7.3: שכפול DNA
- Page ID
- 207549

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)מקור: BiochemFFA_7_2.pdf. ספר הלימוד כולו זמין בחינם מהמחברים בכתובת http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy

הוראות העתקה
הדרך היחידה ליצור תאים חדשים היא על ידי חלוקת תאים קיימים. אורגניזמים חד-תאיים עוברים חלוקה כדי לייצר יותר תאים כמוהם, בעוד שאורגניזמים רב-תאיים נוצרים באמצעות חלוקה של תא בודד, בדרך כלל הביצית המופרית. בכל פעם שתא מתחלק, יש להעתיק את כל ה- DNA שלו נאמנה כדי שניתן יהיה להעביר עותק של מידע זה לתא הבת. תהליך זה נקרא שכפול DNA. זהו האמצעי שבאמצעותו ניתן להעביר מידע גנטי לאורך דורות של תאים, והוא מבטיח שלכל תא חדש יהיה עותק מלא של הגנום. בחלק הבא נבחן את התהליך שבאמצעותו מועתק ה- DNA של התא באופן מלא ומדויק.
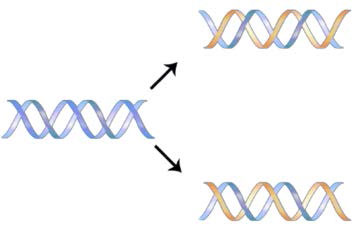
מבנה ה- DNA שהובהר על ידי ווטסון וקריק בשנת 1953 הציע מיד מנגנון שבאמצעותו ניתן להעתיק DNA דו-גדילי כדי לתת שני עותקים זהים של ה- DNA. הם הציעו ששני הגדילים של מולקולת ה-DNA, המוחזקים יחד על ידי קשרי מימן בין הנוקלאוטידים המשויכים לבסיס, ייפרדו וכל אחד מהם ישמש כתבנית שעליה ניתן להרכיב גדיל משלים (איור 7.8). כללי זיווג הבסיסים יבטיחו שתהליך זה יביא לייצור שתי מולקולות DNA זהות. הפשטות היפה של תכנית זו הוכחה כנכונה בניסויים הבאים של Meselson ו-Stahl, שהוכיחו כי שכפול ה-DNA היה שמרני למחצה, כלומר, שלאחר השכפול, כל אחת משתי מולקולות ה-DNA שהתקבלו מורכבת מגדיל ישן אחד וגדיל אחד חדש שהורכב מולו (איור 7.9).
חומרי בניין
מהם המרכיבים הדרושים לבניית מולקולת DNA חדשה? כפי שצוין לעיל, מולקולת ה-DNA המקורית או ההורית משמשת כתבנית. מולקולות DNA חדשות מורכבות מול כל תבנית על ידי חיבור נוקלאוטידים חופשיים של DNA לפי הוראות כללי זיווג הבסיסים, עם As מול Ts ו- Gs מול Cs.
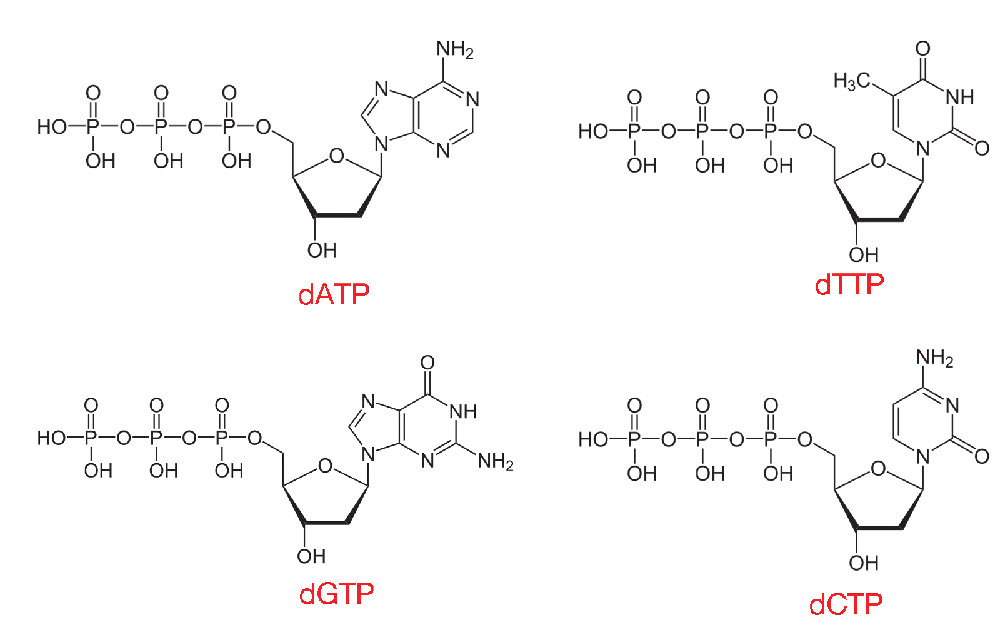
הנוקלאוטידים המשמשים בסינתזת DNA הם דאוקסיריבונוקלאוזיד טריפוספטים או dNTPs. כפי שניתן להסיק משמם, לנוקלאוטידים כאלה יש סוכר deoxyribose ושלושה פוספטים, בנוסף לאחד מארבעת בסיסי ה- DNA, A, T, C או G (איור 7.10).
כאשר dNTPs מתווספים לגדיל DNA גדל, שניים מאותם פוספטים יתנתקו, כמתואר מאוחר יותר, וישאירו את הנוקלאוטידים במולקולת DNA עם פוספט אחד בלבד לכל נוקלאוטיד. תגובה זו מזורזת על ידי אנזימים המכונים פולימראזות DNA, היוצרים קישורי פוספודיסטר בין נוקלאוטיד אחד למשנהו.
אתגרים
לפני בחינת התהליך בפועל של שכפול ה- DNA, כדאי לחשוב מה נדרש כדי לבצע משימה זו בהצלחה. שקול את האתגרים העומדים בפני תא בתהליך זה:
- המספר העצום של נוקלאוטידים שיש להעתיק הוא עצום: למשל, בתאים אנושיים, בסדר גודל של כמה מיליארדים.
- יש לפרוק מולקולת DNA הורית כפולה כדי לחשוף גדילים בודדים של DNA שיכולים לשמש תבניות לסינתזה של גדילי DNA חדשים.
- יש לבצע את ההתרה מבלי להכניס עיוות טופולוגי למולקולה.
- יש למנוע מהגדילים הבודדים של ה- DNA המפותלים לחזור יחד מספיק זמן כדי שהגדילים החדשים יסונתזו.
- פולימראזות DNA אינן יכולות להתחיל בסינתזה של גדיל DNA חדש דה נובו ודורשות OH 3' חופשי שאליו הן יכולות להוסיף deoxynucleotides.
- פולימראזות DNA יכולות להאריך גדיל רק בכיוון 5' עד 3'. הצמיחה של 5 'עד 3' של שני הגדילים החדשים פירושה שאחד הגדילים מיוצר בחלקים.
- השימוש בפריימרים של RNA מחייב להסיר את נוקלאוטידים ה-RNA ולהחליף אותם בנוקלאוטידים של DNA ולחבר את שברי ה-DNA המתקבלים.
- ההעתקה של כל ה- DNA ההורי חייבת להיות מדויקת, כך שלא יוכנסו מוטציות ל- DNA החדש שנוצר.

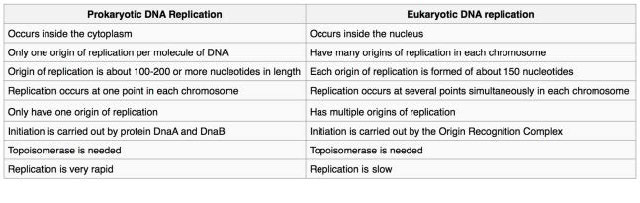
איור 7.14 - שכפול DNA פרוקריוטי לעומת אוקריוטי - ויקיפדיה
התמודדות עם אתגרים
עם זאת בחשבון, אנו יכולים להתחיל לבחון כיצד תאים מתמודדים עם כל אחד מהאתגרים הללו. ההבנה שלנו לגבי תהליך שכפול ה-DNA נגזרת ממחקרים המשתמשים בחיידקים, שמרים ומערכות אחרות. מחקירות אלה עולה כי שכפול ה- DNA מתבצע על ידי פעולה של מספר גדול של חלבונים הפועלים יחד כמכונת חלבון מורכבת. חלבונים רבים המעורבים בשכפול זוהו ואופיינו, כולל פולימראזות DNA שונות הן בפרוקריוטים והן באיקריוטים. למרות שהחלבונים הספציפיים המעורבים שונים בחיידקים ובאוקריוטים, כדאי להבין את השיקולים הבסיסיים הרלוונטיים בכל התאים. תיאור כללי של השלבים בשכפול ה-DNA מוצג להלן, המתמקד באתגרים שהוזכרו לעיל.
- המספר העצום של נוקלאוטידים שיש להעתיק הוא עצום: למשל, בתאים אנושיים, בסדר גודל של כמה מיליארדים.
