
5.3: אנרגיה - פוטופוספורילציה
- Page ID
- 207614

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)מקור: BiochemFFA_5_3.pdf. ספר הלימוד כולו זמין בחינם מהמחברים בכתובת http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy

פוטופוספורילציה
הסוג השלישי של זרחון לייצור ATP נמצא רק בתאים המבצעים פוטוסינתזה. תהליך זה דומה לזרחון חמצוני בכמה אופנים. ההבדל העיקרי הוא המקור האולטימטיבי לאנרגיה לסינתזת ATP. בזרחן חמצוני האנרגיה מגיעה מאלקטרונים המיוצרים על ידי חמצון של מולקולות ביולוגיות. בפוטוסינתזה האנרגיה מגיעה מאור השמש. פוטונים מהשמש מקיימים אינטראקציה עם מולקולות כלורופיל במרכזי תגובה בכלורופלסטים (דמויות \(\PageIndex{1}\) ו\(\PageIndex{2}\)) של צמחים או ממברנות של חיידקים פוטוסינתטיים.
קווי הדמיון של פוטופוספורילציה לזרחון חמצוני כוללים:
- שרשרת הובלת אלקטרונים הקשורה לממברנה
- יצירת שיפוע פרוטון
- קצירת אנרגיה של שיפוע הפרוטונים על ידי יצירת ATP בעזרת סינתאז ATP.
חלק מההבדלים כוללים:
- מקור האלקטרונים — H2O לפוטוסינתזה לעומת NADH/FADH2 עבור זרחון חמצוני
- כיוון שאיבת פרוטונים - לחלל התילקואיד של הכלורופלסטים לעומת מחוץ למטריצה של המיטוכונדריון
- תנועה של פרוטונים במהלך סינתזת ATP - מחוץ לחלל התילקואיד בפוטוסינתזה לעומת לתוך המטריצה המיטוכונדריאלית בזרחן חמצוני
- אופיו של מקבל האלקטרונים הסופי - NADP+ בפוטוסינתזה לעומת O2 בזרחון חמצוני.
הובלת אלקטרונים: כלורופלסטים לעומת מיטוכונדריה
במובנים מסוימים, תנועת האלקטרונים בכלורופלסטים במהלך הפוטוסינתזה מנוגדת לזו של הובלת אלקטרונים במיטוכונדריה. בפוטוסינתזה, מים הם מקור האלקטרונים והיעד הסופי שלהם הוא NADP+ לייצור NADPH. במיטוכונדריה, NADH/FADH2 הם מקורות אלקטרונים ו- H2O הוא היעד הסופי שלהם. כיצד מערכות ביולוגיות גורמות לאלקטרונים ללכת לשני הכיוונים? נראה שזה המקבילה ללכת למקום מסוים וממנו תוך ירידה תמיד, מכיוון שאלקטרונים ינועו בהתאם לפוטנציאל.
אנרגיה סולארית
