
8.8: תמלול הפוך
- Page ID
- 207769

בדוגמה המרכזית, DNA מקודד ל- mRNA, המקודד לחלבון. יוצא מן הכלל הידוע לדוגמה המרכזית מוצג על ידי רטרו-וירוסים. לנגיפים המקודדים ל-RNA הללו יש שלב במחזור החיים שלהם שבו ה-RNA הגנומי שלהם מומר בחזרה ל-DNA על ידי אנזים מקודד ויראלי המכונה טרנסקריפטאז הפוך. היכולת להמיר RNA ל- DNA היא שיטה רצויה במעבדה מסיבות רבות. לדוגמה, המרת RNA מעניינים ל- cDNA משמשת ב- RT-PCR כמו גם ביישומים אחרים כמו ניתוח מיקרו-מערכים.
תהליך
ראשית, יוצרים אוליגונוקלאוטיד DNA שישמש כפריימר לשימוש טרנסקריפטאז הפוך על RNA מטרה. הפריימר חייב, כמובן, להיות משלים לקטע (ליד קצה 3') של ה-RNA כדי להיות מוגבר. ה-RNA, טרנסקריפטאז הפוך, הפריימר וארבעה dNTPs מעורבים. עם סיבוב שכפול אחד, ה- RNA הופך לגדיל יחיד של DNA. דנטורציה משחררת את ה-cDNA החד-גדילי, שניתן להשתמש בו כפי שהוא, או להמיר ל-cDNA דו-גדילי, בהתאם ליישום.
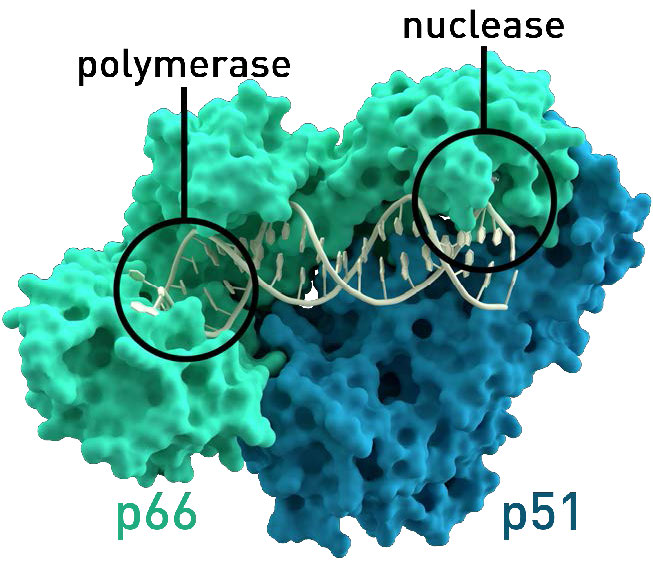
איור 8.37 - טרנסקריפטאז הפוך של HIV. פונקציית הגרעין נחוצה למחזור החיים הנגיפי, אך לא לשימוש במעבדה. ויקיפדיה
איתור אינטראקציות מולקולריות
חקר הביוכימיה הוא בעצם חקר האינטראקציות של מולקולות תאיות. שיטות לאיתור אינטראקציות בין ביומולקולות הן, מסיבה זו, שימושיות מאוד לביוכימאים. כעת נדון בכמה שיטות שונות מאוד לאיתור אינטראקציות בין-מולקולריות אלה.
מערכת שמרים דו-היברידית (Y2H)

בדיקת שמרים דו-היברידית היא טכניקה מתוחכמת לזיהוי אילו חלבונים, מתוך אוסף של כל חלבוני התא, מקיימים אינטראקציה עם חלבון ספציפי בעל עניין. השיטה מסתמכת על האינטראקציה בין שני חלבונים כדי לשחזר מפעיל תעתיק פונקציונלי בתוך תאי שמרים. אולי תזכור שמפעילי תעתיק רבים הם חלבונים מודולריים שיש להם תחום הנקשר ל-DNA ותחום אחר שמפעיל שעתוק (איור 8.38).
אם גורם השעתוק מפוצל, כך שתחום הקישור מחובר לחלבון אחד, ותחום ההפעלה לחלבון אחר, ניתן ליצור מחדש מפעיל תעתיק פונקציונלי רק אם שני חלבוני ה"נשא" מתקרבים - כלומר, הם מקיימים אינטראקציה. ניתן לזהות את נוכחותו של מפעיל פונקציונלי זה על ידי ביטוי של גן כתב.