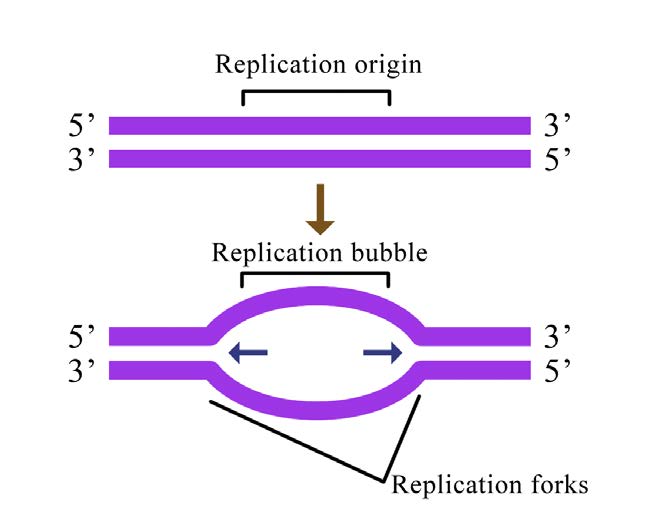
תאים, בין אם הם חיידקיים או אוקריוטים, צריכים לשכפל את כל ה- DNA שלהם לפני שהם יכולים להתחלק. בתאים כמו שלנו, הכמות העצומה של ה-DNA מתפרקת לכרומוזומים רבים, שכל אחד מהם מורכב מחוט DNA ליניארי (איור 7.12). בתאים כמו אלה של אי - קולי, יש כרומוזום מעגלי יחיד.
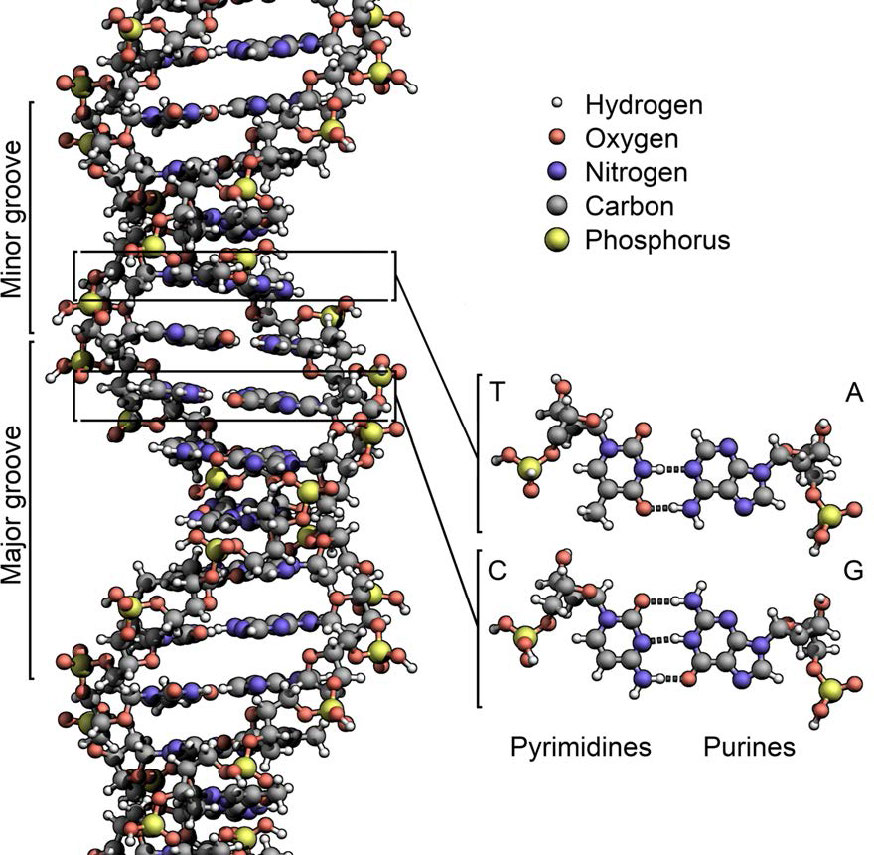
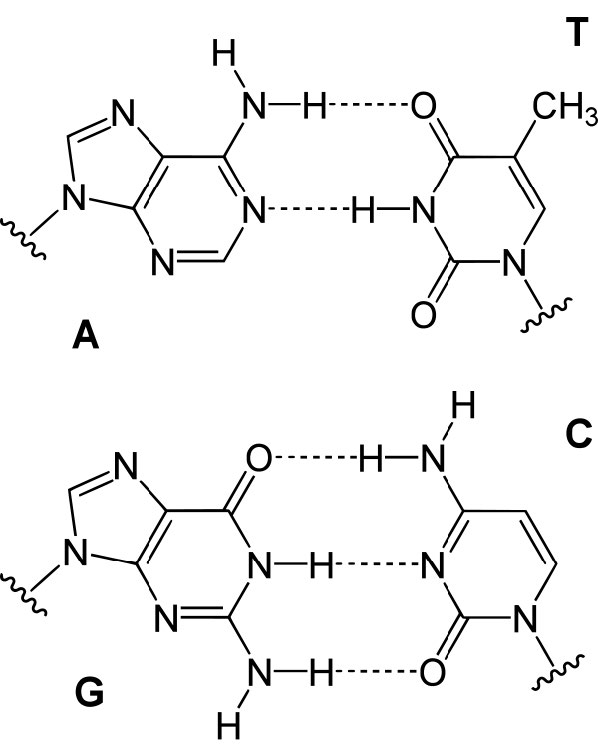
בשני המצבים, שכפול ה- DNA מתחיל באתרים הנקראים מקורות שכפול. אלה אזורים במולקולת ה- DNA המוכרים על ידי חלבונים מיוחדים הנקראים חלבוני יוזם הקושרים את ה- DNA. ב-E.coli, למקורות יש אזורים קטנים של רצפים עשירים ב-A-T ש"נמסים" כדי להפריד בין הגדילים, כאשר חלבוני היוזם נקשרים למקור או לשכפול. כפי שאתה אולי זוכר, זוגות בסיס A-T, שיש להם שני קשרי מימן ביניהם מופרעים בקלות רבה יותר מזוגות בסיס G-C שיש להם שלושה כל אחד (איור 7.15).
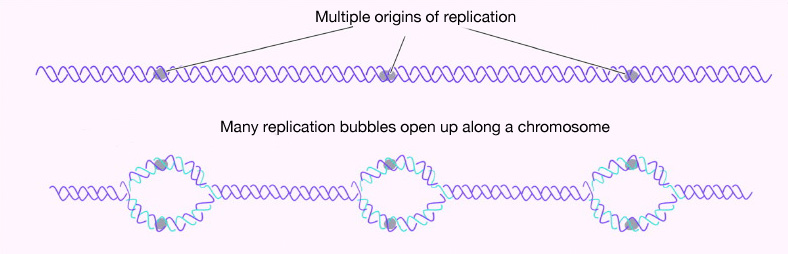
כמה מקורות שכפול יש בכרומוזום? במקרה של E. coli, יש מקור שכפול יחיד בכרומוזום המעגלי שלו. בתאים אוקריוטים עשויים להיות אלפים רבים של מקורות שכפול, כאשר לכל כרומוזום יש מאות (איור 7.16). שכפול ה- DNA מתחיל, אם כן, במספר נקודות לאורך כל כרומוזום באיקריוטים. מיקרוגרפים אלקטרונים של DNA משכפל מתאי אוקריוטים מראים בועות שכפול רבות על כרומוזום בודד. זה הגיוני לאור כמות ה-DNA הגדולה שיש להעתיק בתאים כמו שלנו, כאשר התחלה בקצה אחד של כל כרומוזום ושכפול עד לקצה השני ממקור יחיד פשוט ייקח יותר מדי זמן. זאת למרות העובדה שפולימראזות ה-DNA בתאים אנושיים מסוגלות לבנות גדילי DNA חדשים בקצב מכובד מאוד של כ-50 נוקלאוטידים בשנייה!
- יש לפתוח מולקולת הורים כפולה סלילית כדי לחשוף גדילים בודדים של DNA שיכולים לשמש תבניות לסינתזה של גדילי DNA חדשים.
הרפה
ברגע שנפתח אזור קטן של ה-DNA בכל מקור שכפול, יש לפתוח את סליל ה-DNA כדי לאפשר לשכפול להמשיך. פירוק סליל ה- DNA דורש פעולה של אנזים הנקרא הליקאז.
