
3.3: שיקולים אחרים בממברנות
- Page ID
- 207679

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)מקור: BiochemFFA_3_3.pdf. ספר הלימוד כולו זמין בחינם מהמחברים בכתובת http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy
ישנם פונקציות וגורמים רבים הקשורים לממברנות התא שאינן מתאימות לקטגוריות רחבות. פריטים אלה יהיו המוקד של סעיף זה.
אנדוציטוזיס
מלבד חלבוני טרנספורטר ותעלות יונים, דרך נפוצה נוספת לחומרים להיכנס לתאים היא על ידי תהליך האנדוציטוזיס. אנדוציטוזיס היא צורה חלופית של הובלה פעילה להכנסת חומרים לתאים. חלק מהתהליכים הללו, כגון פגוציטוזיס, מסוגלים לייבא חלקיקים גדולים בהרבה ממה שהיה אפשרי באמצעות חלבון טרנספורטר. בדומה לחלבוני טרנספורטר, אנדוציטוזיס משתמשת באנרגיה למטרה (אם כי היא אינה נראית כמו אצל מובילי חלבון), אך בניגוד למובילי חלבון, התהליך אינו ספציפי כמעט למולקולות בודדות.
כתוצאה מכך, התהליך כרוך בדרך כלל בייבוא של מולקולות רבות ושונות בכל פעם שהוא מתרחש. רשימת התרכובות הנכנסות לתאים בדרך זו כוללת LDL ותכולת השומנים שלהם, אך היא כוללת גם דברים כמו ברזל (ארוז בטרנספרין), ויטמינים, הורמונים, חלבונים ואפילו כמה וירוסים מתגנבים באמצעים אלה. ישנם שלושה סוגים של אנדוציטוזיס שנשקול (איור 3.53).
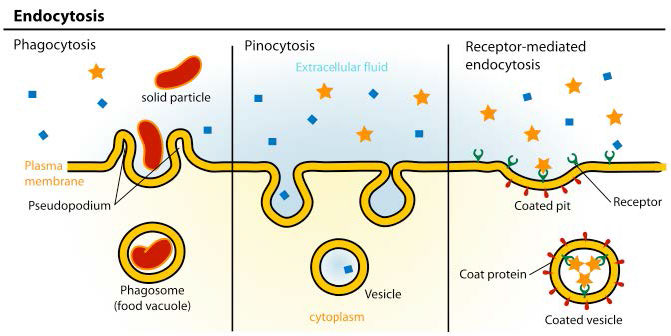
אנדוציטוזה בתיווך קולטן
תהליך האנדוציטוזה בתיווך קולטן הוא אמצעי ספציפי יחסית להכנסת מולקולות לתאים מכיוון שהוא דורש שהחומר הנכנס יהיה קשור איכשהו לקולטן ספציפי של פני התא. בדוגמה של איור 3.53, הקולטן הוא קולטן ה-LDL הסלולרי. פלישות מצופות קלתרין, כפי שמוצג באיור ידועות בשם "בורות מצופים". המנגנון ממשיך עם ניצנים פנימיים של קולטן קרום הפלזמה (שלפוחיות מצופות). קישור הליגנד (ApoB-100 של ה-LDL, למשל, באיור 3.54) לקולטן ה-LDL מוביל להיווצרות של פלישת ממברנה. חלקיקי ה-LDL הנספגים מתמזגים ליצירת אנדוזום מוקדם (איור 3.55) והתוכן ממוינים ומעובדים לאחר מכן לשימוש התא.
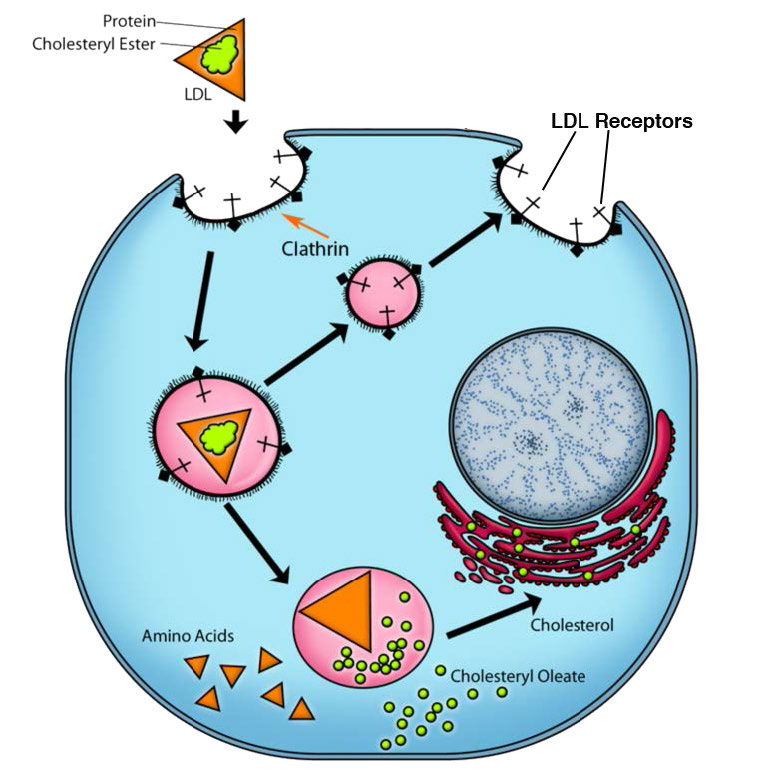
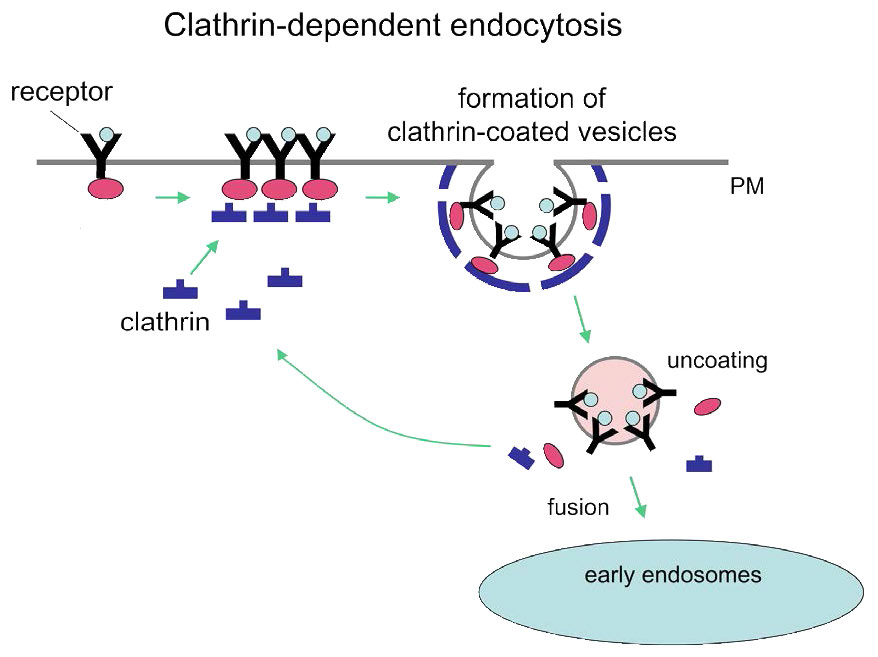
הרכיבים מהשלפוחית המצופה ממוחזרים לקרום הפלזמה לפעולות נוספות. אנדוציטוזה בתיווך קולטן יכולה גם למלא תפקיד בהפנמה של קולטנים תאיים המתפקדים בתהליך האיתות. כאן, קולטן הקשור לליגנד מובא לתא ועשוי בסופו של דבר ליצור תגובה בגרעין.
בעוד שאנדוציטוזה בתיווך קולטן של קולטנים יכולה להשפיע על העברת אות פנימה לתא, היא יכולה גם להפחית את כמות האיתות הכוללת המתרחשת, מכיוון שמספר הקולטנים על פני התא יורד בתהליך.
אנדוציטוזה שאינה קלתרין
ישנם שלושה סוגים של אנדוציטוזיס המתרחשים בתאים שאינם כוללים קלתרין. הם 1) אנדוציטוזה מבוססת מערות, 2) מקרופינוציטוזיס ו-3) פגוציטוזיס. אנדוציטוזה מבוססת Caveolae מבוססת על מולקולת קולטן המכונה caveolin. Caveolins הם מחלקה של חלבוני ממברנה אינטגרליים המחלקים ומרכזים מולקולות איתות בתהליך האנדוציטוזיס. הם נקראים על שם מבני המערות דמויי המערות של קרום הפלזמה שבו הם נמצאים.