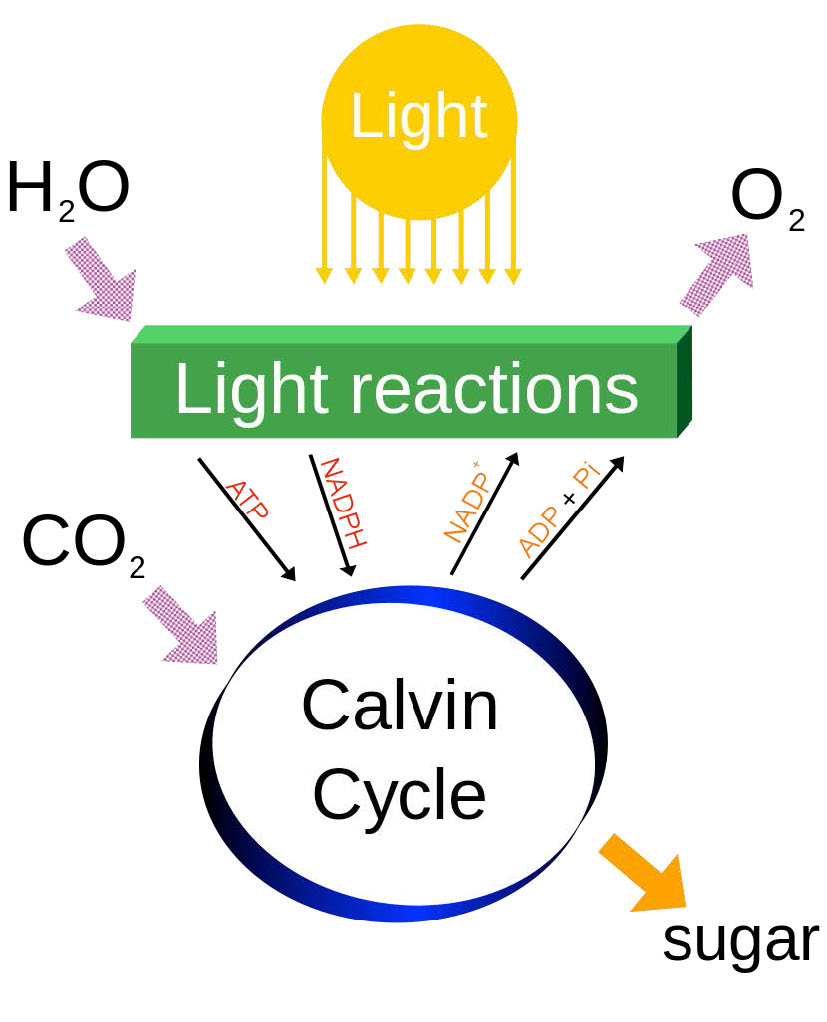
התשובה היא האנרגיה שנלכדה של הפוטונים מהשמש (איור 5.59), שמעלה אלקטרונים לאנרגיה שבה הם נעים "בירידה" ליעד ה- NADPH שלהם בתכנית בצורת Z. תנועת האלקטרונים באמצעות תכנית זו בצמחים דורשת אנרגיה מפוטונים בשני מקומות כדי "להרים" את האנרגיה של האלקטרונים מספיק.
לבסוף, יש לציין כי לפוטוסינתזה יש למעשה שני שלבים, המכונים מחזור האור (שתואר לעיל) והמחזור האפל, שהוא קבוצה של תגובות כימיות הלוכדת CO2 מהאטמוספירה ו"מתקן "אותו, בסופו של דבר לגלוקוז. המחזור האפל מכונה גם מחזור קלווין ונדון כאן.
פוטוסינתזה
פוטוסינתזה היא תהליך לכידת אנרגיה המצוי בצמחים ובאורגניזמים אחרים כדי לקצור אנרגיית אור ולהמיר אותה לאנרגיה כימית. אנרגיה פוטוכימית זו מאוחסנת בסופו של דבר בפחמימות המיוצרות באמצעות ATP (מקצירת האנרגיה), פחמן דו חמצני ומים. ברוב המקרים, תוצר לוואי של התהליך הוא חמצן, המשתחרר מהמים בתהליך הלכידה. הפוטוסינתזה אחראית לרוב החמצן באטמוספירה והיא מספקת את החומרים האורגניים ואת רוב האנרגיה המשמשת את החיים על פני כדור הארץ.
צעדים
השלבים בתהליך הפוטוסינתזה משתנים מעט בין אורגניזמים. בסקירה רחבה, זה תמיד מתחיל בלכידת אנרגיה מאור על ידי מתחמי חלבון, המכילים פיגמנטים של כלורופיל, הנקראים מרכזי תגובה. צמחים חוטפים חלבונים אלה בכלורופלסטים, אך חיידקים, שאין להם אברונים, מטמיעים אותם בממברנות הפלזמה שלהם.
אנרגיה מהאור משמשת להפשטת אלקטרונים מתורמי אלקטרונים (בדרך כלל מים) ולהשאיר תוצר לוואי (חמצן, אם נעשה שימוש במים). אלקטרונים נתרמים לנשא ובסופו של דבר מתקבלים על ידי NADP+, כדי להפוך ל- NADPH. כאשר אלקטרונים נעים לכיוון NADP+, הם יוצרים שיפוע פרוטונים על פני הממברנה התילקואידית, המשמשת להנעת סינתזה של ATP. לפיכך NADPH, ATP וחמצן הם תוצרי השלב הראשון של הפוטוסינתזה הנקרא תגובות האור. אנרגיה מ-ATP ואלקטרונים מ-NADPH משמשים להפחתת CO2 ולבניית סוכרים, שהם אחסון האנרגיה האולטימטיבי הנובע ישירות מפוטוסינתזה.