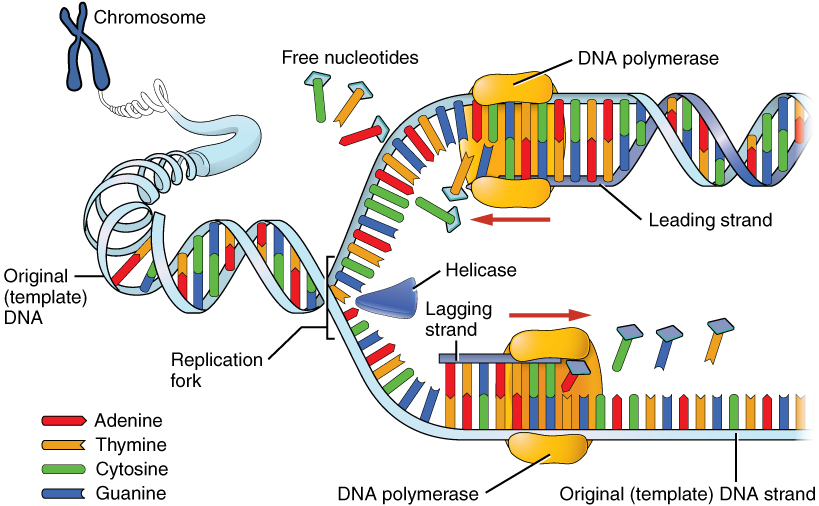
Helicase משתמש באנרגיה המשתחררת כאשר ATP עובר הידרוליזה, כדי לשבור את קשרי המימן בין הבסיסים ב-DNA ולהפריד בין שני הגדילים (איור 7.17). שים לב שבועת שכפול מורכבת משני מזלגות שכפול ש"זזים "או נפתחים, בכיוונים מנוגדים. בכל מזלג שכפול, יש לפתוח את גדילי ה-DNA ההוריים כדי לחשוף קטעים חדשים של תבנית חד-גדילית.
- הרפיה זו חייבת להתבצע מבלי להכניס עיוות טופולוגי למולקולה.
מהי ההשפעה של פירוק אזור אחד של הסליל הכפול? התנתקות מקומית של הסליל הכפול גורמת לפיתול יתר (סליל-על חיובי מוגבר) לפני האזור המתפרק.
ה-DNA שלפני מזלג השכפול צריך להסתובב, או שהוא יתעוות על עצמו ויעצור את השכפול. זוהי בעיה מרכזית, לא רק לכרומוזומים חיידקיים מעגליים, אלא גם לכרומוזומים אוקריוטיים ליניאריים, אשר באופן עקרוני יכולים להסתובב כדי להקל על הלחץ הנגרם כתוצאה מסליל העל המוגבר.
טופואיזומראזות
הסיבה שזה בעייתי היא שלא ניתן לסובב את כל אורכו של הכרומוזום, על מיליוני זוגות הבסיס שלו, מכיוון שה-DNA במזלג השכפול מתפרק. כיצד, אם כן, נפתרה בעיה זו? אנזימים הנקראים טופואיזומראזים יכולים להקל על הלחץ הטופולוגי הנגרם על ידי "התנתקות" מקומית של הרוחות הנוספות של הסליל הכפול. הם עושים זאת על ידי חיתוך אחד או שני גדילי ה- DNA ומאפשרים לחוטים להסתובב זה סביב זה כדי לשחרר את המתח לפני שהם מצטרפים מחדש לקצוות. ב- E. coli, הטופואיזומראז שמבצע פונקציה זו נקרא gyrase.
- יש למנוע מהגדילים הבודדים המופרדים של ה- DNA לחזור זה לזה כדי לסנתז את הגדילים החדשים.
חלבון קושר DNA חד-גדילי
ברגע ששני הגדילים של מולקולת ה-DNA ההורית מופרדים, יש למנוע מהם לחזור יחד ליצירת DNA דו-גדילי. כדי להבטיח שאזורים לא מפותלים של ה-DNA ההורי יישארו חד-גדילים וזמינים להעתקה, הגדילים המופרדים של ה-DNA ההורי נקשרים למולקולות רבות של חלבון הנקרא חלבון מחייב DNA חד-גדילי (SSB - איור 7.18).
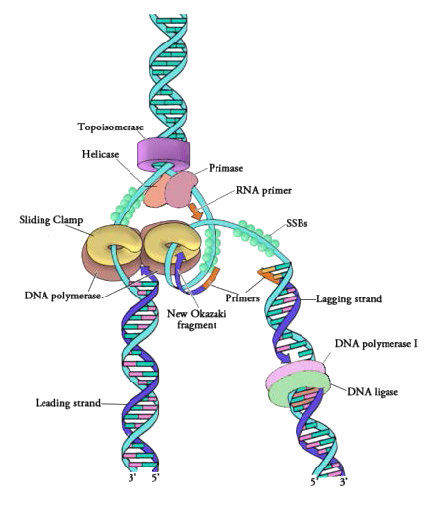
איור 7.18 - חלבונים במזלג שכפול DNA פרוקריוטי - תמונה מאת מרתה בייקר
- פולימראזות DNA אינן יכולות להתחיל בסינתזה של גדיל DNA חדש דה נובו ודורשות OH 3' חופשי שאליו הן יכולות להוסיף נוקלאוטידים של DNA.
למרות ש- DNA הורי חד-גדילי זמין כעת להעתקה, פולימראזות DNA אינן יכולות להתחיל בסינתזה של גדיל משלים דה נובו. זה פשוט אומר שפולימראזות DNA יכולות להוסיף נוקלאוטידים חדשים רק לקצה 3' של שרשרת קיימת, ואינן יכולות להתחיל שרשרת של נוקלאוטידים בכוחות עצמן. בגלל מגבלה זו, אנזים כלשהו מלבד פולימראז DNA חייב ליצור תחילה אזור קטן של חומצת גרעין, המשלים לגדיל ההורי, שיכול לספק 3' OH חופשי שאליו פולימראז DNA יכול להוסיף deoxyribonucleotide. משימה זו מתבצעת על ידי אנזים הנקרא פרימאז, המרכיב קטע קצר של בסיס RNA המותאם לתבנית ה- DNA ההורית. זה מספק אזור מזווג בסיס קצר, הנקרא פריימר RNA, עם קבוצת 3'OH חופשית שאליה פולימראז DNA יכול להוסיף את נוקלאוטיד ה- DNA החדש הראשון (איור 7.12).
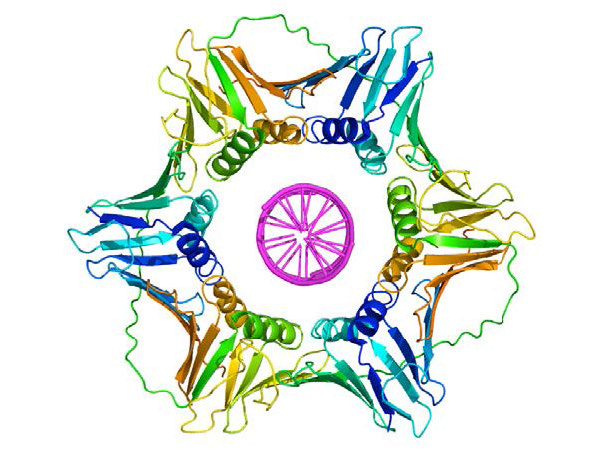
מהדק הזזה
ברגע שפריימר מספק 3'OH בחינם להארכה, חלבונים אחרים נכנסים לפעולה. חלבונים אלה מעורבים בהעמסת ה-DNA פולימראז על התבנית המוכנה ושמירה עליו קשור ל-DNA. הראשון שבהם הוא מעמיס מהדק. כפי ששמו מרמז, מעמיס המהדק עוזר להעמיס קומפלקס חלבון הנקרא מהדק הזזה על ה-DNA במזלג השכפול (איור 7.19 ו-7.20). מהדק ההזזה, חלבון בצורת טבעת מרובת יחידות משנה, מצטרף לאחר מכן על ידי ה- DNA Polymerase. תפקידו של מהדק הזזה הוא לשמור על הפולימראז הקשור למזלג השכפול - למעשה, הוא תואר כחגורת בטיחות לפולימראז ה- DNA. מהדק הזזה מבטיח כי פולימראז ה- DNA מסוגל לסנתז קטעים ארוכים של DNA חדש לפני שהוא מתנתק מהתבנית. המאפיין של להישאר קשור לתבנית זמן רב לפני הניתוק ידוע כתהליכי האנזים. בנוכחות מהדק הזזה, פולימראזות DNA הן הרבה יותר עיבוד, מה שהופך את השכפול למהיר ויעיל יותר.
הארכת הפריימר
פולימראז ה- DNA מוכן כעת להתחיל בסינתזה של גדיל ה- DNA החדש (ב אי - קולי, הפולימראז המשכפל העיקרי נקרא DNA פולימראז III). כפי שאתה כבר יודע, הסינתזה של DNA חדש מושגת על ידי הוספת נוקלאוטידים חדשים המשלימים לאלה שעל גדיל ההורים. פולימראז DNA מזרז את התגובה שבאמצעותה מתווסף דאוקסיריבונוקלאוטיד נכנס, המשלים לתבנית, לקצה 3 'של הנוקלאוטיד הקודם, החל מה- 3'OH בקצה פריימר ה- RNA. חשיבותה של קבוצת 3'OH נעוצה באופי התגובה הבונה שרשרת נוקלאוטידים.
התגובה המזרזת על ידי פולימראז ה- DNA מתרחשת באמצעות ההתקפה הנוקלאופילית על ידי קבוצת 3'OH בקצה גדיל חומצת גרעין על הפוספט α של ה- dNTP הנכנס (איור 7.21). ההידרוליזה המיידית של הפירופוספט שנבקע מה- dNTP הנכנס מניעה את התגובה קדימה. התוספת הרציפה של נוקלאוטידים חדשים בקצה 3' של שרשרת ה-DNA ההולכת וגדלה מסבירה את העובדה שהגדיל גדל בכיוון 5' עד 3'.
