
4.1: מבוא: התמונה הגדולה
- Page ID
- 207724

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)כבר התחלנו לבחון גנטיקה התפתחותית וקיבלנו טעימה מהמורכבות של אינטראקציות גנטיות. לדוגמה, הגן Drosophila Bicoid מועתק ל- mRNA בתאי האם ונשאב לתוך העובר. שם הביטוי שלו מוסדר על ידי אותות לוקליזציה על ה- mRNA, כמו גם השפלה ולוקליזציה של החלבון. חלבון ביקואידי עצמו פועל הן כמווסת של שעתוק והן כתרגום של גנים במורד הזרם. האפקט הסופי הוא עובר עם גנים שונים המתבטאים לאורך ציר A/P.
כעת ניקח צעד אחורה ונסתכל על תמונה רחבה יותר של גנטיקה התפתחותית, מנקודת המבט של מסלולי איתות. מסלול איתות מאפשר לתאים לתקשר עם הסביבה החיצונית שלהם. בביולוגיה התפתחותית, בדרך כלל מדובר באינטראקציות בין תאים לתאים. אינטראקציות מסוג זה חשובות להפליא מכיוון שכל תא צריך לעקוב אחר מסלול ההתפתחות שלו בתיאום עם כל התאים סביבו. לדוגמה, דמיינו ניצן גפיים של יונקים גדל. כל תא בניצן הגפה צריך לדעת אם הוא נמצא בצד האגודל או הזרת של הגפה וכמה הוא קרוב לגוף. לא רק זה, הוא גם צריך לדעת כמה זמן התקדמה ההתפתחות. תארו לעצמכם אם תא אוסטאוגני התחיל ליצור מטריצת עצם בשלב מוקדם בהתפתחות הגפיים. זה לא יעבור מספיק חלוקות תאים כדי ליצור את המספר הנכון של אוסטאובלסטים, מה שמוביל למעט מדי עצם ובעל חיים עם איבר פגום.
ערכת הכלים הגנטית
(קובץ PDF מבוא ניתן למצוא כאן: גילוי מחדש של ביולוגיה www.learner.org/קורסים/ביולוגיה/pdf/7_gendev.pdf)