כלורופלסטים
כלורופלסטים נמצאים כמעט בכל תאי הצמח מעל הקרקע, אך מרוכזים בעיקר בעלים. פנים העלה, מתחת לאפידרמיס מורכב מרקמת פוטוסינתזה הנקראת מזופיל, שיכולה להכיל עד 800,000 כלורופלסטים למילימטר רבוע.
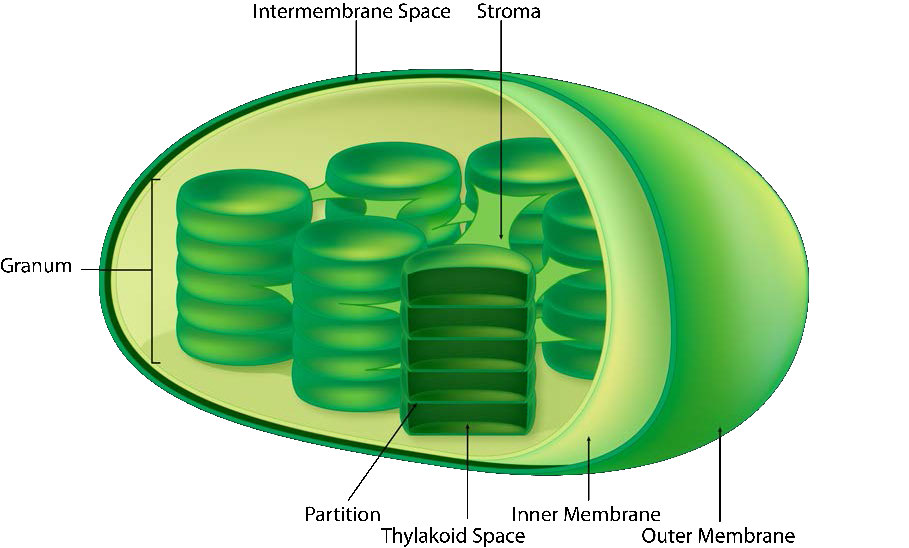
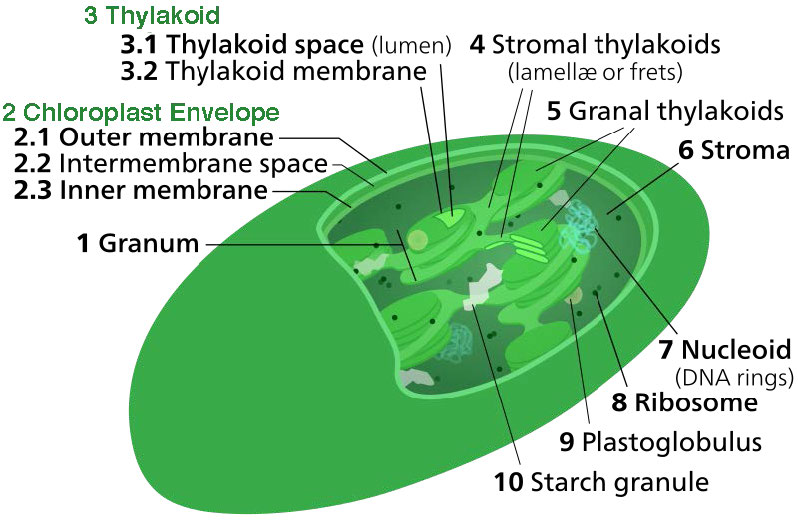
לקרום הכלורופלסט יש קרום פנימי פוספוליפיד, קרום חיצוני של פוספוליפיד ואזור ביניהם הנקרא החלל הבין-ממברני (איור 5.61). בתוך קרום הכלורופלסט הפנימי נמצאת הסטרומה, בה נמצאים ה- DNA של הכלורופלסט והאנזימים של מחזור קלווין. כמו כן בתוך הסטרומה מוערמים דיסקים שטוחים המכונים תילקואידים המוגדרים על ידי ממברנות התילקואיד שלהם. החלל בתוך ממברנות התילקואיד נקרא מרחבי התילקואיד או לומן התילקואיד. מתחמי החלבון המכילים את הפיגמנטים הסופגים אור, המכונים מערכות צילום, ממוקמים על הממברנה התילקואידית. מלבד כלורופילים, קיימים גם קרוטנים וקסנטופילים המאפשרים ספיגת אנרגיית אור בטווח רחב יותר. אותם פיגמנטים משמשים אצות ירוקות וצמחי אדמה.