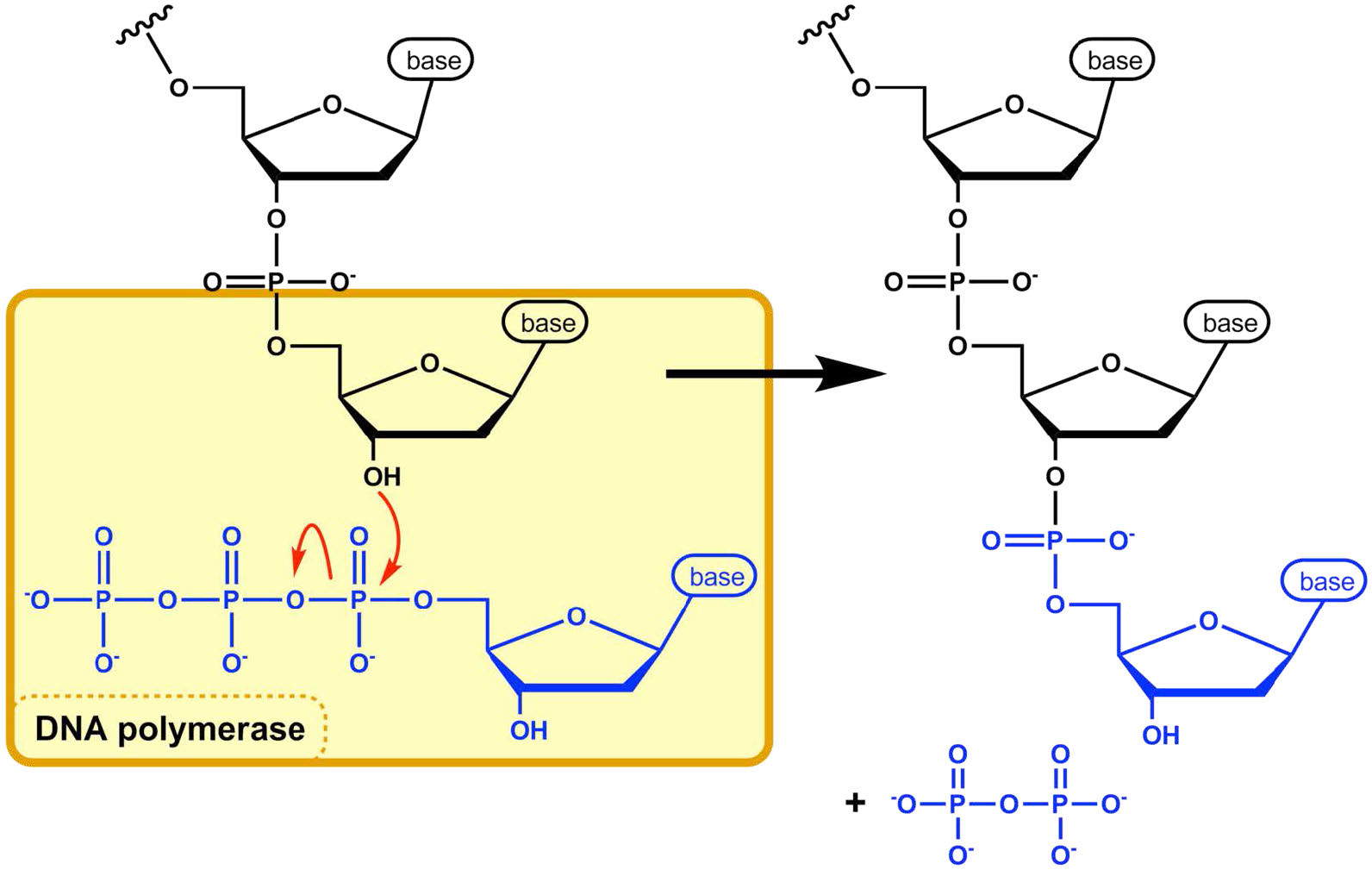
הפוספט 5' על כל נוקלאוטיד נכנס מצטרף על ידי פולימראז ה-DNA ל-3' OH בקצה שרשרת חומצות הגרעין הגדלה, כדי ליצור קשר פוספודיסטר. כל נוקלאוטיד שנוסף מספק 3'OH חדש, המאפשר להאריך את השרשרת כל עוד פולימראז ה- DNA ממשיך לסנתז את הגדיל החדש. כפי שכבר ציינו, גדילי ה- DNA החדשים מסונתזים על ידי הוספת נוקלאוטידים של DNA לקצה פריימר RNA. למולקולת ה- DNA החדשה יש אפוא חתיכת RNA קצרה בהתחלה.
- פולימראזות DNA יכולות להאריך גדיל רק בכיוון 5' עד 3'. הצמיחה של 5 'עד 3' של שני הגדילים החדשים פירושה שאחד הגדילים מיוצר בחלקים.
גדיל מוביל
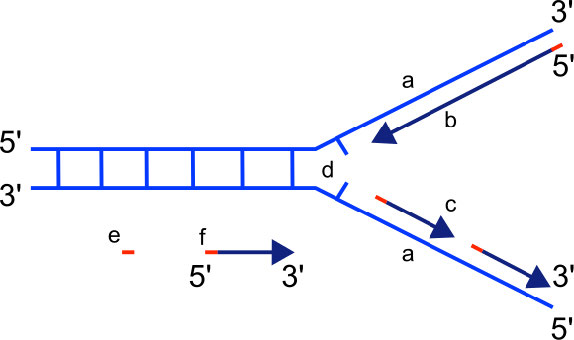
אנו יודעים שפולימראזות DNA יכולות לבנות רק גדיל DNA חדש בכיוון 5' עד 3'. אנו גם יודעים ששני גדילי ה- DNA ההוריים הם אנטי מקבילים. המשמעות היא שבכל מזלג שכפול, ניתן לסנתז גדיל חדש אחד, הנקרא גדיל מוביל ברציפות בכיוון 5 'עד 3' מכיוון שהוא מיוצר באותו כיוון שמזלג השכפול נפתח.
גדיל בפיגור
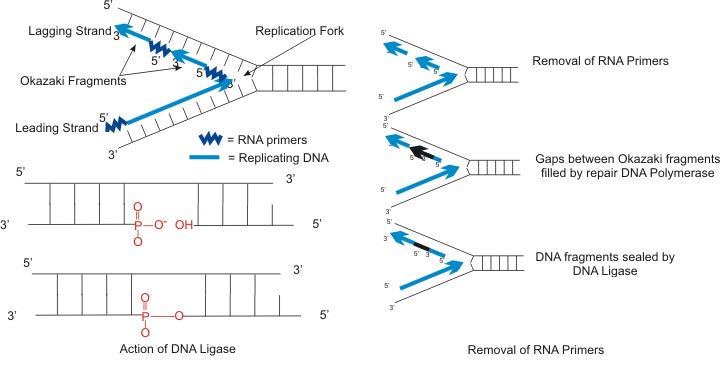
הסינתזה של הגדיל החדש השני, הנקרא גדיל הפיגור, ממשיכה גם בכיוון 5' עד 3'. אך מכיוון שגדילי התבנית פועלים בכיוונים מנוגדים, הגדיל הפיגור מורחב בכיוון ההפוך לפתיחת מזלג השכפול (איור 7.22). כאשר מזלג השכפול נפתח, יהיה צורך להעתיק את האזור שמאחורי נקודת ההתחלה המקורית של הגדיל המפגר. המשמעות היא שיש להניח ולהרחיב פריימר RNA נוסף. תהליך זה חוזר על עצמו כאשר מזלג השכפול נפתח, עם מספר פריימרים של RNA מונחים ומורחבים, ומייצרים חלקים קצרים רבים המחוברים מאוחר יותר. חלקי חומצת הגרעין הקצרים הללו, שכל אחד מהם מורכב מקטע קטן של פריימר RNA וכ- 1000-2000 נוקלאוטידים של DNA, נקראים שברי אוקזאקי, עבור רייג'י אוקזאקי, המדען שהוכיח לראשונה את קיומם.
- השימוש בפריימרים של RNA מחייב להסיר את נוקלאוטידים ה- RNA ולהחליף אותם בנוקלאוטידים של DNA.
הסרת פריימר
ראינו שכל פיסת DNA שסונתזה לאחרונה מתחילה עם פריימר RNA, ויוצרת למעשה גדיל חומצת גרעין חדש שהוא חלק RNA וחלק DNA. לא ניתן לאפשר לגדיל ה- DNA החדש שנוצר לחבר חתיכות RNA. לכן, יש להסיר את נוקלאוטידים ה-RNA ולמלא את הפערים בנוקלאוטידים של DNA (איור 7.23). זה נעשה על ידי DNA פולימראז I ב אי - קולי. אנזים זה מתחיל להוסיף נוקלאוטידים של DNA בסוף כל שבר Okazaki. עם זאת, סופו של שבר אוקאזאקי אחד צמוד לפריימר ה- RNA בתחילת שבר אוקאזאקי הבא. ל-DNA פולימראז I יש פעילות אקסונוקלאז הפועלת בכיוון 5' עד 3' שמסירה את נוקלאוטידים ה-RNA לפניו, בעוד שפעילות הפולימראז מחליפה את נוקלאוטידים ה-RNA ב-dNTPs. לאחר הסרת כל נוקלאוטידים ה- RNA, הגדיל המפגר מורכב מקטעי DNA. לאחר מכן מחברים את חלקי ה- DNA על ידי האנזים DNA ligase.