קאבאולינים
לקאולינים יש זיקה לכולסטרול ומתחברים אליו בקרום התאים, וגורמים להיווצרות פלישות ממברנה של כ- 50 ננומטר. חלבוני הקאולין יכולים לבצע אוליגומריזציה וזה חשוב לציפוי ויצירת המבנים דמויי המערה.
ישנם שלושה גנים של קאולין המצויים בתאי חוליות, CAV1, CAV2 ו- CAV3. ויסות מטה של caveolin-1 מביא להגירה סלולרית פחות יעילה במבחנה. Caveolins מעורבים הן בהיווצרות והן בדיכוי של גידולים. ביטוי גבוה שלהם מעכב מסלולי איתות של גורמי גדילה הקשורים לסרטן, אך חלק מתאי הסרטן המבטאים קייבולין הם אגרסיביים וגרורתיים יותר, אפשריים בשל יכולת משופרת לצמיחה בלתי תלויה בעיגון.
מקרופינוציטוזיס
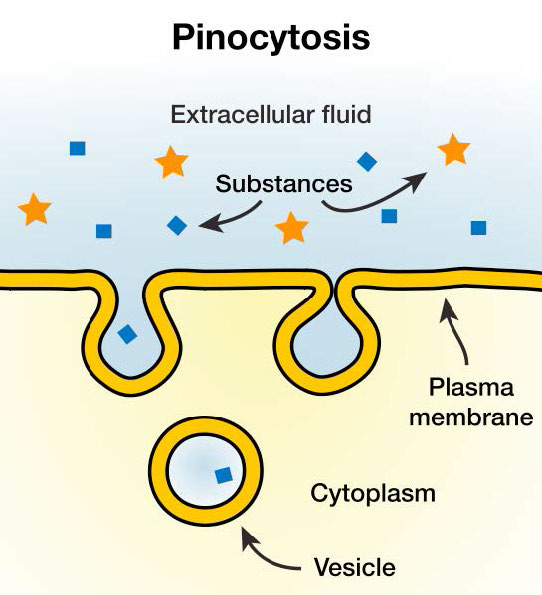
תופעה המכונה "שתיית תאים", מקרופינוציטוזיס כוללת פשוטו כמשמעו תא "לוקח לגימה" של הנוזל החוץ תאי. הוא עושה זאת, כפי שמוצג באיור 3.56, על ידי פלישה פשוטה של תכונות פני השטח הפרועות של קרום הפלזמה. נוצר כיס, שצובט פנימה ליצירת שלפוחית המכילה נוזל חוץ תאי ומולקולות מומסות. בתוך הציטוזול, שלפוחית מופנמת זו תתמזג עם אנדוזומים וליזוזומים. התהליך אינו ספציפי לחומרים שהופנמו.
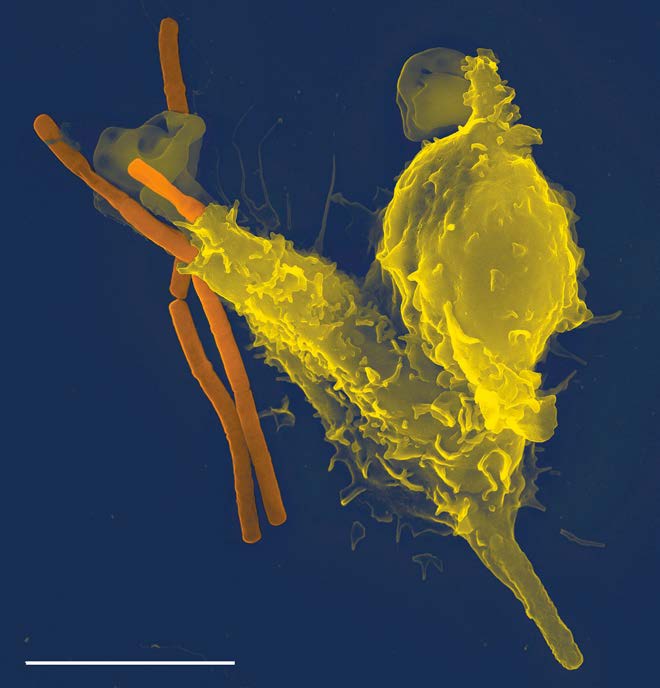
פגוציטוזיס
פגוציטוזיס הוא תהליך שבו מכוונים חלקיקים גדולים יחסית (קוטר 0.75 מיקרומטר). תאים של מערכת החיסון, כגון נויטרופילים, מקרופאגים ואחרים, משתמשים בפגוציטוזיס להפנמת פסולת תאים, תאים אפופטוטיים ומיקרואורגניזמים.
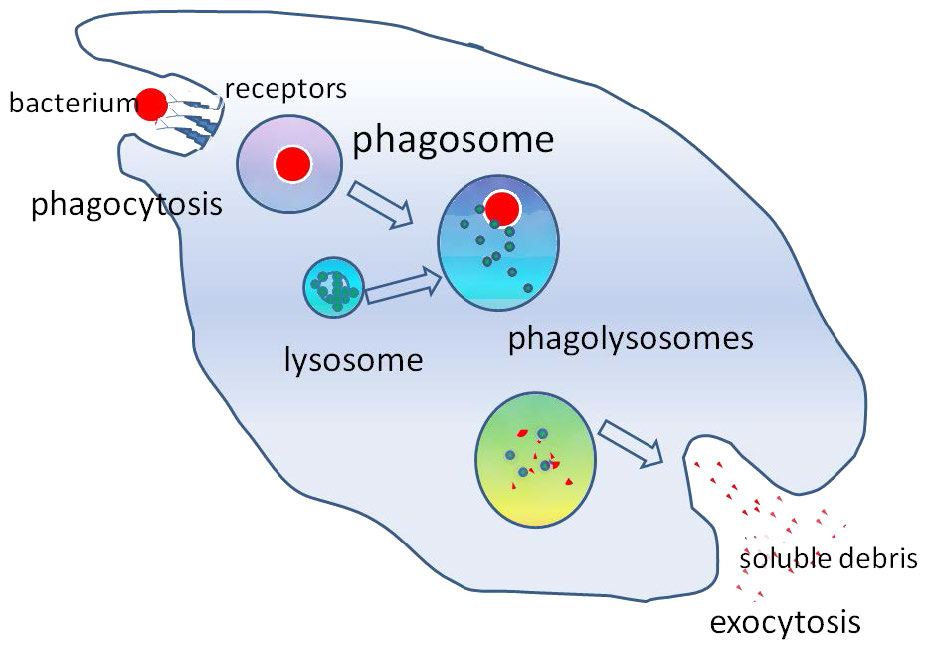
התהליך פועל באמצעות קולטנים ספציפיים על פני התא ותא פגוציטוזינג בולע את מטרתו על ידי גידול סביבו. המבנה המופנם ידוע כפגוזום, שמתמזג במהירות עם ליזוזום ליצירת פגוליזוזום (איור 3.58), אשר מכניס את החלקיק הנבלע לתנאים רעילים כדי להרוג אותו, אם הוא תא, ו/או לעכל אותו לחתיכות קטנות יותר. במקרים מסוימים, כפי שמוצג באיור, פסולת מסיסה עשויה להשתחרר על ידי התא הפגוציטוזינג.
אנדוזומים
חומר מופנם מאנדוציטוזיס שאינו כרוך בפגוציטוזיס עובר דרך מבנה מופנם הנקרא אנדוזום. אנדוזומים הם מבנים מוגבלים בממברנה בתוך תאים אוקריוטיים הממלאים תפקיד באנדוציטוזה (איור 3.59). יש להם פונקציית מיון של חומר שהופנם לתא, ומספקת אחזור של חומרים שאינם מיועדים להרס בליזוזומים. LDLs, למשל, ממוקדים לאחר אנדוציטוזה לאנדוזומים לעיבוד לפני שחלק מהם מועבר לליזוזום. האנדוזומים יכולים גם לקבל מולקולות מרשת הטרנס-גולגי. אלה יכולים להימסר גם לליזוזומים, או להפנות אותם חזרה לגולגי. אנדוזומים מגיעים בשלוש צורות - 1) מוקדם, 2) מאוחר ו -3) מיחזור.