אצות חומות ודיאטומים מוסיפים פוקוקסנטין (קסנטופיל) ואצות אדומות מוסיפות פיקואריתרין לתערובת. בצמחים ובאצות, הפיגמנטים מוחזקים בצורה מאורגנת מאוד קומפלקסים הנקראים חלבוני אנטנה המסייעים בהעברת אנרגיה, באמצעות העברת אנרגיית תהודה, לכלורופילים של מרכז התגובה. מערכת מאורגנת כל כך נקראת קומפלקס קציר קל. מתחמי הובלת האלקטרונים של הפוטוסינתזה ממוקמים גם על ממברנות התילקואיד.
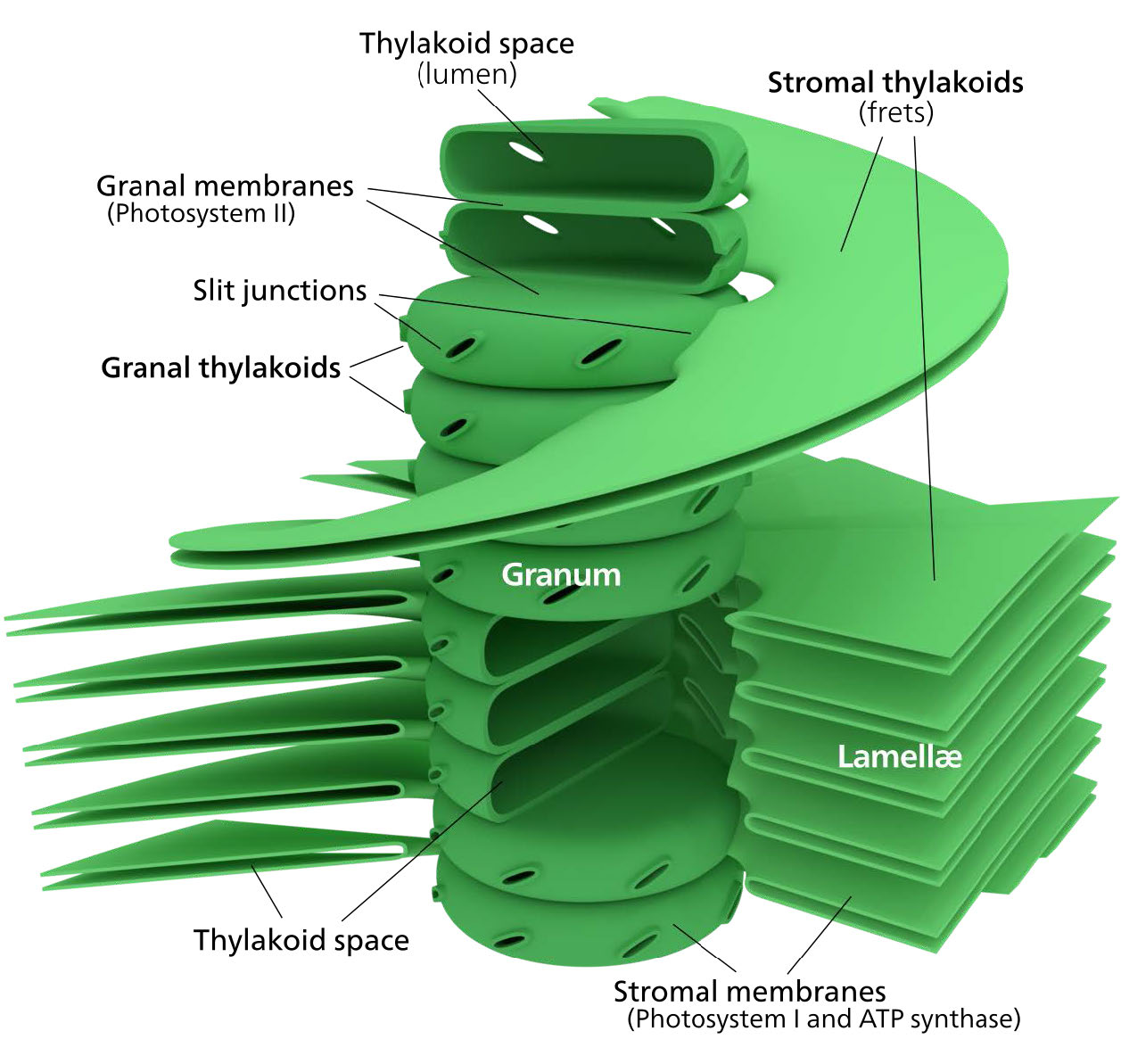