השלבים המתוארים לעיל משלימים בעצם את תהליך שכפול ה- DNA. אבל נושא אחד עדיין נשאר.
- הבטחת דיוק בהעתקה של כל כך הרבה מידע
דיוק
עד כמה מדויקת העתקת המידע על ידי פולימראז DNA? כפי שאתה מודע, שינויים ברצף ה-DNA (מוטציות) יכולים לשנות את רצף חומצות האמינו של החלבונים המקודדים וכי זה לעתים קרובות, אם כי לא תמיד, מזיק לתפקוד האורגניזם. כאשר מיליארדי בסיסים ב-DNA מועתקים במהלך השכפול, כיצד תאים מבטיחים שה-DNA החדש שסונתז הוא עותק נאמן של המידע המקורי?
פולימראזות DNA, כפי שציינו קודם לכן פועלות מהר (בממוצע 50 בסיסים בשנייה בתאים אנושיים ועד פי 200 מהר יותר ב אי - קולי). עם זאת, נראה שתאים אנושיים וחיידקיים משכפלים את ה-DNA שלהם בצורה מדויקת למדי. הסיבה לכך היא שלפולימראזות DNA רפליקטיביות יש פונקציית הגהה המאפשרת לפולימראז לזהות מתי הוכנס הבסיס הלא נכון מול גדיל תבנית, לגבות ולהסיר את הבסיס שהוכנס בטעות, לפני שתמשיך בסינתזה (איור 7.24).
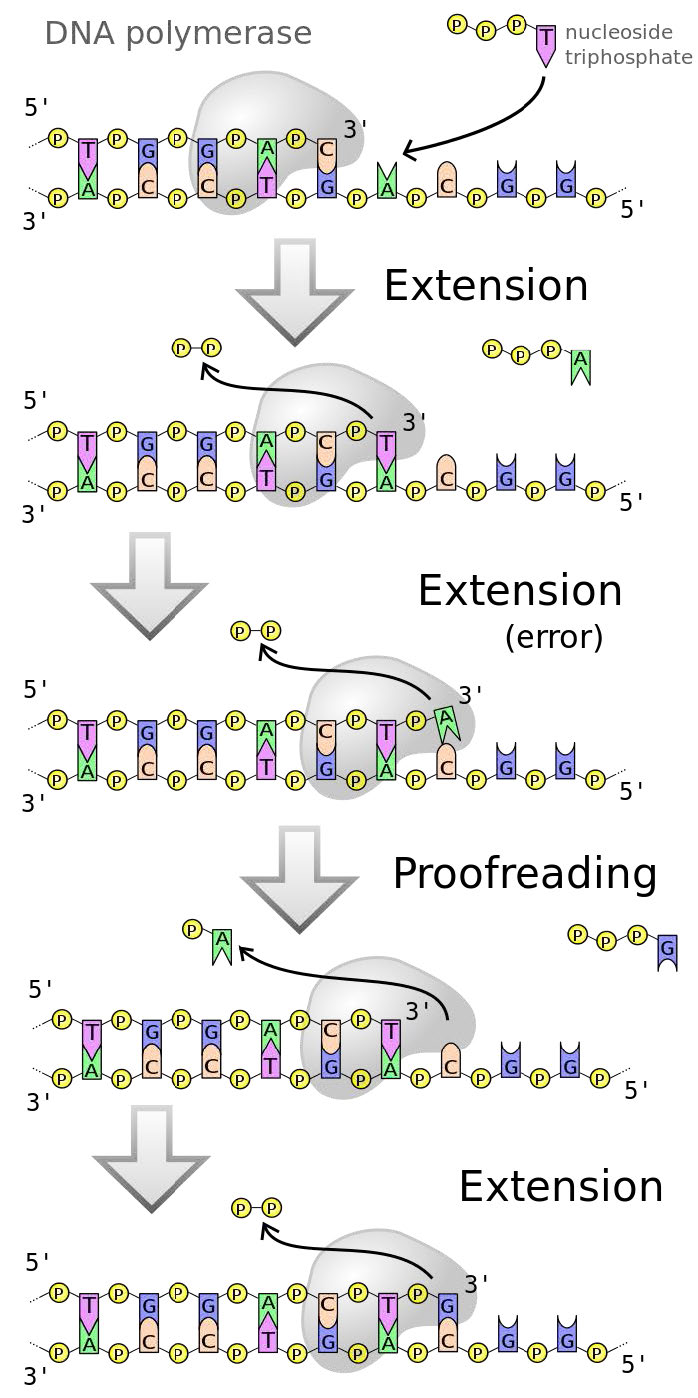
איור 7.24 - שגיאה שתוקנה על ידי פולימראזות DNA
פעילויות מרובות
זה אפשרי מכיוון שרוב פולימראזות ה- DNA הן אנזימים בעלי תפקוד כפול. הם יכולים להאריך שרשרת DNA מכוח פעילות הפולימראז 5' עד 3' שלהם. כמה פולימראזות כמו DNA פולימראז I יכולות גם להסיר פריימרים של RNA בכיוון 5' עד 3', אם כי זו אינה פעילות נפוצה של פולימראזות. עם זאת, לפולימראזות רבות יש יכולת לחזור אחורה ולהסיר את הבסיס שהוכנס האחרון מכיוון שיש להן פעילות אקסונוקלאז של 3 'עד 5'.
פעילות האקסונוקלאז של פולימראז DNA מאפשרת לו לכרות בסיס שהוכנס בצורה לא נכונה, ולאחר מכן פעילות הפולימראז מכניסה את הבסיס הנכון וממשיכה בהרחבת הגדיל.
במילים אחרות, ה-DNA פולימראז עוקב אחר הדיוק שלו (המכונה גם נאמנותו) כשהוא יוצר DNA חדש, מתקן טעויות מיד לפני שהוא ממשיך להוסיף את הבסיס הבא. מנגנון זה, הפועל במהלך שכפול ה- DNA, מתקן שגיאות רבות כשהן מתרחשות, ומפחית בערך פי 100 את הטעויות שנעשו בעת העתקת ה- DNA.
פולימראזות DNA
כפי שצוין קודם לכן, לתאים פרוקריוטים ואיקריוטים יש פולימראזות DNA מרובות. ב- E.coli, למשל, DNA פולימראז III הוא הפולימראז המשכפל העיקרי (aka replicase) בעוד ש- DNA פולימראז I אחראי לתיקון DNA כמו גם להסרת פריימרים של RNA והחלפתם בנוקלאוטידים של DNA במהלך השכפול. DNA פולימראז II ממלא תפקיד בהפעלה מחדש של שכפול לאחר שנזק ל-DNA מעכב את השכפול, בעוד שפולימראזות DNA IV ו-V נדרשות שניהם בסינתזה טרנס-נגע, או מעקף, המאפשרת שכפול בעבר של אתרים של נזק ל-DNA.
פולימראזות אוקריוטיות
באיקריוטים, ישנם למעלה מחמישה עשר פולימראזות DNA שונות. הפולימראזות המשכפלות העיקריות בגרעין הן ו ε. DNA פולימראז α חשוב גם לשכפול מכיוון שיש לו פעילויות פרימאז ותיקון. השכפול מתחיל בתאים אוקריוטיים על ידי DNA פולימראז α, הנקשר לקומפלקס ההתחלה במקור ומניח פריימר RNA, ואחריו כ-25 נוקלאוטידים של DNA. לאחר מכן הוא מוחלף בפולימראז אחר, בשלב הנקרא מתג פול. DNA פולימראז או ε ממשיך לסנתז DNA, בהתאם לגדיל. נראה כי תפקידו של פולימראז ε הוא סינתזה של הגדיל המוביל בשל התהליכות והדיוק הגבוהים שלו, ואילו פולימראז מרחיב שברי Okazaki על הגדילים המפגרים. חלבונים מקבילים למעמיס המהדק ומהדק הזזה קיימים גם הם. החלבון RFC ממלא את התפקיד של מעמיס מהדק, בעוד חלבון אחר, PCNA פועל כמו מהדק הזזה. מספר פולימראזות DNA אחרות כמו β, γ ו- μ מתפקדות בתיקון פערים. עם זאת, אחרים מעורבים בסינתזת טרנס-נגע בעקבות נזק ל-DNA וקשורים להיפרמוטציה.