אחד הממצאים העיקריים של Evo-Devo הוא שאורגניזמים חולקים קבוצה משותפת של גנים לבניית גופם המגוון מאוד. בפרט, בעלי חיים חולקים חבילה של גנים של "תוכנית גוף" המבצעים פונקציות דומות בהתפתחות. נראה כי ערבוב והתאמה של דפוסי ביטוי גנים שונים על פני רקמות וסוגי תאים הם המנגנון העיקרי ליצירת הגופים הייחודיים שאנו רואים בכל קבוצת בעלי חיים. נראה כי מאפיינים אבולוציוניים חדשים (כמו קוצים, נוצות, לבבות תאיים, מחושים וכו ') מסתמכים על שימוש חוזר בגנים של ערכת כלים בזמנים ובמקומות חדשים בהתפתחות (ראה טפסים אינסופיים לפרטים נוספים). להלן, נתחיל להסתכל על הגנטיקה שמאחורי ערכת הכלים הגנטית הזו - כיצד הגנים הללו מתקשרים זה עם זה?
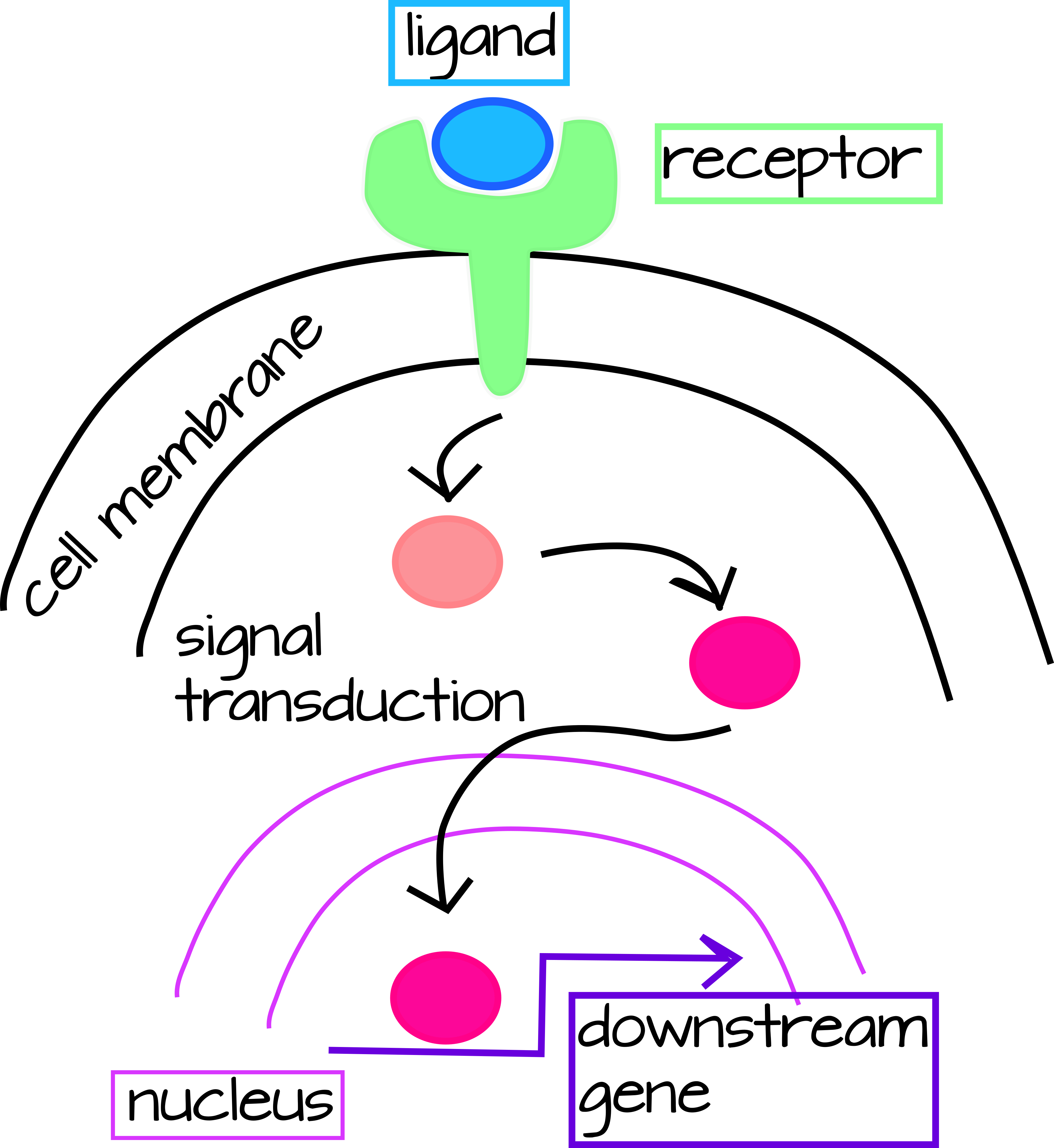
ב- Evo/Devo, ישנן שתי דרכים עיקריות בהן אנו יכולים לחשוב על אינטראקציות גנטיות, אך מכיוון שהן קשורות זו בזו, נשקול את שתיהן בו זמנית. האחד הוא מסלולי התמרת אותות, בעלי אות חיצוני (בביולוגיה התפתחותית זהו בדרך כלל חלבון מופרש) המתקבל בדרך כלל על ידי קולטן קרום תא שרק תאים מסוימים מבטאים. כאשר הליגנד (החלבון המופרש) והקולטן נקשרים, הדבר מעורר מפל של שינויים בחלבון המובילים להפעלה תעתיק של גנים במורד הזרם (איור 1).
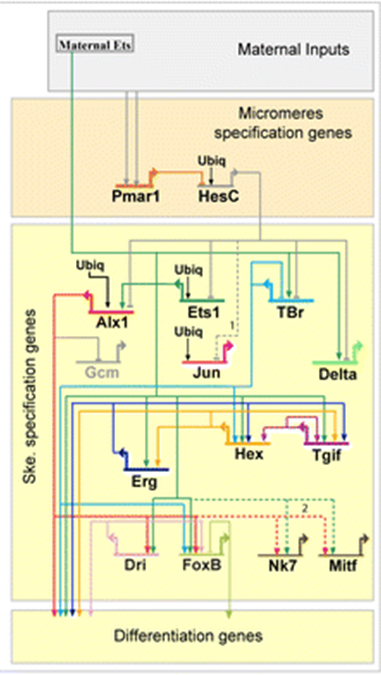
הדרך השנייה שבה Evo/Devo חושב על איתות היא רשתות רגולטוריות גנטיות (GRN). זה כולל את מסלול העברת האותות והשפעותיו במורד הזרם על ביטוי גנים. אם נגביר את הביטוי של גן אחד, על אילו גנים אחרים הוא משפיע? GRNs מיוצגים לעתים קרובות כדיאגרמות חיווט עם חצים וסורגים המציגים את ההשפעה של חלבונים על הביטוי של גנים במורד הזרם (איור 2).