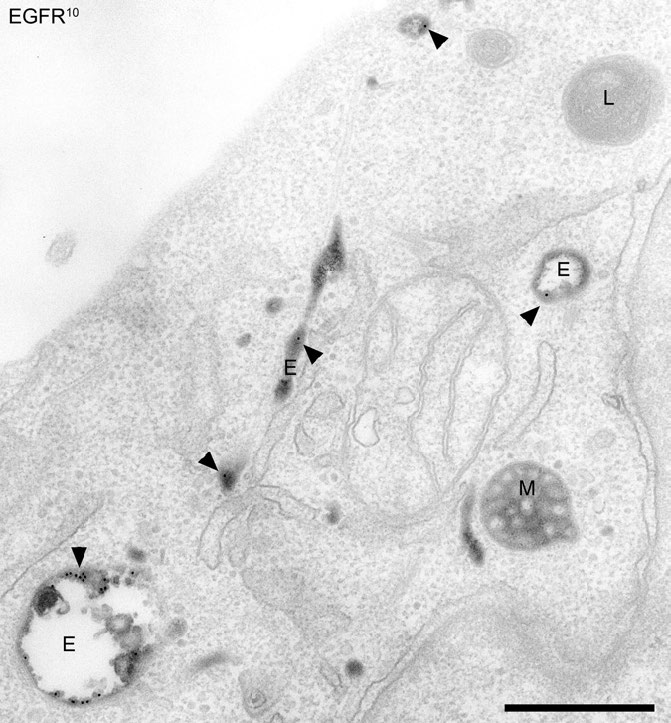
איור 3.59 - הפנמה של קולטן גורם הגדילה האפידרמיס (EGFR) לאנדוזומים. אנדוזומים וליזוזומים מוקדמים (E) ומאוחרים (M) מסומנים. - ויקיפדיה
אקסוציטוזיס
תהליך האקסוציטוזיס משמש את התאים לייצוא מולקולות מתוך תאים שאחרת לא היו עוברים בקלות דרך קרום הפלזמה. בתהליך, שלפוחיות הפרשה מתמזגות עם קרום הפלזמה ומשחררות את תוכנן מחוץ לתאים. חומרים, כגון חלבונים ושומנים המוטבעים בממברנות השלפוחיות הופכים לחלק מממברנת הפלזמה כאשר מתרחש מיזוג בינה לבין שלפוחית השתן.
היתוך ממברנה
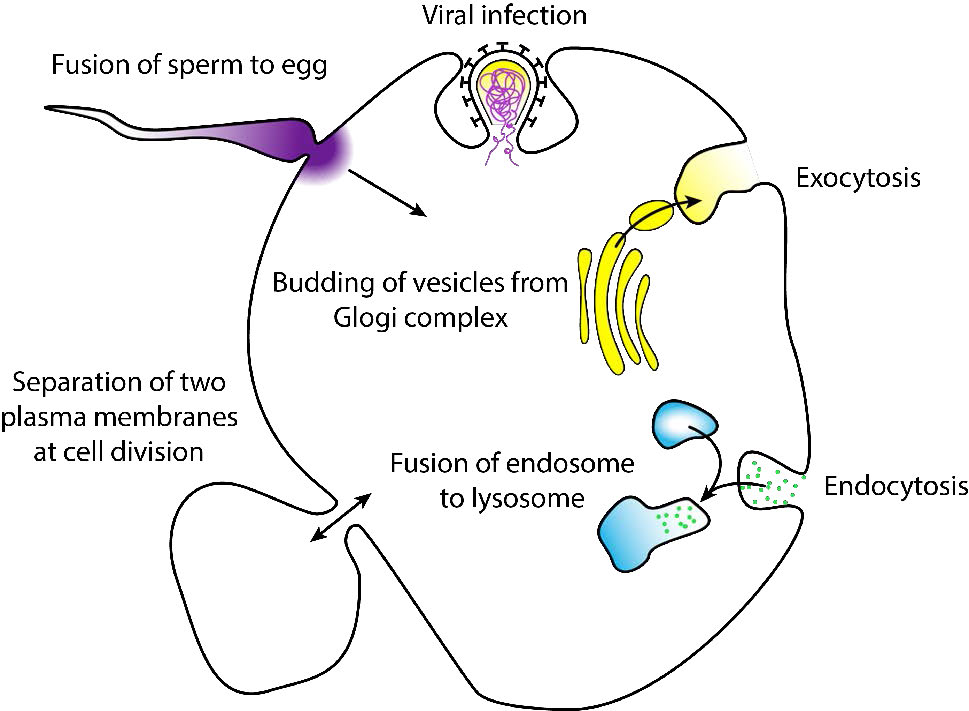
היתוך הוא תהליך ממברנה שבו שתי שכבות שומנים נפרדות ממזגות את הליבות ההידרופוביות שלהן, ומייצרות מבנה אחד מחובר זה לזה. היתוך ממברנה הכולל שלפוחיות הוא המנגנון שבאמצעותו מתרחשים תהליכי האנדוציטוזה והאקסוציטוזיס.
כאשר ההיתוך עובר דרך שני העלונים של שתי השכבות הדו-שכבות, נוצר גשר מימי ותכולת שני המבנים מתערבבת. תהליכים נפוצים הכוללים היתוך ממברנה (איור 3.60) כוללים הפריה של ביצית על ידי זרע, הפרדת ממברנות בחלוקת התא, הובלת מוצרי פסולת ושחרור נוירוטרנסמיטר (איור 3.61). ממברנות מלאכותיות כמו ליפוזומים יכולות להתמזג גם עם ממברנות תאיות. היתוך חשוב גם להובלת שומנים מנקודת הסינתזה בתוך התא לקרום שבו הם משמשים. כניסה של פתוגנים יכולה להיות נשלטת גם על ידי היתוך, שכן וירוסים רבים מצופים דו -שכבתי משתמשים בחלבוני היתוך בכניסה לתאי מארח.
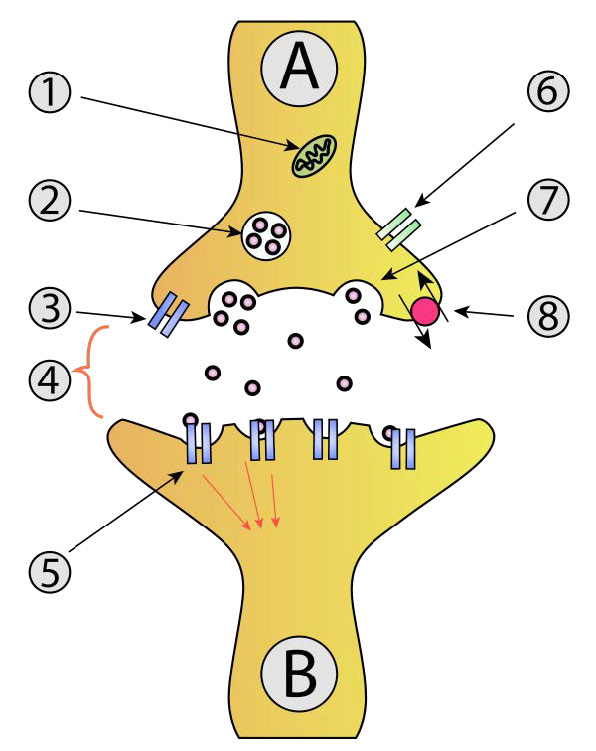
חלבוני מלכודת
תיווך של היתוך שלפוחיות באקסוציטוזיס מתבצע על ידי חלבונים המכונים SNAREs (קולטן חלבון מצורף NSF מסיס). משפחת-על גדולה זו של חלבונים משתרעת על פני טווח ביולוגי רחב, משמרים ועד יונקים. היתוך שלפוחית נפוץ מתרחש כאשר שלפוחיות סינפטיות עוגנות עם נוירונים (איור 3.61) ומשחררות נוירוטרנסמיטורים. אלה נלמדים היטב. ה-SNAREs המעורבים בתהליך זה יכולים להיבקע פרוטאוליטית על ידי נוירוטוקסינים חיידקיים המולידים את התנאים של בוטוליזם וטטנוס.
SNAREs נמצאים בשני מיקומים. V-SNARES נמצאים בממברנות של שלפוחיות הובלה במהלך תהליך הנביטה, ואילו T-SNARES ניתן למצוא בממברנות של תאים ממוקדים.