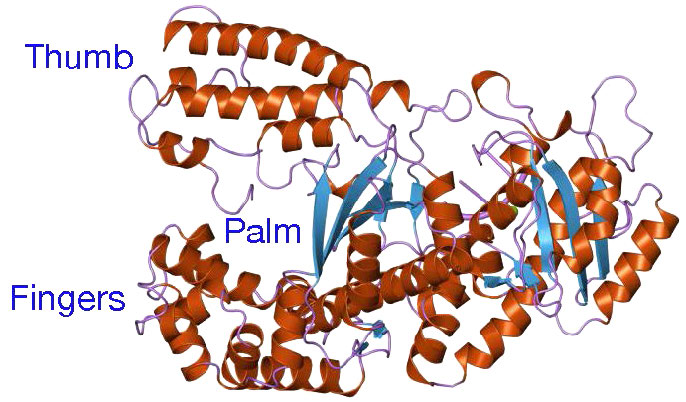
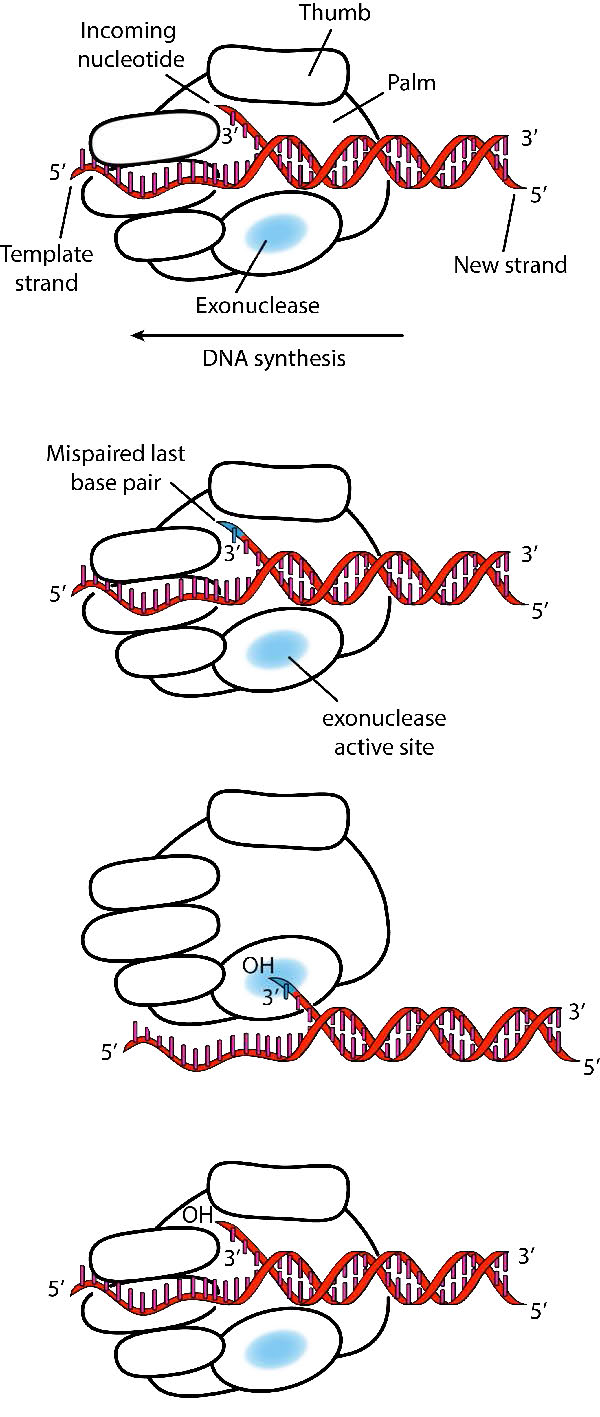
למרות המגוון שלהם, פולימראזות DNA חולקות כמה תכונות מבניות נפוצות. מחקרים קריסטלוגרפיים בקרני רנטגן הראו שלאנזימים אלו יש מבנה שהושווה ליד ימין אנושית (איורים 7.25 ו-7.26). "כף היד" של היד יוצרת שסע בו נמצא ה- DNA. השסע הוא גם המקום בו שוכנת הפעילות הקטליטית של הפולימראז. זה המקום שבו הנוקלאוטיד הנכנס מתווסף לשרשרת ההולכת וגדלה. "האצבעות" ממקמות את ה-DNA באתר הפעיל, בעוד ה"אגודל" מחזיק את ה-DNA כשהוא יוצא מהפולימראז. תחום נפרד מכיל את פעילות האקסונוקלאז (הגהה) של האנזים.
האנזים מתחלף בין פעילות הפילמור שלו לבין פעילות ההגהה שלו. כאשר זוג בסיסים לא תואם נמצא באתר הקטליטי של הפולימראז, קצה ה-3' של הגדיל הגדל מועבר מאתר הפולימראז לאתר הפעיל של האקסונוקלאז (איור 7.26). חוסר ההתאמה בסוף מוסר על ידי האקסונוקלאז, ואחריו מיקום מחדש של קצה 3' באתר הפעיל של הפולימראז כדי להמשיך בסינתזה.
סיום שכפול
בכרומוזומים חיידקיים מעגליים, ישנם רצפים ספציפיים המכונים אתרי טרמינטור או Ter. אלו הם רצפים קצרים מרובים המשמשים כאתרי סיום, המאפשרים למזלגות השכפול הנעים בכיוון השעון ונגד כיוון השעון על פני הכרומוזום המעגלי להיפגש באחד האתרים.
הקישור של חלבון, Tus, באתר Ter מונע תנועה נוספת של מזלג השכפול ומסיים את השכפול. ה- DNA המעגלי ההורי והחדש שנוצר קשורים זה בזה בשלב זה טופולוגית ויש להפריד אותם בעזרת טופואיזומראז.
בעיית השכפול הסופי
אין אתר קבוע לסיום בכרומוזומים אוקריוטיים ליניאריים. כאשר מזלגות השכפול מגיעים לקצוות הכרומוזום, ניתן לסנתז את הגדיל המוביל עד לקצה גדיל התבנית. על הגדיל המפגר, הצורך בפריימר RNA כדי להתחיל בסינתזה יוצר אתגר. כאשר מסירים את פריימר ה-RNA בקצה הקיצוני של הגדיל המפגר, יש מתיחה קטנה של גדיל התבנית שלא ניתן להעתיק. כתוצאה מכך, בכל סבב שכפול יאבד רצף קצר בקצות הכרומוזום. עם הזמן, עם מחזורי שכפול רבים, הכרומוזומים יתקצרו באופן ניכר. קיצור זה של הכרומוזומים נצפה במבחנה, בתאים סומטיים של יונקים מתורבתים. זה נראה גם באורגניזמים שלמים, עם העלייה בגיל.
טלומרים
איזו השפעה יש לאובדן הרצף מקצות הכרומוזומים על התאים? אנו יודעים שקצות הכרומוזומים מאופיינים במבנים הנקראים טלומרים (איור 7.28). הטלומרים מורכבים מעותקים רבים של רצפים חוזרים ונשנים קצרים (בבני אדם, החזרה היא TTAGGG) וחלבונים מיוחדים הנקשרים באופן ספציפי לרצפים אלה. מבנה זה של טלומרים שימושי בהבחנה בין קצות הכרומוזומים לבין שבירות כפולות ב-DNA, ובכך מונע ממנגנוני תיקון ה-DNA בתאים להצטרף לכרומוזומים מקצה לקצה.
היתרון הנוסף של הרצפים החוזרים, שאינם מקודדים לחלבונים, הוא שאיבוד חלק מהחזרות אינו מוביל לאובדן מידע קידוד חשוב. לפיכך, החזרות פועלות כמעין אזור חיץ, שבו אובדן הרצף אינו גורע לתא. עם זאת, קיצור הכרומוזומים אינו יכול להימשך ללא הגבלת זמן. לאחר מספר מסוים של מחזורי שכפול, ידוע שהתאים מפסיקים להתחלק ונכנסים למצב המכונה הזדקנות רפליקטיבית. זה מצביע על כך שקיצור הטלומרים משמש מעין שעון, כאשר מידת הצטמקות הכרומוזומים משמשת מדד להזדקנות. בסופו של דבר תאים שנכנסים להזדקנות ימותו.
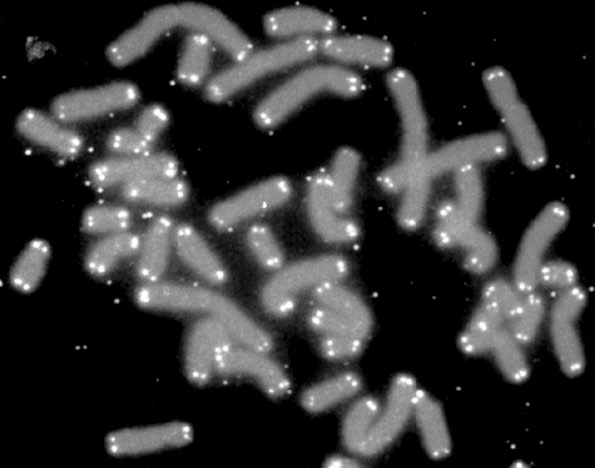
איור 7.28 - כרומוזומים עם טלומרים מסומנים בלבן
בעיות עם אובדן רצף
גם אם התאים שלנו מסוגלים לתפקד עם כרומוזומים קצרים יותר במהלך חיינו, הדבר מותיר אותנו עם בעיה נוספת. אם הכרומוזומים שלנו יתקצרו עם הגיל, אז יש להניח שילדינו, שירשו את הכרומוזומים שלנו ייוולדו עם כרומוזומים קצרים יותר ממה שהתחלנו איתם. הם, בתורם, היו מתכווצים הכרומוזומים שלהם ככל שהם מתבגרים, ולילדיהם יהיו כרומוזומים קצרים עוד יותר. במהלך דורות מרובים, זה יוביל לנקודה שבה הצטמקות כרומוזומים נוספת תגרום לתאים שייכנסו להזדקנות מוקדם מאוד בחיים וימותו זמן קצר לאחר מכן. ברור שזה לא קורה. דור אחר דור של ילדים נולדים עם כרומוזומים באורך מלא, ולכן יש מנגנון שחייב להבטיח שלפחות בתאי הרבייה הכרומוזומים לא יתקצרו.
