate/Lag-12 (DSL) וחלבון חריץ גליקוזילציה באורך מלא. מסלול ה- Notch משמש לעתים קרובות בהחלטות גורל תא הפעלה/כיבוי, המפורסמות בעיכוב רוחבי שבו אות "פועל" בתא אחד מפעיל אות "כבוי" בכל התאים שמסביב.
הגורם החשוב השני המסדיר את התועלת של מסלול העברת אותות הוא הרגולציה שלו. ניתן לווסת את רוב המסלולים במידה מסוימת על ידי גני המטרה שלהם במורד הזרם באמצעות משוב שלילי או חיובי. במשוב שלילי גני המטרה במורד הזרם מכבים בסופו של דבר את המסלול. לדוגמה, מסלול איתות יכול להגביר את השעתוק של מעכב מסלול. במשוב חיובי הם מטרות במורד הזרם שומרות על המסלול. לדוגמה, מסלול איתות יכול להגביר את התעתיק של הקולטן שלו. לפיכך, משוב שלילי טוב לקידום אות חד פעמי חולף, בעוד שמשוב חיובי ממיר אות חולף להחלטה קבועה של גורל התא. מסלולים יכולים גם להיות מווסתים זה על ידי זה וניתן לשנות את התפוקה שלהם על ידי גורמי שעתוק מקומיים, שיכולים להיות שונים בין סוגי התאים. נושא זה מכוסה בהרחבה יותר בשלושה הרגלים של מסלולי איתות יעילים ביותר. מאת בורולו ופוסקוני.
בתוך התא: רשתות רגולטוריות של גנים
בתחילת שנות האלפיים, אריק דוידסון ואיזבל פיטר כתבו סדרת מאמרים המציעים מסגרת פילוסופית להבנת רשתות רגולטוריות גנטיות (GRN). בשנת 2011 הם כתבו מאמר מכריע על האבולוציה של רשתות אלה. הם הגדירו שלושה סוגים בסיסיים של אינטראקציה גנטית ליבה המשמשים לציון סוגי תאים:
- גרעינים: אלה אינטראקציות בלתי גמישות מבחינה אבולוציונית המציינות חלק בגוף.
- סוללות: אלה מעורבים בהתמיינות תאים או רקמות והם גמישים יותר מבחינה אבולוציונית מאשר גרעינים.
- תוספים: אלה הם תת-מעגלים קטנים שמתרגלים בהקשרים התפתחותיים רבים ושונים 7.
כאם לחובבי לגו, אני משתמשת בערכות לגו כאנלוגיה כדי להבין זאת. גרעין יהיה חלקים מיוחדים שהולכים יחד, כמו זוג גלגלים וציר. הם מציינים פונקציה מסוימת ואינם גמישים באינטראקציה שלהם - יש להצמיד גלגל אחד לכל צד של הציר כדי להפוך אותו לשימושי. "סוללה" גמישה יותר אך עדיין משמשת בדרך כלל למערך מטרות דומה. דוגמה לכך תהיה חבילת סוללות מילולית בערכת לגו - היא משמשת להפעלת תנועה אך גמישה בכך שהיא יכולה להניע סוגים רבים של קונסטרוקציות. תוסף דומה לבנים סטנדרטיות, ניתן להשתמש בהן לבניית כל חפץ קובואידי שדמיונך מציע על מנת ליצור קונסטרוקציות מהודרות, עם זאת, אתה זקוק גם לגרעינים וסוללות.