 |
.png) |
| איור 1: מסלול העברת אותות גנרי. ליגנד חוץ תאי נקשר לקולטן הקשור לממברנה המפעיל הפעלה של מפל העברת אותות בציטופלזמה. מפל זה עשוי להיות פשוט (כמו במקרה של מסלול Notch למטה) או מורכב מאוד הדורש מספר שלבים וסוגי קלט. השלב האחרון של מפל התמרה הוא טרנסלוקציה של גורם שעתוק ציטופלזמי לגרעין שם הוא יפעיל או יעכב את התעתיק של גן במורד הזרם. ניתן להוריד קובץ svg הניתן לעריכה של דמות זו בכתובת https://scholarlycommons.pacific.edu/open-images/18/ | איור 2: דיאגרמת חיווט חלקית של רשת רגולטורית גנטית למפרט המזודרם היוצר שלד בקיפוד הים, Strongylocentrotus purpuratus. כל גן מיוצג על ידי קו אופקי המחובר לחץ באותו צבע ייחודי. הקו האופקי מציין את cis-אזור הרגולציה של הגן והחץ מציין שעתוק של אזור קידוד ה- mRNA. קווים הנובעים מאזור קידוד ה- mRNA מציינים כיצד תוצר החלבון של הגן מווסת גנים אחרים במורד הזרם. תשומות לאזורים הרגולטוריים cis על ידי מוצרי החלבון של גנים אחרים עשויות להיות מופעלות (חץ) או מעכבות (בר). "Ubiq" הוא מפעילים לא ידועים המתבטאים בכל התאים. איור מאת דילוס ואח '1 פורסם תחת רישיון CC BY 4.0. הדמות נחתכה מהמקור. |
התמונה הגדולה: גורלות התא מוגבלים בהדרגה ככל שהפיתוח מתקדם
במהלך ההתפתחות יש לציין את כל האיברים, הרקמות וסוגי התאים. אצל בעלי חיים זה מושג על ידי הגבלת גורל חוזרת ונשנית 2. לדוגמה, העובר הטוטיפוטנטי עובר מספר סבבי חלוקה ליצירת בלסטולה (כדור תאים). חלק מהתאים הללו יקבלו גורל מזודרמלי. בשלב זה הם יכולים להפוך לכל סוג של תא מזודרמלי. בבעלי חיים מסוימים בוצעו ניסויים כדי להראות שתא מזודרמלי גנרי יכול להיגרם על ידי שכניו להתפתח למגוון עצום של סוגי תאים מזודרמליים 3. עם זאת, עם הזמן, תאים מזודרמליים אלה מתחלקים, עוברים שינויי צורה, נודדים ומתמיינים. כשהם מקבלים אותות מסביבתם (בעיקר התאים סביבם) הם מתמיינים לסוגי תאים ספציפיים יותר. לדוגמה, איור 3 מציג תא מזודרמלי המתמיין לתאי שריר, עצם ודם. ככל שהזמן מתקדם, תאי אבות שרירים שונים (מיוגניים) יתמיינו לסוגים שונים של שרירים, למשל חלקים ומפוספסים. כל אחת מהחלטות גורל התא הללו תלויה בדפוס ביטוי הגנים בתא, התלוי בהיסטוריה של התא ובשכניו.

אנו מכנים סוג זה של בידול פרוגרסיבי "היררכי" עם סוגי תאים גנריים יותר (כמו מזודרם, איור 3) בראש ההיררכיה וסוגי תאים ספציפיים יותר (כמו נויטרופילים) בתחתית 4. ככל שהפיתוח מתקדם, אנו עוברים במורד ההיררכיה. פיתוח מתחיל מפרט ציר, צירים אלה נקראים במהרה לאזורים - למשל ראש, גזע/בטן וזנב. בתוך אזורים שונים אלה רקמות מוגדרות, תחילה ברמת שכבות הנבט ומאוחר יותר ברמות ספציפיות יותר. לדוגמה, אקטודרם מתפצל לנוירואקטודרם ואקטודרם אפידרמיס בהתאם למיקום הרקמה. לאחר מכן, סוגי תאים בתוך הרקמות מפורטים. לדוגמה, neuroectoderm יכול להפוך לנוירונים או לתאי גליה. שני סוגי התאים הללו מתמיינים מאוחר יותר לסוגים הספציפיים הרבים של נוירונים וגליה. בשלב זה, התאים נבדלים פחות או יותר סופנית - גורלם הבוגר מוגדר והם אינם יכולים לחזור לצורות פלוריפוטנטיות. עם זאת, רוב התאים עדיין פתוחים לסביבה ויכולים להגיב לאותות סביבתיים גם ברגע שהם מובחנים סופנית. אנו מתייחסים לזה כאל פלסטיות סלולרית.
ה-GRN באיור 2 משקף את ההיררכיה הזו, כאשר גנים הפועלים מוקדם בחלק העליון מציינים רקמות כלליות יותר וגנים הפועלים מאוחר יותר בתחתית מציינים סוגי תאים ספציפיים. עם זאת, חשוב לציין כי הגנים הפועלים ברמות היררכיות שונות אלה נמצאים בשימוש חוזר ברמות אחרות של ההיררכיה וברקמות מתפתחות אחרות. למעשה, חלקי הליבה של GRNs ומסלולי העברת אותות נמצאים בשימוש חוזר לאורך כל הפיתוח ויש להם תפוקות שונות בהתאם להקשר שלהם.