כדי להבין את המנגנון הזה, יש צורך לבחון תחילה את סופה של מולקולת DNA חדשה שנוצרה (איור 7.29). בעוד שהגדיל המוביל, הגדל באותו כיוון כמו תנועת מזלג השכפול, יכול להעתיק את התבנית שלו עד הסוף, הגדיל המפגר נתקל בבעיה. פריימרים של RNA נחוצים, כפי שציינו, כדי להתחיל כל אחד משברי Okazaki של הגדיל המפגר. יש להסיר את הפריימרים מאוחר יותר, ולהחליף את נוקלאוטידים ה- RNA בנוקלאוטידים של DNA. כאשר מסירים את פריימר ה-RNA מול קצה הגדיל ההורי, לא ניתן להחליף את נוקלאוטידים ה-RNA בנוקלאוטידים של DNA מכיוון שלפולימראז ה-DNA אין פריימר להתחיל ממנו. לכן לא ניתן להעתיק אזור קצר בתבנית.
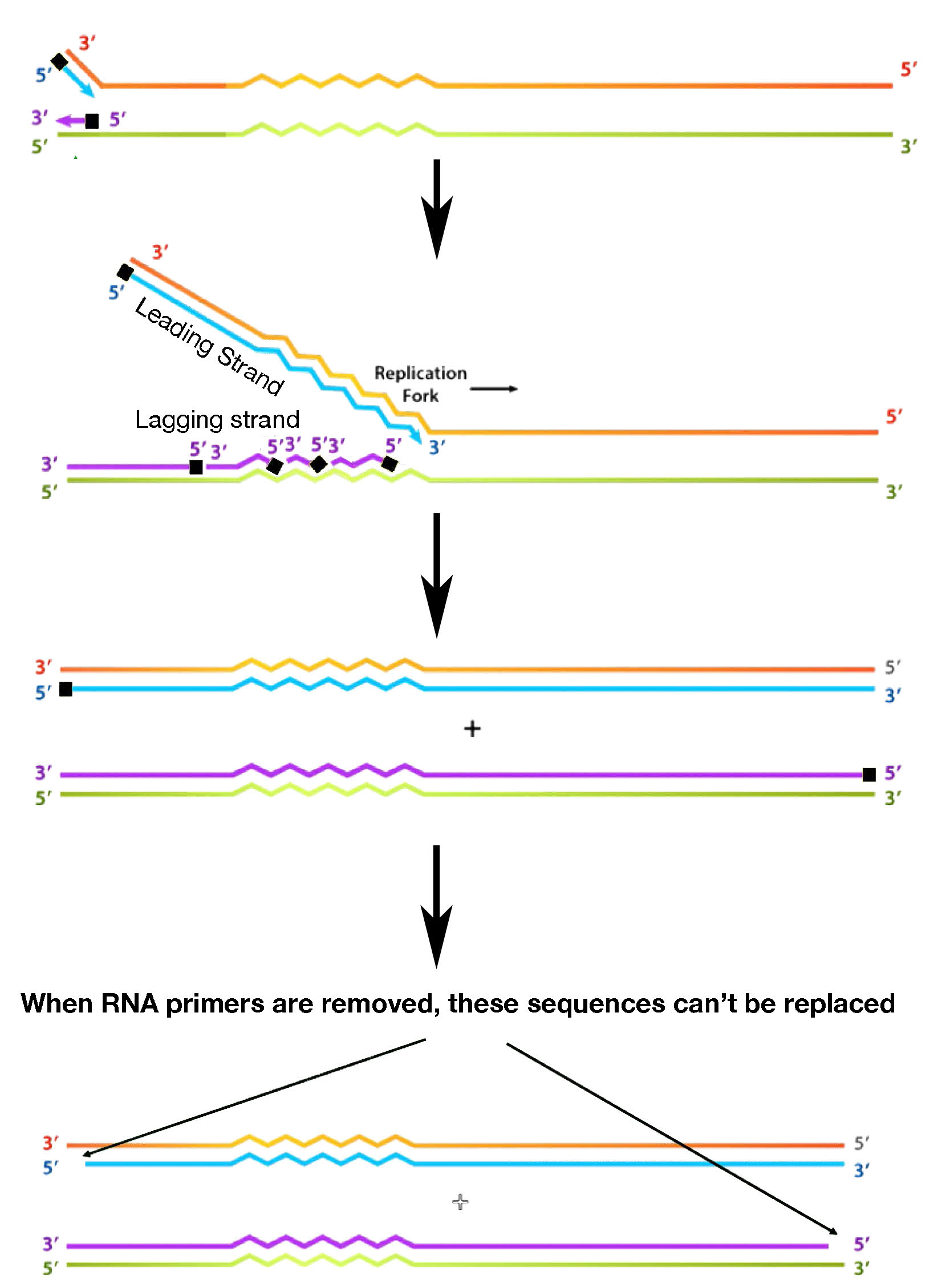
איור 7.29 - שכפול של כרומוזום ליניארי גורם לאובדן רצפים ממש בקצוות עם כל סבב שכפול
טלומראז
כיצד ניתן לפתור בעיה זו? ניתן לראות מאיור 7.29 שלקצה גדיל התבנית המקורי יש תלייה קצרה של 3 'הנובעת מהסרת פריימר ה-RNA מולו. על מנת למלא אזור זה, יהיה צורך בפריימר נוסף, הממוקם מעבר לקצה גדיל התבנית. אך על מנת לבנות פריימר כזה, יהיה צורך שתליית התבנית תהיה ארוכה יותר. אם ניתן היה להאריך את גדיל התבנית, ניתן היה למקם פריימר נוסף מול קצהו ולהעתיק את קצה הגדיל. הרחבה כזו של גדיל התבנית היא בדיוק מה שקורה בתאי הרבייה שלנו. גדיל התבנית ההורית מורחב על ידי האנזים טלומראז, שמוסיף חזרות טלומרים ומאריך את התבנית. נראה בקרוב כיצד הוא משיג את ההישג הזה.
תבנית RNA
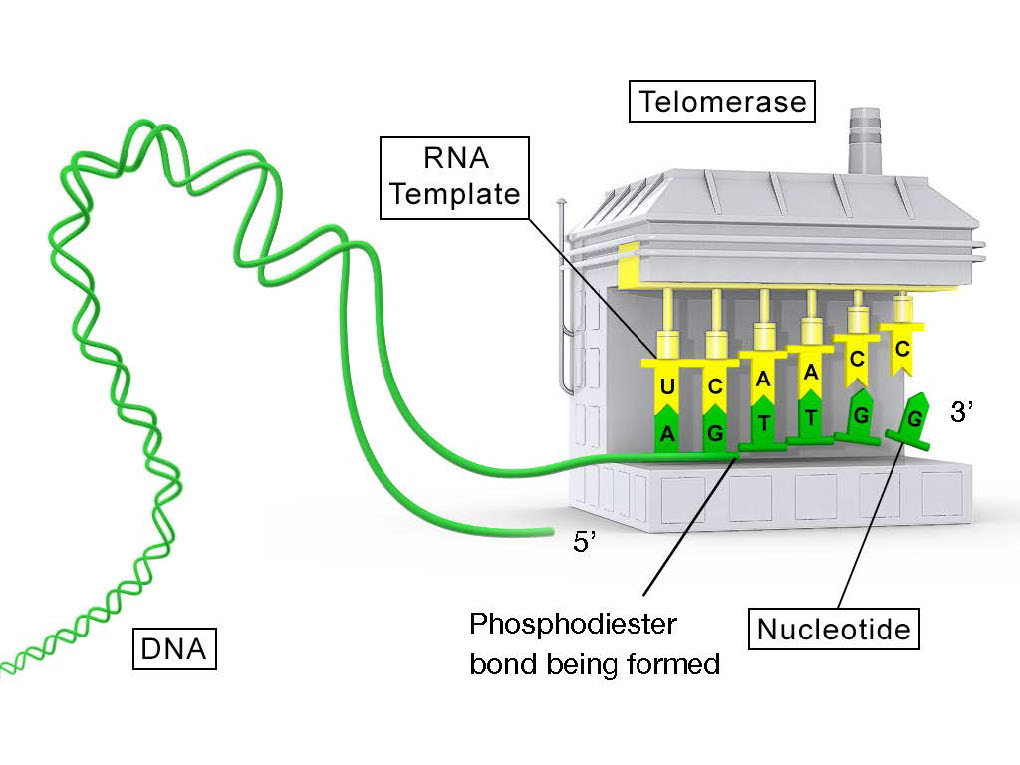
טלומראז הוא אנזים יוצא דופן, בכך שהוא מורכב משני רכיבים, RNA ותעתיק הפוך. טרנסקריפטאז הפוך הוא פולימראז DNA תלוי RNA, אנזים המעתיק תבנית RNA ליצירת DNA. לרכיב ה- RNA של הטלומראז האנושי, הנקרא hTERC, יש רצף המשלים לחזרת הטלומרים, TAGGG. כפי שניתן לראות באיור 7.31, RNA זה יכול להתאים בסיס עם החזרה האחרונה של הטלומרים על גדיל ה-DNA ההורי, בעוד שחלק מה-RNA נשאר ללא זיווג.