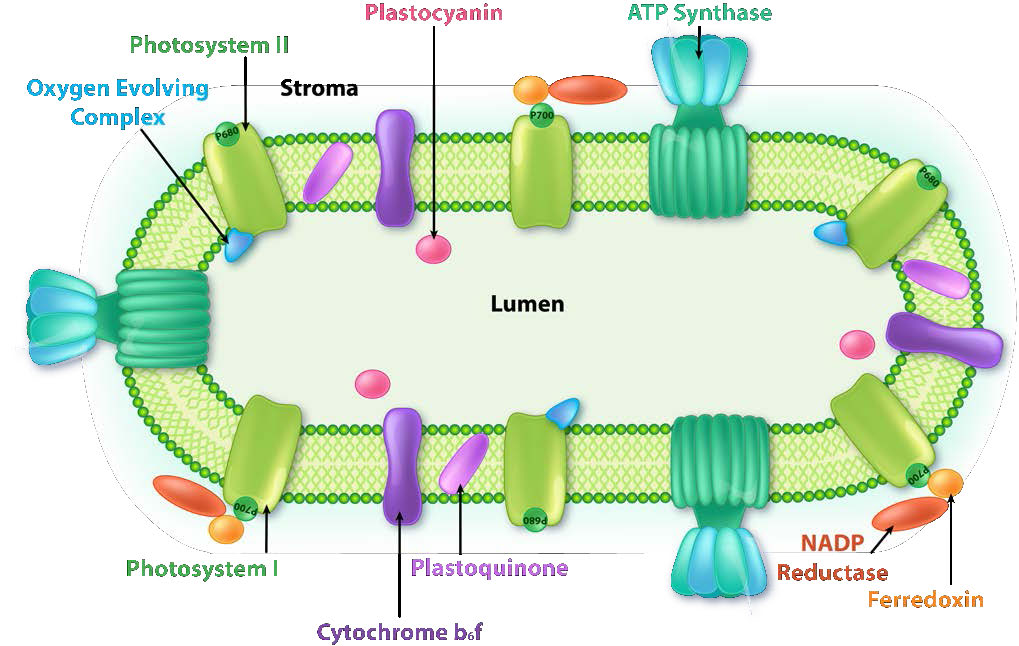
איור \(\PageIndex{6}\): קומפלקסים בקרום התילקואיד. תמונה על ידי אליה קים
תגובות אור של פוטוסינתזה
בכלורופלסטים, תגובות האור של הפוטוסינתזה הכרוכות בהעברת אלקטרונים מתרחשות בממברנות התילקואיד (איור). \(\PageIndex{6}\) תגובות ביוכימיות נפרדות הכוללות הטמעה של פחמן דו חמצני לייצור גלוקוז מכונות מחזור קלווין, המכונה לפעמים גם "התגובות האפלות". זה יידון במקומות אחרים בסעיף על מטבוליזם (כאן).