הפעולה של היתוך שלפוחית עולה בקנה אחד עם עלייה של סידן תאיים. היתוך של שלפוחיות סינפטיות בהעברה עצבית גורם להפעלה של תעלות סידן תלויות מתח בתא הממוקד. זרם של סידן עוזר לעורר היתוך שלפוחית. במערכת האנדוקרינית, קשירת הורמונים לקולטנים מצמידים לחלבון G מפעילה את מערכת ה- IP3/DAG להגברת רמות הסידן.
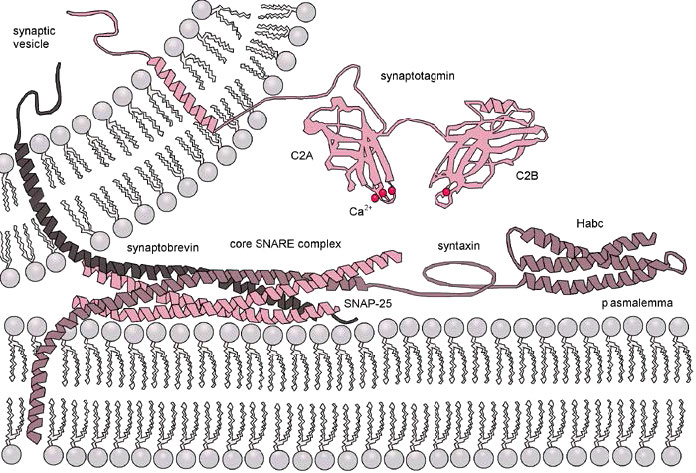
בתהליך של היתוך ממברנה (איור 3.62), ה-V-SNARES של שלפוחית הפרשה (למעלה משמאל) מקיימים אינטראקציה עם ה-T-snares של קרום מטרה (למטה). ה- v- ו- T-snares "רוכסנים" את עצמם יחד כדי לקרב את שלפוחית הממברנה ואת קרום המטרה זה לזה.
רוכסן גורם גם לשטוח ולמתח לרוחב של משטחי הממברנה המעוקלים, ומעדיף המיפוזיה של השכבות החיצוניות של כל קרום. מתח מתמשך גורם לאיחוי עוקב גם של הממברנות הפנימיות, ומניב פתיחה של תוכן תא השלפוחית למטרה שלו (בדרך כלל מחוץ לתא).
שאטלים
דרך נוספת להעביר פריטים על פני קרום שאין לו מערכת תחבורה ספציפית זמינה היא השימוש במעבורות. מעבורות חשובות כאשר אין מנגנון הובלה להעברת חומר על פני קרום שאין לו מערכת הובלה.
דוגמה מצוינת היא NADH. NADH הוא נשא אלקטרונים חשוב המיוצר בציטופלזמה ובמיטוכונדריה של התא. NADH המיוצר במיטוכונדריון עובר ישירות למערכת הובלת האלקטרונים ומספק אלקטרונים למתחם I. NADH המיוצר בציטופלזמה (כגון מגליקוליזה) אין אפשרות זו, שכן הממברנה הפנימית של המיטוכונדריון אטומה למולקולה ולא קיים טרנספורטר להעביר אותו לרוחב. החלק החשוב ב- NADH הוא מטען האלקטרונים שלו, ולכן התאים פיתחו שתי דרכים להעביר את האלקטרונים למטריצה המיטוכונדריאלית מלבד NADH.
שתי השיטות כוללות הסעות. בכל מקרה, מולקולת מקבל מקבלת אלקטרונים מ-NADH והצורה המופחתת של מולקולת המקבל מועברת. הוא מועבר למטריצה שם הוא מתחמצן (אלקטרונים הולכים לאיבוד) ואז נתרם למערכת הובלת האלקטרונים.
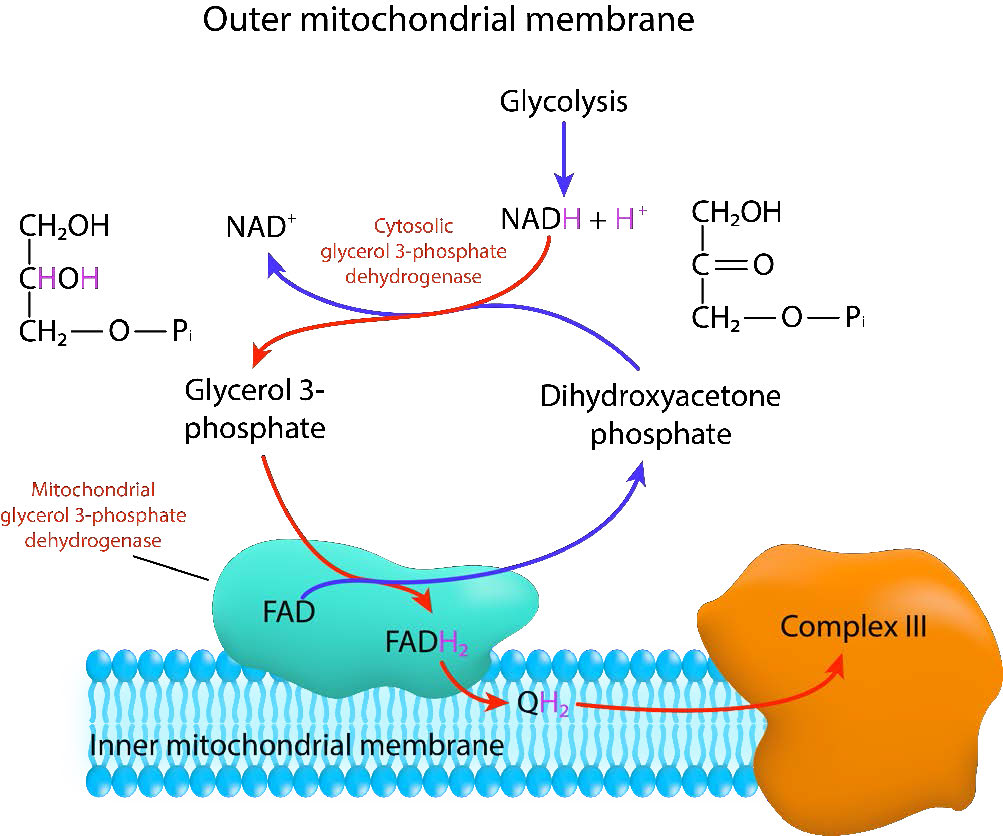
מעבורת פוספט גליצרול
הראשון של שיטות אלה הוא לפחות יעיל, אבל זה מהיר. זה נמצא בדרך כלל בשרירים שיש להם צרכים לאנרגיה מהירה ורקמת מוח. מעבורת זו מכונה מעבורת גליצרול פוספט ומוצגת באיור 3.63. הוא פועל בחלל הבין-ממברני בין הממברנות המיטוכונדריות הפנימיות והחיצוניות. הקרום המיטוכונדריאלי החיצוני נקבובי מאוד, ומאפשר לחומרים רבים לעבור דרכו בחופשיות. בחלל הבין-ממברני, האנזים הציטופלזמי, גליצראלדהיד-3-פוספט דהידרוגנאז (cGpD) מזרז העברת אלקטרונים מ-NADH לפוספט דידידרוקסיאצטון (#2 באיור), ומניב NAD+ וגליצרלדהיד-3-פוספט (#1 באיור). לאחר מכן הגליצרלדהיד-3-פוספט נקשר לגליצראלדהיד-3-פוספט דהידרוגנאז (mGPD) המוטמע בחלק החיצוני של הממברנה המיטוכונדריאלית הפנימית. mGpD מזרז את העברת האלקטרונים מגליצראלדהיד-3-פוספט ל- FAD, ומייצר פוספט דיהיקרוקסיאצטון ו- FADH2. לאחר מכן FADH2 מעביר את האלקטרונים שלו למערכת הובלת האלקטרונים דרך CoQ (Q למעלה), ויוצר CoQH2 (QH2 לעיל). כפי שנדון בסעיף על הובלת אלקטרונים, זו אינה מערכת הסעות יעילה מכיוון שהיא אינה גורמת לייצור של ATP רב כפי שקורה כאשר אלקטרונים מועברים ל- NAD+ במקום ל- FAD.
שאטל מלאטה אספרטט
מערכת יעילה יותר להעברת אלקטרונים היא מעבורת מאלט-אספרטט והיא מוצגת באיור 3.64. כפי שעולה באיור, מעבורת זו כוללת יותר צעדים מאשר מעבורת גליצרול פוספט, אך היתרון של מעבורת מאלט-אספרטט הוא שהיא יעילה יותר. NADH מחוץ למיטוכונדריון מעביר את האלקטרונים שלו למעבורת ואז NADH נוצר מחדש בחלק הפנימי של המעבורת. שום אנרגיה לא מושקעת בתהליך.