תבנית עבור הרחבה
תפקידו של האזור הלא מזווג של ה-RNA הוא לשמש כתבנית שניתן להשתמש בה כדי להרחיב את קצה ה-3' התלוי של מולקולת ה-DNA המקורית. למרכיב החלבון של הטלומראז יש פעילות טרנסקריפטאז הפוכה והוא יכול להעתיק את רצף ה- RNA ל- DNA. בטלומראז אנושי, מרכיב החלבון ידוע בשם hTERT (telomerase reverse transcriptase). כפי שניתן לראות באיור 7.31 ו- 7.32, הטרנסקריפטאז ההפוך מרחיב את הסככה המקורית של 3 'באמצעות רכיב ה- RNA כתבנית שלו. לאחר מכן הטלומראז יכול להתנתק ולחזור על התהליך מספר פעמים כדי להוסיף חזרות רבות של רצף הטלומרים.
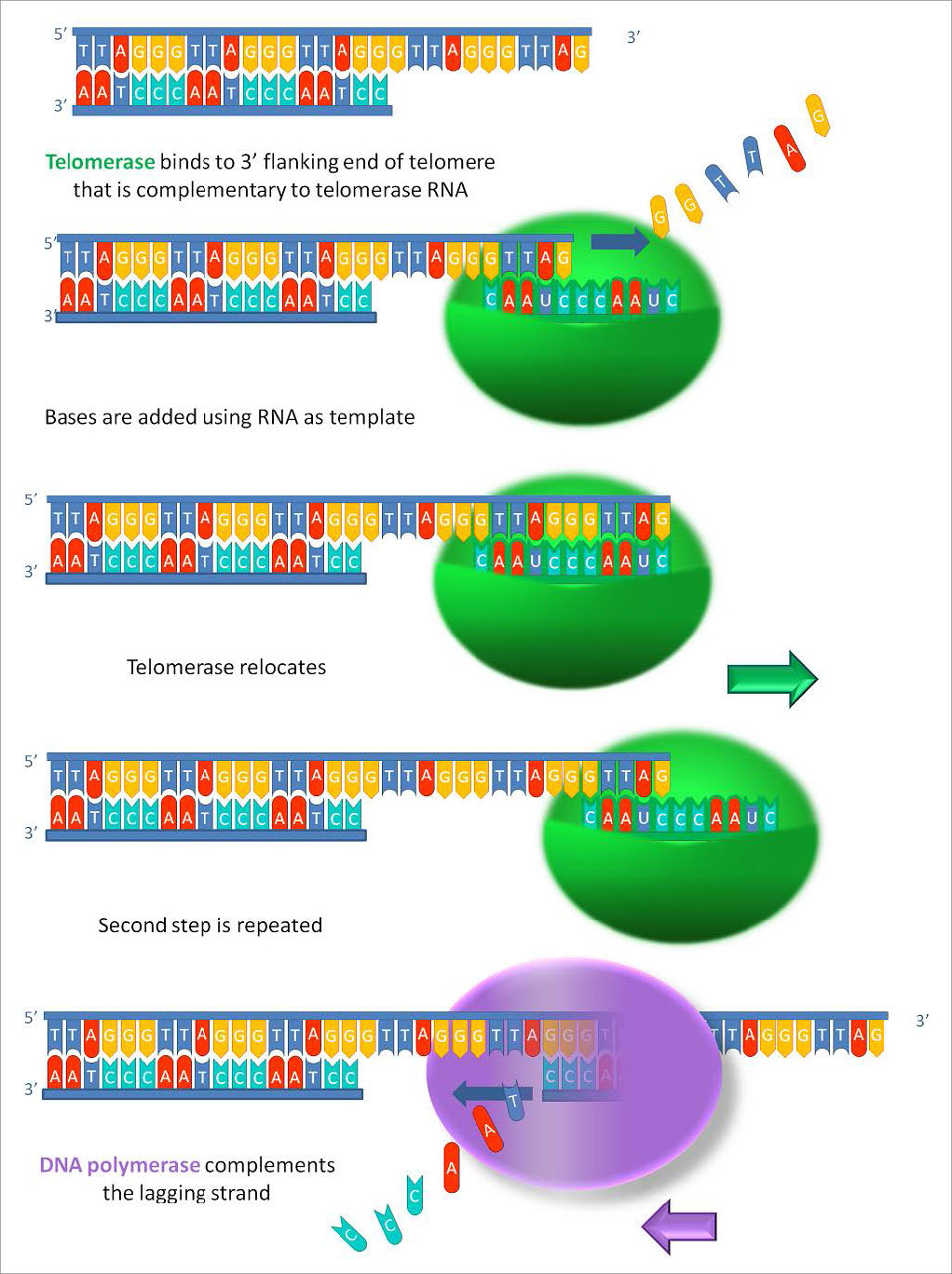
לאחר שהתליה הורחבה על ידי הוספת לפחות מספר חזרות של טלומרים, יש כעת מקום לסינתזה של פריימר RNA המשלים לתלייה המורחבת החדשה (מצביע לאחור לכיוון שאר הכרומוזום). לאחר מכן ניתן להרחיב את הפריימר הזה לסינתזה מלאה של הגדיל המפגר עד לקצה גדיל ה-DNA ההורי המקורי. לפיכך, תוספת של חזרות טלומרים על גדילי ה-DNA ההוריים מונעת מגדילי ה-DNA החדשים שנוצרו להתקצר עם כל מחזור שכפול. העובדה שזה קורה בתאי נבט (תאי רבייה) מסבירה מדוע לכל דור אין כרומוזומים קצרים יותר מדור ההורים.
פונקציית ההגהה של פולימראזות DNA עוקבת אחר הדיוק של שכפול ה-DNA בעוד שהאנזים טלומראז שומר על כרומוזומים שיועברו לצאצאים מהתקצרות. ביניהן שתי פעילויות אלה מבטיחות כי המידע הגנטי מועתק במדויק, וכי הדורות הבאים יקבלו השלמה מלאה של המידע הגנטי
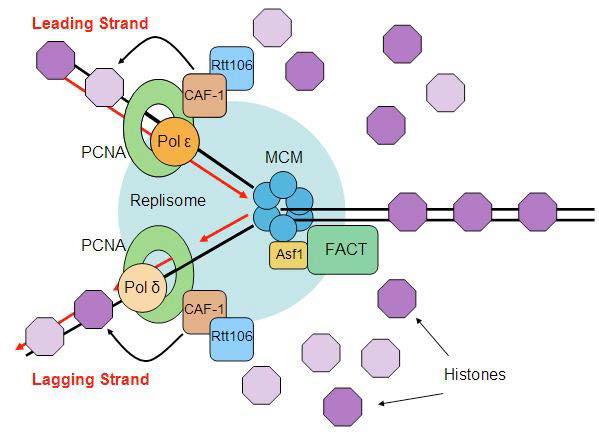
פירוק והרכבה מחדש של נוקלאוזומים
לאירועי השכפול יש טוויסט נוסף באיקריוטים. נזכיר כי DNA נמצא בתאים אוקריוטיים ככרומטין, קומפלקס של ה- DNA עם חלבונים. הכרומטין המעובה לפחות נראה כמו מחרוזת חרוזים, המורכבת מה- DNA העטוף סביב ליבות היסטון ליצירת נוקלאוזומים. יש לשבש את מבנה הנוקלאוזום כדי להפוך את ה-DNA לזמין לשכפול ולשחזר אותו לאחר השלמת השכפול (איור 7.33).
לפני מזלג השכפול, מבנה הכרומטין מפורק על ידי מתחמי שיפוץ כרומטין תלויי ATP, המאפשרים גישה לתבנית ה-DNA. לאחר סינתזה של גדילי ה- DNA החדשים, יש להרכיב מחדש את הנוקלאוזומים המקוריים וגם את הנוקלאוזומים החדשים מאחורי מזלג השכפול. מכיוון ששכפול מוליד שתי מולקולות DNA בהן הייתה אחת, יש צורך בכמות כפולה של היסטונים כדי לארוז את ה- DNA. ההכנה לשכפול DNA, אם כן, כוללת סינתזה של כמויות גדולות של היסטונים כדי לספק את הצורך. מעניין לציין כי DNA שסונתז לאחרונה ארוז לתוך נוקלאוזומים באמצעות ההיסטונים המקוריים שנעקרו כדי לאפשר למזלג השכפול לעבור, כמו גם היסטונים שסונתזו לאחרונה.
אנו גם יודעים ששינויים שלאחר תרגום כמו אצטילציה, מתילציה או זרחון של ההיסטונים יכולים לווסת את המידה שבה אזור נתון בגנום נגיש לשימוש. שאלה אחת שנותרה נושא למחקר אינטנסיבי היא כיצד שינויים אלה מועברים במדויק לנוקלאוזומים החדשים.