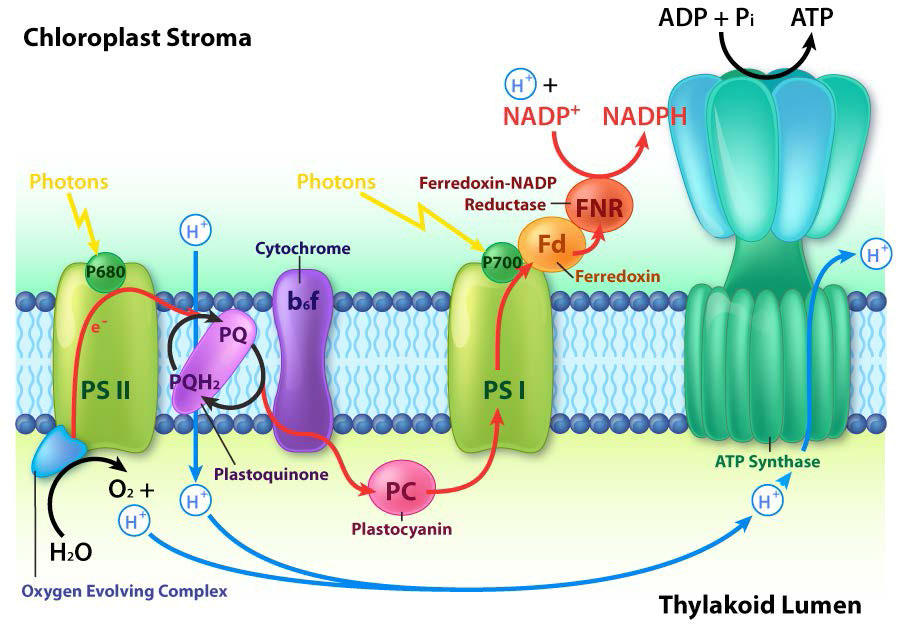
הכלורופלסטים הם המקום בו נלכדת אנרגיית האור, אלקטרונים מופשטים מהמים, חמצן משתחרר, מתרחשת הובלת אלקטרונים, נוצר NADPH ונוצר ATP. הממברנה התילקואידית תואמת את הממברנה הפנימית של המיטוכונדריון להובלת אלקטרונים ושאיבת פרוטונים (איור). \(\PageIndex{4}\)
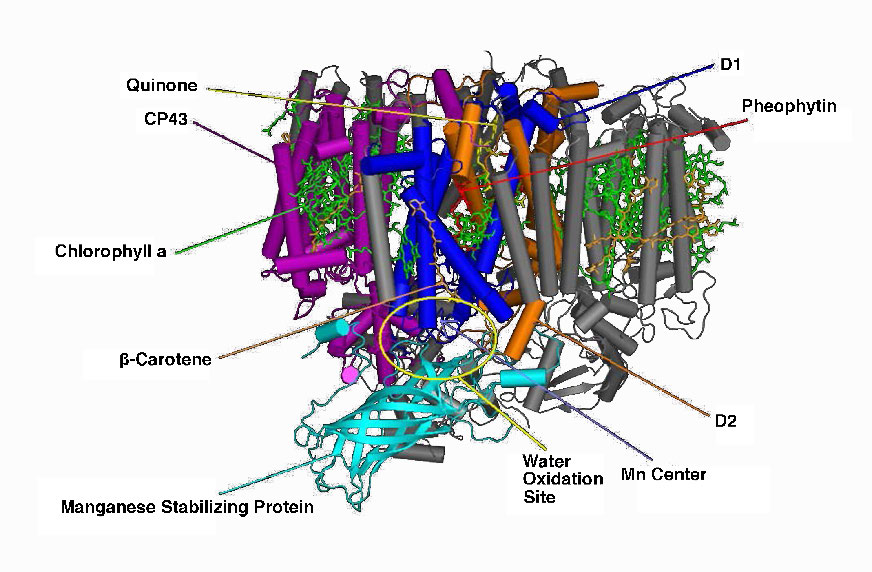
קרום התילקואיד עושה את הקסם שלו באמצעות ארבעה מתחמי חלבון עיקריים. אלה כוללים מערכת צילום II (PS II), קומפלקס ציטוכרום b6f (CB6f), מערכת צילום I (PS I) וסינתאז ATP. התפקידים של קומפלקסים אלה, בהתאמה, הם ללכוד אנרגיית אור, ליצור שיפוע פרוטונים מתנועת אלקטרונים, ללכוד אנרגיית אור (שוב) ולהשתמש באנרגיית שיפוע פרוטונים מהתהליך הכולל כדי לסנתז ATP.