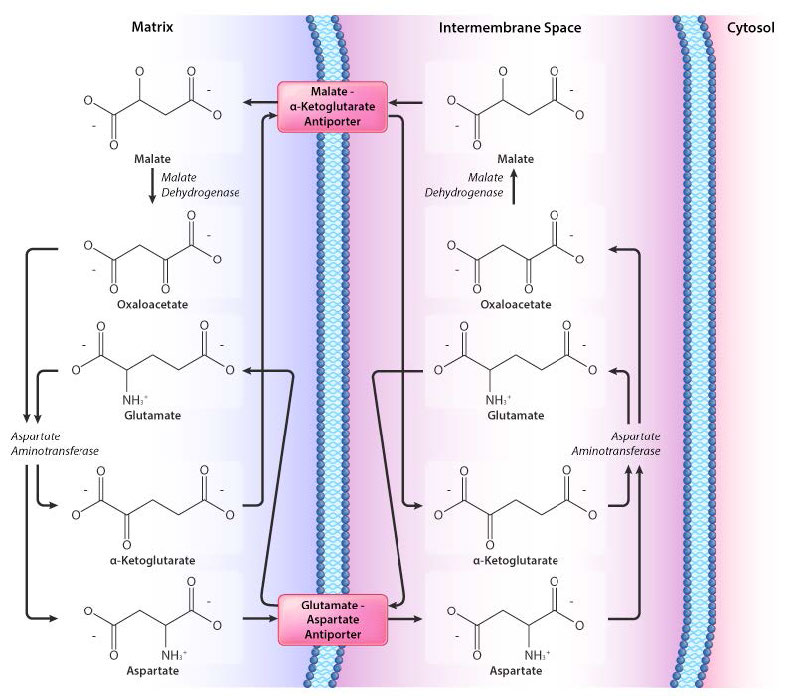
כאשר NADH מצטבר בציטופלזמה, הוא עובר לחלל הבין-ממברני שבו האנזים malate dehydrogenase מזרז את העברת האלקטרונים לאוקסלואצטט כדי להניב NAD+ ומלאט. מערכת הובלה למלט מעבירה מאלאט לתוך המטריצה המיטוכונדריאלית בתמורה ל-α-ketoglutarate.
בתוך המיטוכונדריון, מאלאט מתחמצן מחדש לאוקסלואצטט ואלקטרונים ניתנים ל-NAD+ כדי ליצור מחדש את NADH. לאחר מכן NADH תורם אלקטרונים למתחם I של מערכת הובלת האלקטרונים. זה באמת כל מה שיש למעבורת. השלבים הנותרים הם פשוט לאזן את משוואת התהליך. אוקסלואצטט מקבל קבוצת אמין מחומצה גלוטמית כדי להניב חומצה אספרטית ו-α-ketoglutarate. לאחר מכן אספרטט יוצא מהמיטוכונדריון דרך חלבון הובלה אנטי-פורט המחליף אותו לגלוטמט. סדרה של תגובות במרחב הבין-ממברני מאזנת את המשוואה.
קל ללכת לאיבוד בבלגן של משוואות איזון. הדבר החשוב ביותר להבין כאן הוא שאוקסלואצטט מקבל אלקטרונים מבחוץ כדי להפוך למלט שהוא נושא האלקטרונים על פני הממברנה. ברגע שנכנס למטריצה, מאלאט מומר בחזרה לאוקסלואצטט והאלקטרונים שלו ניתנים ל- NAD+ ויוצרים NADH. כל השאר הוא איזון משוואות פשוט.
מעבורת אצטיל-CoA
סוג אחר של מעבורת כולל גם את המיטוכונדריון ובמקרה זה, הפריט המועבר הוא מולקולה, לא זוג אלקטרונים. המולקולה המעניינת כאן היא אצטיל-CoA, שלגביה לא פועלת מערכת הובלה, אך יש צורך בציטופלזמה לסינתזת חומצות שומן כאשר לתא יש אנרגיה בשפע.
אצטיל-CoA נמצא בעיקר במיטוכונדריון וכל עוד מחזור חומצת הלימון פועל ביעילות, ריכוזו יציב יחסית. עם זאת, כאשר מחזור חומצת הלימון מאט, אצטיל-CoA והציטראט המיוצר ממנו במחזור מתחילים להצטבר.
קיימת מערכת הובלת קרום לציטראט, ולכן היא מועברת החוצה לציטופלזמה. בציטופלזמה, אנזים המכונה ציטראט ליאז מבקע ציטראט לאצטיל-CoA ואוקסלואצטט. ניתן להפחית את אוקסלואצטט למלט ולהעביר אותו חזרה למיטוכונדריון.
באשר לאצטיל-CoA, ככל שיש יותר תאים אלה בציטופלזמה, כך גדל הסיכוי שהם יתחילו לייצר חומצות שומן ושומן, שכן אצטיל-CoA הוא חומר המוצא לסינתזת חומצות שומן, המתרחשת בציטופלזמה. מתי מתרחש תהליך זה? כפי שצוין לעיל, זה מתרחש כאשר מחזור חומצת הלימון נעצר וזה מתרחש כאשר רמות NADH ו- FADH2 עולות. אלה, כמובן, מתגברים כאשר לא שורפים כמות קלוריות כמו שאוכלים כתוצר לוואי של בקרת נשימה. חוסר פעילות גופנית מוביל לרמות גבוהות יותר של נשאי אלקטרונים מופחתים.
צומת תאים
תאים באורגניזמים רב תאיים נמצאים בקשר הדוק זה עם זה וקישורים ביניהם נקראים צמתים. באורגניזמים של חוליות, ישנם שלושה סוגים עיקריים של צומת תאים ואחד מהם (צומת פערים) חשוב לתנועת חומרים בין תאים. שלושת הסוגים הם
- צומת פערים
- צומת אדהרנס, (צומת עיגון, דסמוזומים והמידסמוזומים)
- צמתים הדוקים
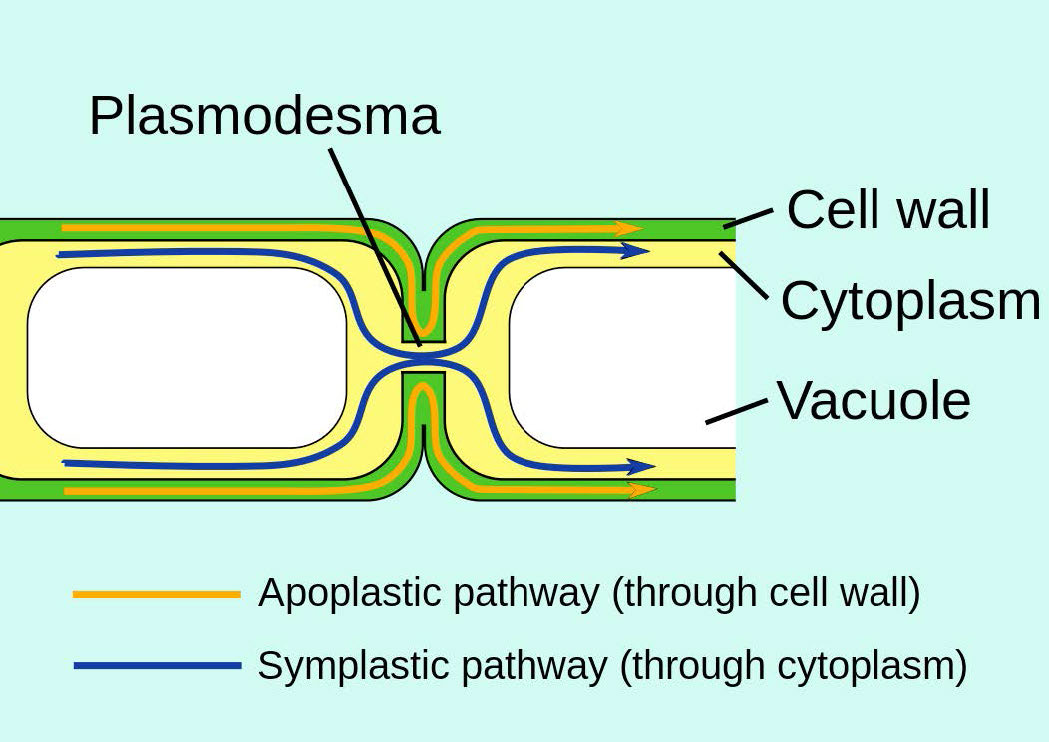
צומת תאים בצמחים רב-תאיים בנויים באופן שונה מאלה שבחולייתנים ונקראים פלסמודסמטה. גם הם מתפקדים בחילופי חומרים בין תאים בודדים.