דרך פשוטה להבין את הרעיון הזה היא לחשוב על מפעיל תעתיק כמכשיר, כמו פנס, שיש לו שני חלקים, הסוללה והמנורה, שחייבים להיות יחד כדי לתפקד. לא אדם שיש לו רק סוללה וגם לא מי שיש לו רק את המנורה לא יוכל לראות בחדר חשוך. אבל אם השניים מתקשרים על ידי התקרבות מספיק כדי להכניס את הסוללה לפנס, ניתן לזהות את האינטראקציה שלהם על ידי העובדה שהפנס יהיה פונקציונלי כעת כפי שמעיד האור המופק.
צריך שניים לטנגו
איור 8.39 (A) מציג את מפעיל תעתיק השמרים הרגיל, GAL4, עם תחומי הקישור ל-DNA (DBD) וההפעלה (AD). הוא מסוגל לעורר שעתוק של הגן הכתב במורד הזרם, לוחות B ו- C מראים מבנים המייצרים את GAL4 DBD ו- AD, בהתאמה, התמזגו לחלבונים אחרים, שאחד מהם מכונה "הפיתיון" והשני כ"טרף". אף אחד מחלבוני ההיתוך הללו לא יכול לעורר שעתוק של הגן lac z. כאשר מבנים המקודדים הן לפיתיון והן לטרף נמצאים באותו תא שמרים, אם חלבון הפיתיון מקיים אינטראקציה עם הטרף, ה-DBD וה-AD של ה-GAL4 יובאו יחד כדי ליצור מחדש GAL4 פונקציונלי. הנוכחות של GAL4 פונקציונלי ניתנת לזיהוי בקלות מכיוון שהיא תעורר ביטוי של הגן lac z reporter. אם חלבוני הפיתיון והטרף אינם מקיימים אינטראקציה, אז לא יהיה ביטוי lac z. כאשר אינטראקציה מזוהה באמצעות ביטוי של הגן הכתב, לאחר מכן ניתן לזהות את חלבון הטרף הספציפי.
מערכת השמרים הדו-היברידית מאפשרת סינון סימולטני של חלבוני טרף רבים, על ידי בניית אוספים גדולים של מבני היתוך, כאשר כל שותף חלבון פוטנציאלי של חלבון הפיתיון התמזג לתחום ההפעלה GAL4.
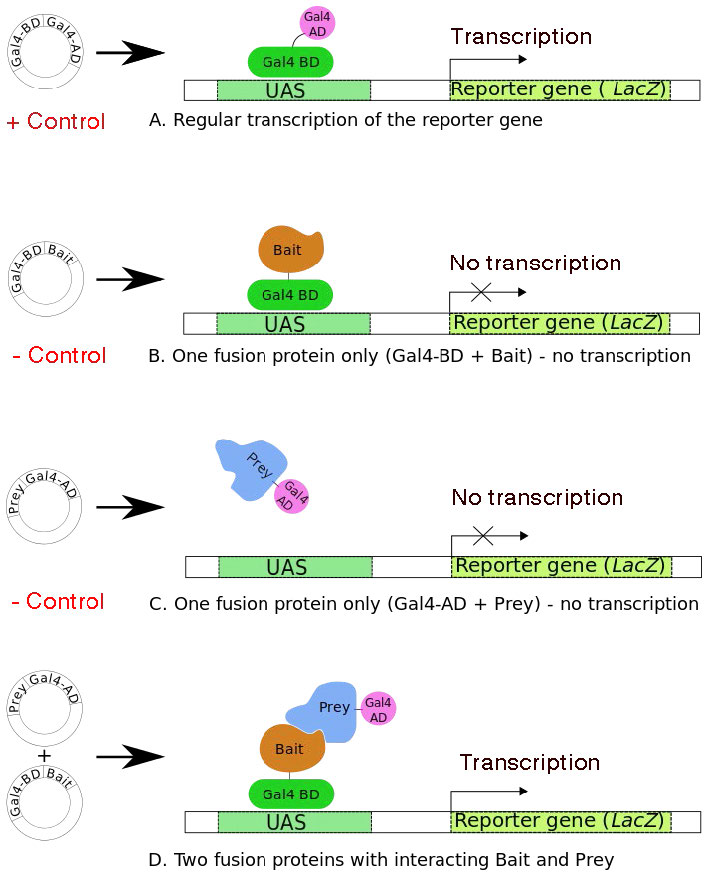
איור 8.39 - ארבעה תרחישים למערכת השמרים הדו-היברידית. UAS = רצפי מפעילים במעלה הזרם - מתנהג כמו מקדם. תרחיש A מראה ששני גורמי השעתוק מתחילים כחלבון אחד. ויקיפדיה