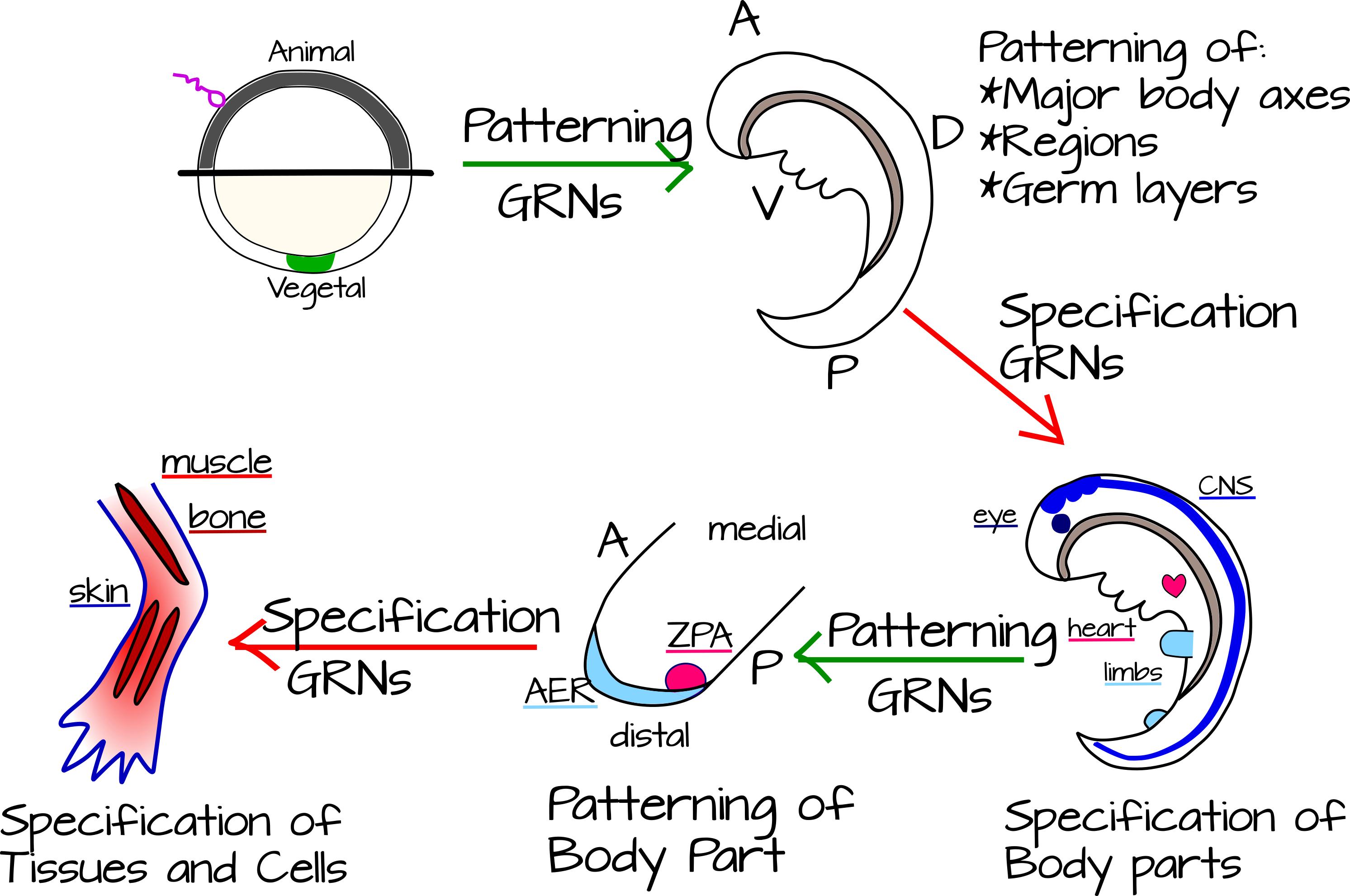
כיצד אנו מקשרים סוגים אלה של אינטראקציות לעובר מתפתח? פיטר ודוידסון תופסים את התפתחות בעלי החיים כשלבים מתחלפים של דפוסים ומפרט 7. דפוס הוא חלוקת גוף או קבוצה של רקמות לחלקים קטנים יותר. מפרט הוא בחירת גורל לתא או לרקמה שעוצבה בדוגמת. בתפיסה זו, צירי הגוף ושכבות הנבט מוגדרים לראשונה באמצעות רשתות גנטיות של דפוסי גוף. לאחר מכן, כל חלק של הגוף מוגדר: neuroectoderm, מעיים, קשתות הלוע (ב chordates), מערכת הדם, וכו 'ואז, סיבוב נוסף של דפוסים של יחידות אלה שצוין מתרחשת - כל חלק הגוף מקבל כעת דפוס לאורך הצירים שלו. לדוגמה, קצות האצבעות לעומת מרפק או הקדמי לעומת האחורי של הלב (איור 5). 8