צומת פערים
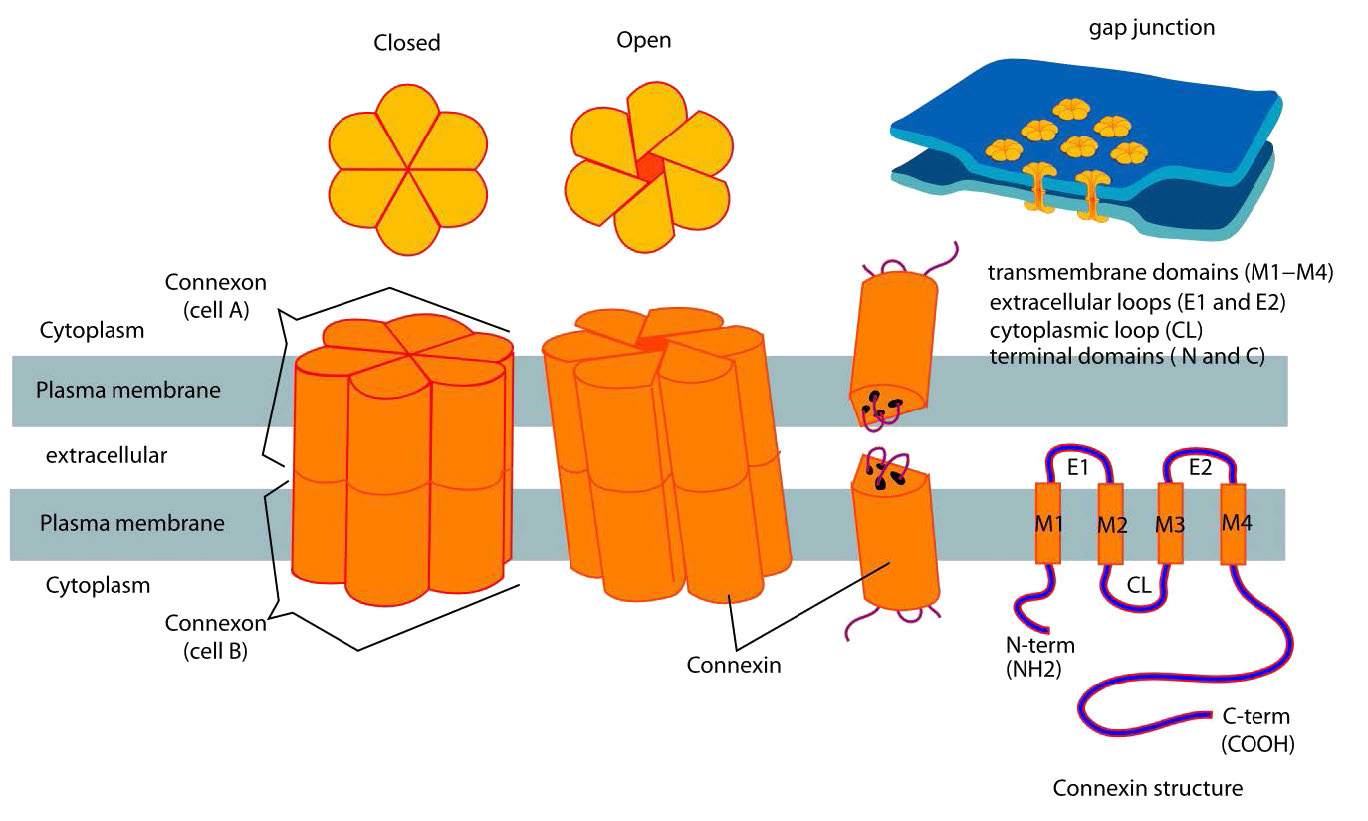
צומת פערים הם מבנים מיוחדים המורכבים משתי קבוצות של מבנים הנקראים קונקסונים (אחד מכל תא - ראה איור 3.65) מקשרים ישירות את הציטופלזמות של התאים המחוברים. צומת פערים מוסדרים כדי לשלוט בזרימת המולקולות, היונים והדחפים החשמליים בין התאים. בצמחים, מבנים דומים המכונים plasmodesmata חוצים את דופן התא (איור 3.66) ומבצעים פונקציות דומות.
צומת אדהרנס
צומת Adherens (איור 3.67) הם מתחמי חלבון בצד הציטופלזמי של ממברנות התא של רקמות אפיתל ואנדותל המקשרים תאים זה לזה או למטריצה החוץ-תאית. הם תואמים את ה- fascia adherens המצויים בתאים שאינם אפיתליים/לא אנדותל.
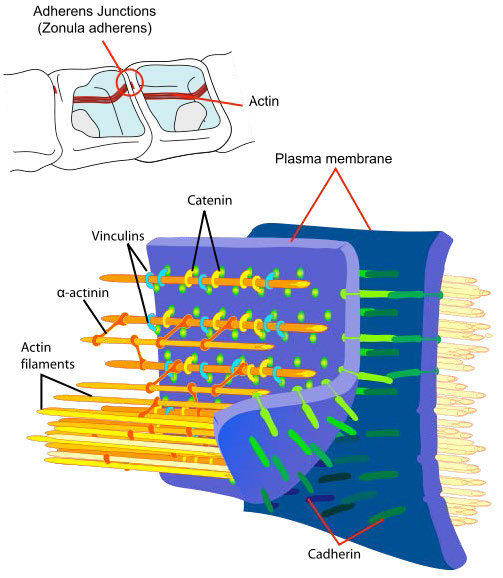
צומת Adherens מכילים את החלבונים הבאים - 1) α-catenin (קושר קדרין דרך β-catenin); 2) β-catenin (התקשרות ל-α-catenin לקדרין; 3) γ-catenin (נקשר לקדרין); 4) קדרינים (קבוצה של חלבונים טרנסממברניים שמתעממים עם קדרינים בתאים סמוכים; 5) p120 (גם נקרא Δ-catenin - נקשר לקדרין); 6) פלקוגלובין (חלבון ממשפחת קטנין הומולוגי ומתנהג כמו β-catenin); 7) אקטין; 8) אקטין; 8) אקטין; ו- 9) וינקולין. צמתים של Adherens עשויים לסייע בשמירה על טבעת התכווצות האקטין הנוצרת בתהליך הציטוקינזיס.
צמתים הדוקים
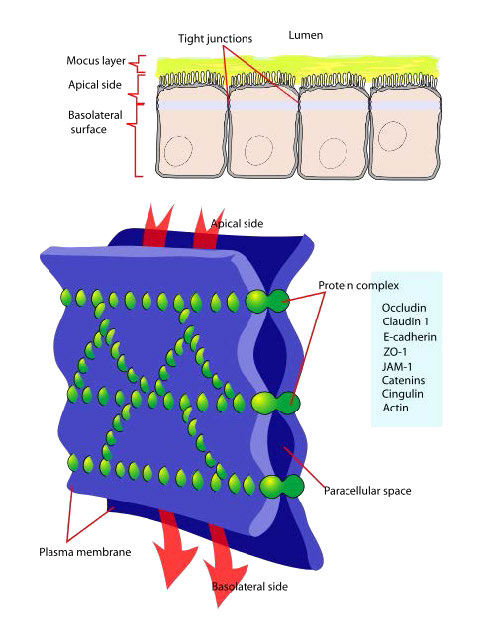
צמתים הדוקים (איור 3.68) הם רשת של גדילי חלבון החותמים תאים יחד ומגבילים את זרימת היונים ברווחים ביניהם. השפעת המבנה שלהם היא להגביל את תנועת החומרים דרך הרקמות על ידי דרישה מהם לעבור דרך תאים במקום סביבם. צמתים הדוקים מחברים יחד את השלד הציטוני של התאים ובאמצעות המבנה שלהם שומרים על הקוטביות האפיקית והבזולטרלית שלהם.