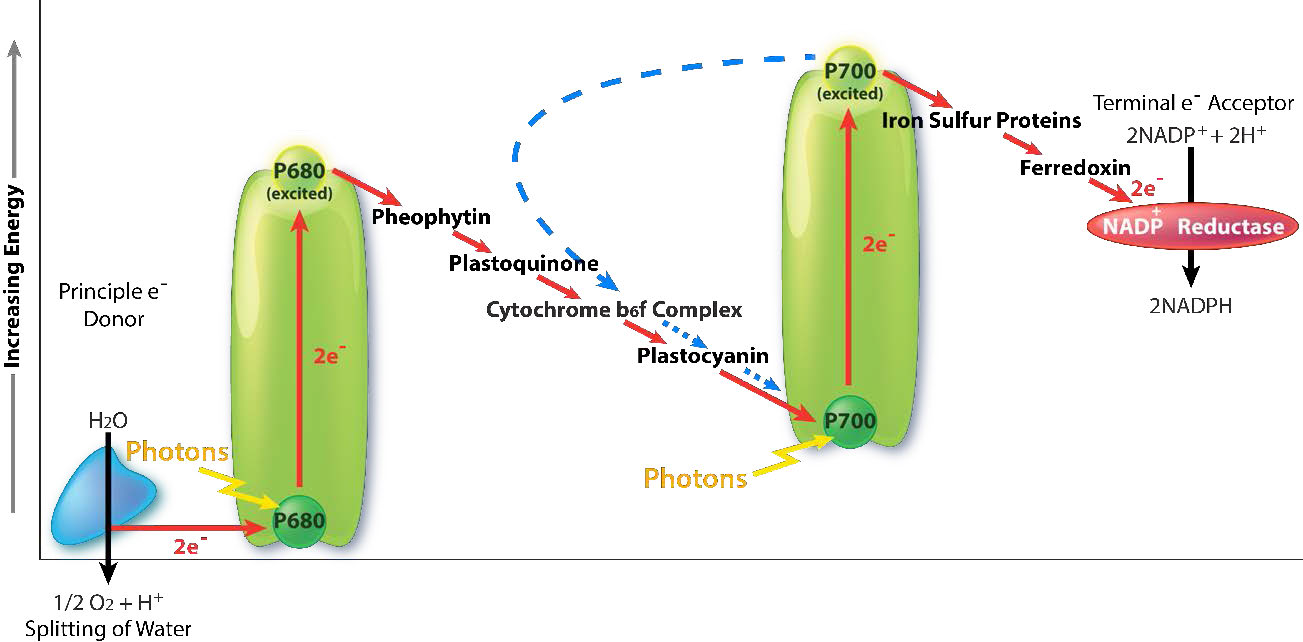
קציר קל
קצירת אנרגיית האור מתחילה ב- PS II עם ספיגת פוטון אור במרכז תגובה. PS II מבצע את החובה הזו בצורה הטובה ביותר עם אור באורך גל של 680 ננומטר והוא מאבד בקלות אלקטרון לעירור כאשר זה קורה, ומשאיר את PS II עם מטען חיובי. יש להחליף את האלקטרון הזה. המקור החלופי האולטימטיבי לאלקטרונים הוא מים, אך מים חייבים לאבד ארבעה אלקטרונים ו- PS II יכול לקבל רק אחד בכל פעם.
מרכזי מנגן
קומפלקס ביניים של חמצן מתפתח (OEC) מכיל ארבעה מרכזי מנגן המספקים את האלקטרון החלופי המיידי ש-PSII דורש. לאחר שארבעה אלקטרונים נתרמו על ידי ה- OEC ל- PS II, ה- OEC מחלץ ארבעה אלקטרונים משתי מולקולות מים, משחרר חמצן ומשליך ארבעה פרוטונים לחלל התילקואיד, ובכך תורם לשיפוע הפרוטונים. יש להעביר את האלקטרון הנרגש מ- PS II לנשא אחר במהירות רבה, שמא יתפורר למצבו המקורי. הוא עושה זאת, נותן את האלקטרון שלו בתוך פיקו-שניות לפאופיטין (איור). \(\PageIndex{8}\)
פיופיטין מעביר את האלקטרון לפלסטוקינונים הקשורים לחלבון. הראשון ידוע בשם PQA. PQA מעביר את האלקטרון לפלסטוקינון שני (PQB), שמחכה לאלקטרון שני ואוסף שני פרוטונים שיהפכו ל-PQH2, הידוע גם בשם פלסטוקינול (איור). \(\PageIndex{9}\) PQH2 מעביר אותם לקומפלקס ציטוכרום b6f (CB6f) המשתמש במעבר אלקטרונים דרכו כדי לשאוב פרוטונים לחלל התילקואיד. סינתאז ATP מייצר ATP משיפוע הפרוטונים שנוצר בדרך זו. CB6f מוריד את האלקטרון בפלסטוציאנין, המחזיק אותו עד שתהליך העירור הבא מתחיל בספיגת פוטון אור נוסף ב-700 ננומטר על ידי PS I.
קליטת אור ב- PS I
עם קליטת פוטון אור על ידי PS I, מתחיל תהליך הדומה לתהליך ב- PS II. PS I צובר מטען חיובי כתוצאה מאובדן אלקטרון נרגש ומושך ממנו את האלקטרון בפלסטוציאנין. בינתיים, האלקטרון הנרגש מ-PS I עובר דרך חלבון ברזל-גופרית, שנותן את האלקטרון לפרדוקסין (עוד חלבון גופרית ברזל). לאחר מכן פרדוקסין מעביר את האלקטרון לחלבון האחרון במערכת המכונה פרדוקסין:NADP+ אוקסידורדוקטאז, המעניק את האלקטרון והפרוטון ל- NADP+, ויוצר NADPH.
שים לב שהפחתת NADP+ ל-NADPH דורשת שני אלקטרונים ופרוטון אחד, כך שארבעת האלקטרונים ושני הפרוטונים מחמצון מים יביאו לייצור של שתי מולקולות של NADPH. בשלב זה מחזור האור הושלם - מים התחמצנו, נוצר ATP ונוצר NADPH. האלקטרונים עשו את דרכם ממים ל- NADPH באמצעות נשאים בקרום התילקואיד ותנועתם שחררה מספיק אנרגיה לייצור ATP. אנרגיה לכל התהליך הגיעה מארבעה פוטונים של אור.
שתי מערכות הצילום המבצעות את כל הקסם הזה הן מתחמי חלבון הדומים במבנה ובאמצעי הפעולה. הם סופגים פוטונים ביעילות גבוהה כך שבכל פעם שפיגמנט במרכז התגובה הפוטוסינתטית סופג פוטון, אלקטרון מהפיגמנט מתרגש ומועבר למולקולה אחרת כמעט באופן מיידי. תגובה זו נקראת הפרדת מטען המושרה על ידי צילום והיא אמצעי ייחודי להפיכת אנרגיית האור לצורות כימיות.
פוטופוספורילציה מחזורית
מלבד הנתיב שתואר לעיל לתנועת אלקטרונים דרך PS I, לצמחים יש מסלול חלופי שאלקטרונים יכולים לעבור. במקום שאלקטרונים יעברו דרך פרדוקסין ליצירת NADPH, הם במקום זאת עוברים נתיב לאחור דרך קומפלקס b6f השואב פרוטונים. מערכת זו, הנקראת פוטופוספורילציה מחזורית (איור\(\PageIndex{8}\)) המייצרת יותר ATP וללא NADPH, דומה למערכת שנמצאת בחיידקי גופרית ירוקה. היכולת של צמחים לעבור בין מערכות צילום לא מחזוריות ומחזוריות מאפשרת להם ליצור את היחס הנכון בין ATP ל-NADPH שהם צריכים להטמעת פחמן בשלב האפל של הפוטוסינתזה. יחס זה מתברר כ-3 ATP ל-2 NADPHs.
איור \(\PageIndex{9}\) - מערכת צילום II של ציאנובקטריה. ויקיפדיה
אנרגיה פוטוסינתטית
הפלט של חלק הפוטופוספורילציה של הפוטוסינתזה (O2, NADPH ו-ATP), כמובן, אינו סוף תהליך הפוטוסינתזה. עבור הצמח הגדל, ה-NADPH וה-ATP משמשים ללכידת פחמן דו חמצני מהאטמוספירה ולהמיר אותו (בסופו של דבר) לגלוקוז ותרכובות פחמן חשובות אחרות. זה, כפי שצוין קודם לכן, מתרחש במחזור קלווין (ראה כאן) במה שמכונה השלב האפל של התהליך. החמצן המשתחרר בתהליך הוא הכרחי לנשימה של כל צורות החיים האירוביות על פני כדור הארץ. ואכן, מאמינים שבעצם כל החמצן באטמוספירה כיום הוא תוצאה של פיצול מים בפוטוסינתזה לאורך עידנים רבים שהתהליך התקיים.