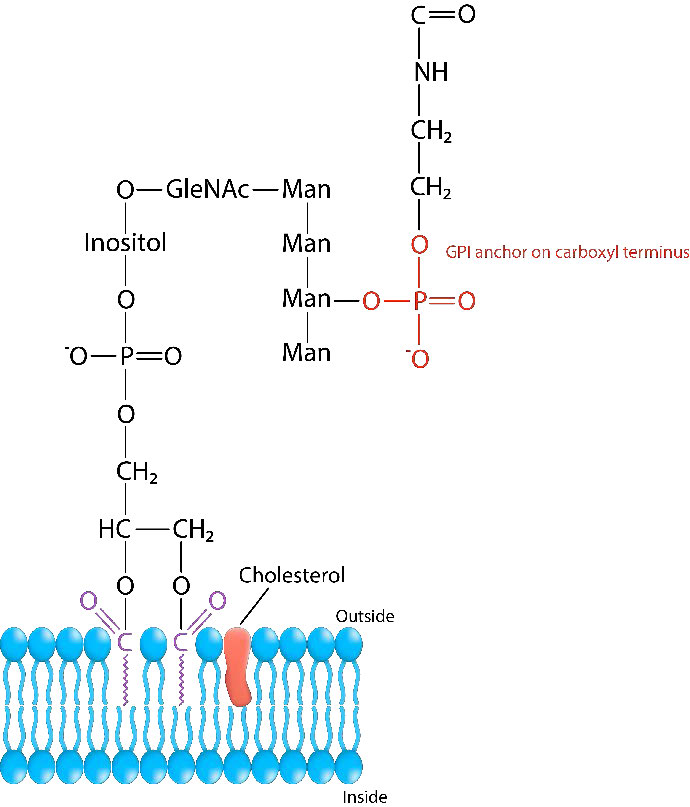
עוגני GPI
חלבוני ממברנה המחוברים לגליקוזילפוספטידילינוזיטול (הידוע גם כעוגן GPI) מכונים גליפיאטים. החלבונים, הממלאים תפקידים חשובים בתהליכים ביוכימיים רבים, מחוברים לעוגן ה- GPI בקצה הקרבוקסיל שלהם. פוספוליפאזים, כגון פוספוליפאז C יכולים לחתוך את הקשר בין החלבון ל- GPI, ולשחרר את החלבון מממברנת התא החיצונית. לחלבונים המיועדים לגליפציה יש שני רצפי אותות. הם 1) רצף אותות N-טרמינלי ו -2) רצף אותות מסוף C המוכר על ידי טרנסאמידאז GPI (GPIT). רצפי האותות N-טרמינליים אחראים על הכוונת הובלה משותפת לתרגום לרשת האנדופלזמית. הרצף C-terminal מזוהה על ידי GPI transamidase, המקשר את קצה הקרבוקסי של חלבון לעוגן GPI.
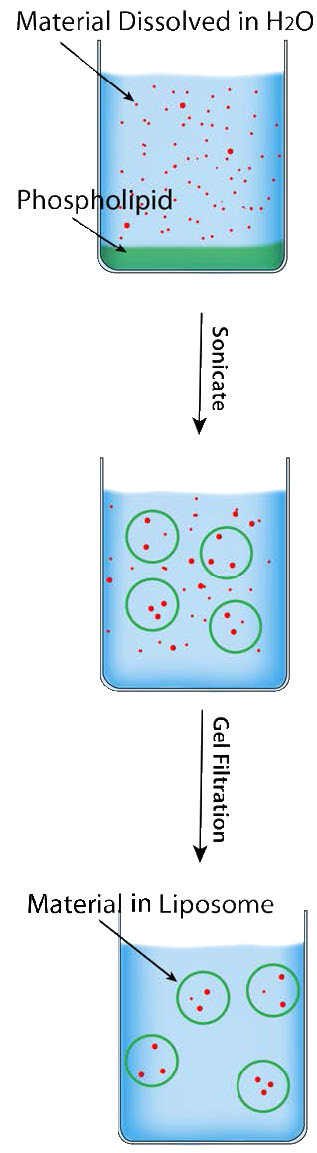
ליפוזומים
היכולת הספונטנית של תרכובות פוספוגליצרוליפידים וספינגוליפידים ליצור דו-שכבות שומנים מנוצלת ביצירת מבנים ממברניים מלאכותיים הנקראים ליפוזומים (איור 3.69). ליפוזומים שימושיים להעברת תוכנם לתאים באמצעות היתוך ממברנה. באיור, פריטים המיועדים למסירה לתאים יהיו עטופים באזור המעגלי האמצעי של הליפוזום וכאשר הליפוזום מתמזג עם קרום התא, הוא יעביר את התוכן ישירות לציטופלזמה.
מדד הידרופתיה
החלק הפנימי של דו שכבת השומנים הוא הידרופובי מאוד, מה שהופך אותו למגביל מאוד לתנועה של יונים וחומרים קוטביים על פניו. מאפיין זה גם מציב מגבלות על סוגי חומצות האמינו שיתקשרו איתו גם כן. בגלל זה, תחומי חלבון טרנסממברניים המצויים בחלבוני ממברנה אינטגרליים מוטים בחומצות האמינו המקיימות אינטראקציה עם דו שכבת השומנים או עם החומר המימי משני צידיה.
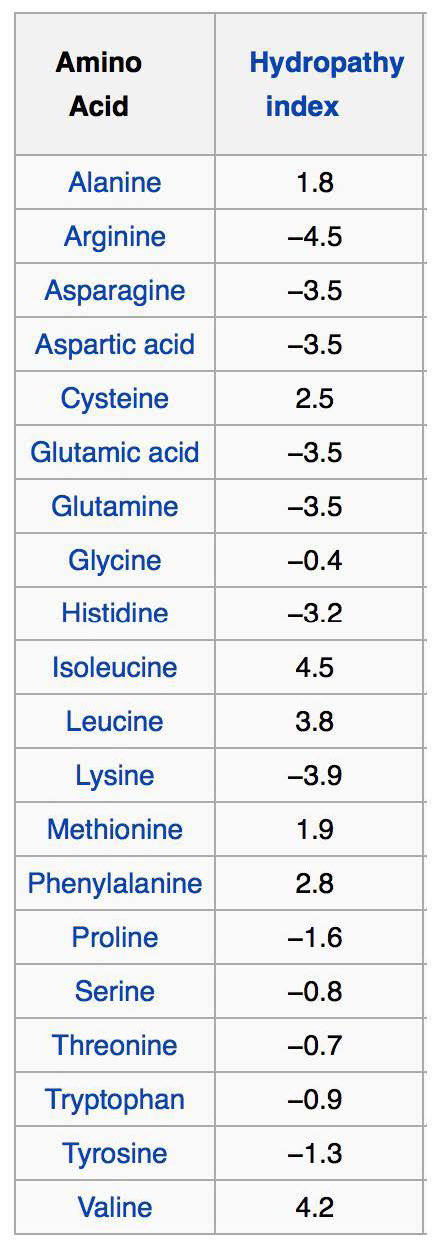
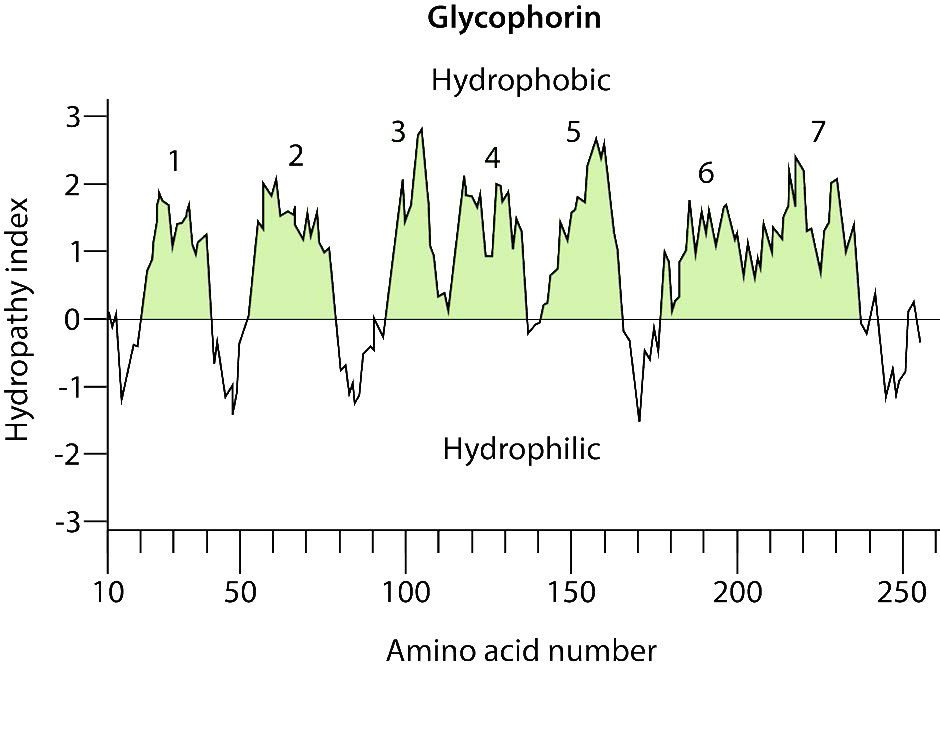
חומצות אמינו הידרופוביות נמצאות בתוך הדו -שכבה, ואילו חומצות אמינו הידרופיליות נמצאות בעיקר על המשטחים. רמז נוסף לזיהוי אזורים חוצים ממברנה של חלבון הוא שטריפטופן או טירוסין ממוקמים בדרך כלל בממשק (ים) לא קוטבים/קוטביים של דו שכבת השומנים לחלבונים אינטגרליים. ניתן לזהות ארגון כזה של חומצות אמינו על ידי ניתוח ממוחשב של רצפי חומצות אמינו באמצעות מה שמכונה אינדקס/ציון הידרופתיה (איור 3.71). למרות שהשמות והניקוד משתנים, הרעיון הוא להקצות מספר (בדרך כלל חיובי) לשרשראות צד של חומצות אמינו בעלות הידרופוביות גבוהה יותר ושליליות לאלו היוניות. עם ציונים אלה, תוכנת מחשב יכולה למצוא בקלות את הציונים הממוצעים של מקטעי חומצות אמינו קצרים (נניח באורך 3 חומצות אמינו) ולאחר מכן לשרטט ערכים אלה בגרף של מדד הידרופוביות לעומת מיקום בשרשרת הפוליפפטיד. פעולה זו עבור חלבון טרנסממברני כגון גליקופורין גורמת לעלילה המוצגת באיור 3.72. ניכר בניתוח שמדובר בחלבון טרנסממברני שיש לו שבעה תחומים החוצים את דו שכבת השומנים, כפי שסומן.
קירות תאים
קירות תאים נמצאים בתאים רבים, כולל צמחים, פטריות וחיידקים, אך אינם נמצאים בתאי בעלי חיים. הם נועדו לספק חוזק ויושרה ולפחות הגנה מסוימת מפני התפרצות מלחץ אוסמוטי (איורים 3.73-3.75).

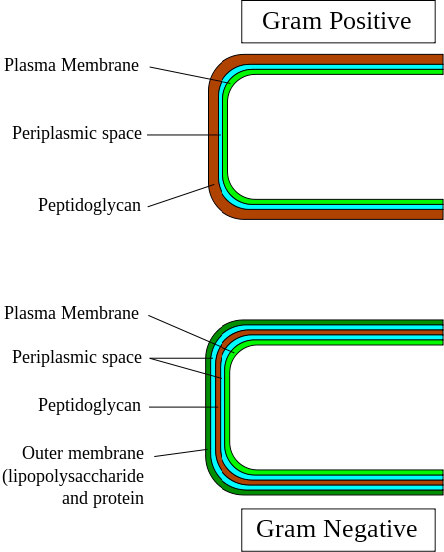
לחיידקים גראם חיוביים (איור 3.75) יש את עיצוב דופן התא הפשוט ביותר. במעבר מחוץ לתא לכיוון הציטופלזמה ישנה שכבה פפטידוגליקנית חיצונית לדופן התא ואחריה חלל פריפלזמי, קרום פלזמה ולאחר מכן הציטופלזמה. חיידקים גראם שליליים מוסיפים שכבת הגנה שנייה חיצונית לכל זה, אז מבחינתם, מתחילים מבחוץ ונעים פנימה, נתקלים בשכבת ליפופוליסכריד חיצונית, חלל פריפלזמי, דופן התא הפפטידוגליקני, חלל פריפלזמי שני, קרום פלזמה ואז הציטופלזמה.
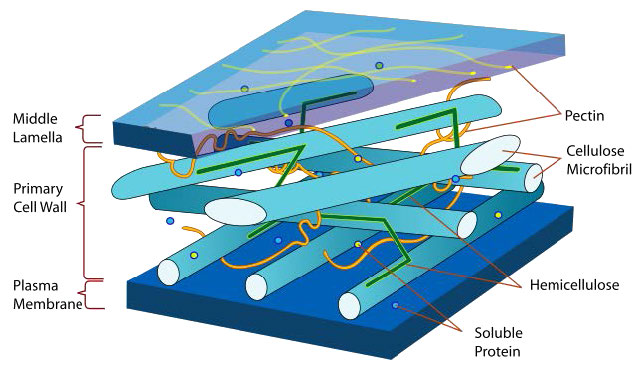
לצמחים עשבוניים יש דופן תא חיצונית נוקשה (המורכבת בעיקר מתאית, המיצלולוזה ופקטין) וקרום פלזמה פנימי. צמחים עציים מוסיפים מפלס שני של קיר עם ליגנין בין דופן התאית לקרום הפלזמה של צמחים עשבוניים.
BB ארץ הפלאות
למנגינה של "ארץ הפלאות החורף"
אתר מנגינות מטבוליות כאן
אולם מילאם - השעה 12:30
ואהרן מקבל מילים
הוא הולך אל וממנו '
בעוד לא מדבר לאט
נותן את זה ל- B-B-4-5-0
שמחתי כשהמונח התחיל
הערות הרצאה וסרטונים בשפע
קובצי MP3 נוספו לאייפוד שלי
אבל לפעמים דקלומים היו משעממים
והבחינות נשכו אותי בערך
כאשר העקומה התבררה מכוערת
אני לא חושב שזה כך
הציונים שלי נמוכים מדי
גולש ליד ב- B-4-5-0
סוף סוף יש בדיקה
ב -9 בדצמבר בשעה 18:00
אני אארוז את הכרטיס שלי עם מידע
אז אני לא צריך לשנן את זה אז
ואני ארגיש כמו חכם
עם כרטיס הפתק הארוז שלי
רק עוד אחד ללכת
ואז הו-הו-הו
אני אסיים עם B-B-4-5-0
הקלטה מאת דיוויד סימונס
מילים על ידי קווין אהרן
הקלטה על ידי דוד סימונס מילים על ידי קווין אהרן
334
תודה לאל שיש סרטון
למנגינה של "תודה לאל שאני ילד כפרי"
אתר מנגינות מטבוליות כאן
יש חבילה של דברים שתלמיד צריך לדעת
והשיחה של אהרן לא ממש איטית
ללמוד זה לא קל/ ההרצאות די קשות
תודה לאל שיש סרטון
ובכן עברנו את המחזורים וגם את האנזימים שלהם
לומדים את הרגולציה הכל חדש
אני חייב להודות שאין לי מושג
מה אני אעשה?
אז השגתי לי כרטיס ברכה וקניתי לי סטרייר
הורידו את האנזימים ואת השמות שהוא דורש
אני מקווה שאוכל לגייס עוד קצת תשוקה
תודה לאל שיש סרטון
בדיוק התעדכנו לגבי
הפרוטונים N-A-D הנעים דרך
אלקטרונים מורכבים של Vee Electrons רוקדים בציטוכרום C
חייבים לשמוע את ה- MP3
חמצון חומצות שומן גורם לאצטיל-CoA
בתוך המטריצה הפנימית של המיטוכונדריה- ay
זה מאוד מסובך, אני מניח שאני חייב לומר
תודה לאל שיש סרטון
אז השגתי לי כרטיס ברכה וקניתי לי סטרייר
הורידו את האנזימים ואת השמות שהוא דורש
אני מקווה שאוכל לגייס עוד קצת תשוקה
תודה לאל שיש סרטון
שכפול די קל בצורה פשוטה
העתקת הבסיסים ב-DNA הפלסמיד Gs הולך עם Cs
ו-Ts הולך עם As הודות לפולימראז
וה-DNA הוא תבנית עבור
סלילי ה-RNA המתפרקים ב-T-A-T-A
סיום קורה, ואז האנזים נעלם
אל תשכח את ה-poly-A
אז קניתי לי כרטיס ברכה וקניתי לי סטרייר
. הורידו את האנזימים ואת השמות שהוא דורש
אני חושב שאני יכול לגייס עוד קצת תשוקה
תודה לאל שיש סרטון
הקלטה מאת דיוויד סימונס
מילים על ידי קווין אהרן
הקלטה על ידי דוד סימונס מילים על ידי קווין אהרן