
3.2: הובלה בממברנות
- Page ID
- 207660

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)מקור: BiochemFFA_3_2.pdf. ספר הלימוד כולו זמין בחינם מהמחברים בכתובת http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy
תנועת חומרים על פני ממברנות
כפי שצוין קודם לכן, חיוני שהתאים יוכלו לספוג חומרים מזינים. פונקציה זו יחד עם תנועה של יונים וחומרים אחרים מסופקת על ידי חלבונים/מתחמי חלבון הספציפיים ביותר לתרכובות שהם מזיזים.
תנועה סלקטיבית של יונים על ידי חלבוני ממברנה וחדירותם הנמוכה ביותר של היונים על פני דו שכבת השומנים חשובים לסיוע בשמירה על האיזון האוסמוטי של התא וגם למתן המנגנון החשוב ביותר עבורו לייצור ATP - תהליך הזרחון החמצוני..
טרמינולוגיה
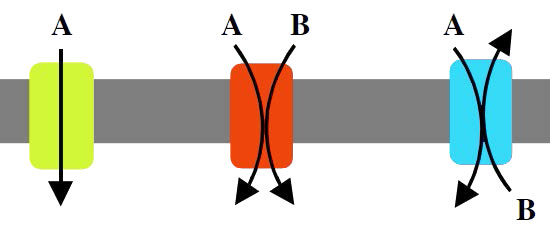
חלבון המעורב בהעברת מולקולה אחת בלבד על פני ממברנה נקרא uniport (איור 3.25). חלבונים המניעים שתי מולקולות באותו כיוון על פני הממברנה נקראים סימפורטים (נקראים גם synporters, synports או symporters). אם שתי מולקולות מועברות בכיוונים מנוגדים על פני הדו -שכבה, החלבון נקרא אנטי-פורט. חלבונים המעורבים בהעברת יונים נקראים יונופורים.
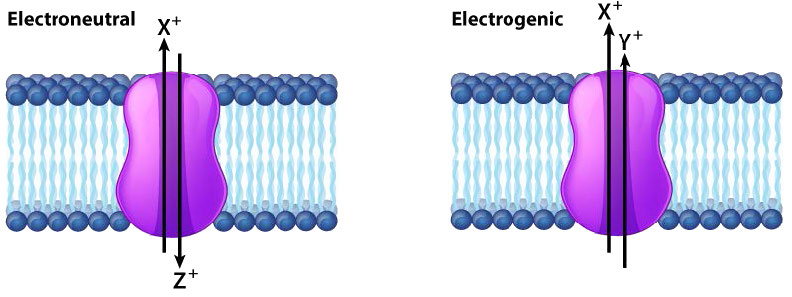
אם פעולתו של חלבון בהעברת יונים על פני ממברנה גורמת לשינוי נטו במטען, החלבון מתואר כאלקטרוגני ואם אין שינוי במטען החלבון מתואר כאלקטרונוטרלי (איור 3.26). כאשר הכוח המניע לתנועה דרך חלבון הממברנה הוא פשוט דיפוזיה, התהליך נקרא דיפוזיה מקלה או הובלה פסיבית וכאשר התהליך דורש קלט אנרגיה אחר, התהליך נקרא הובלה פעילה.
ערוצים ומובילים
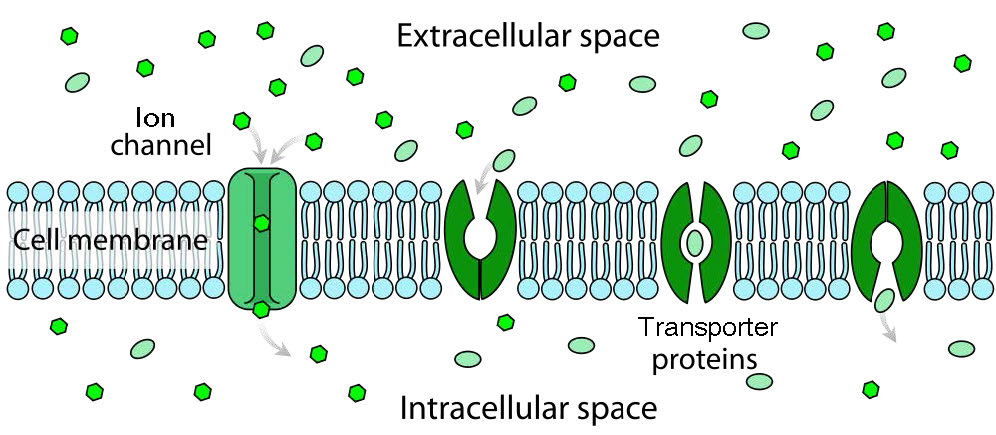
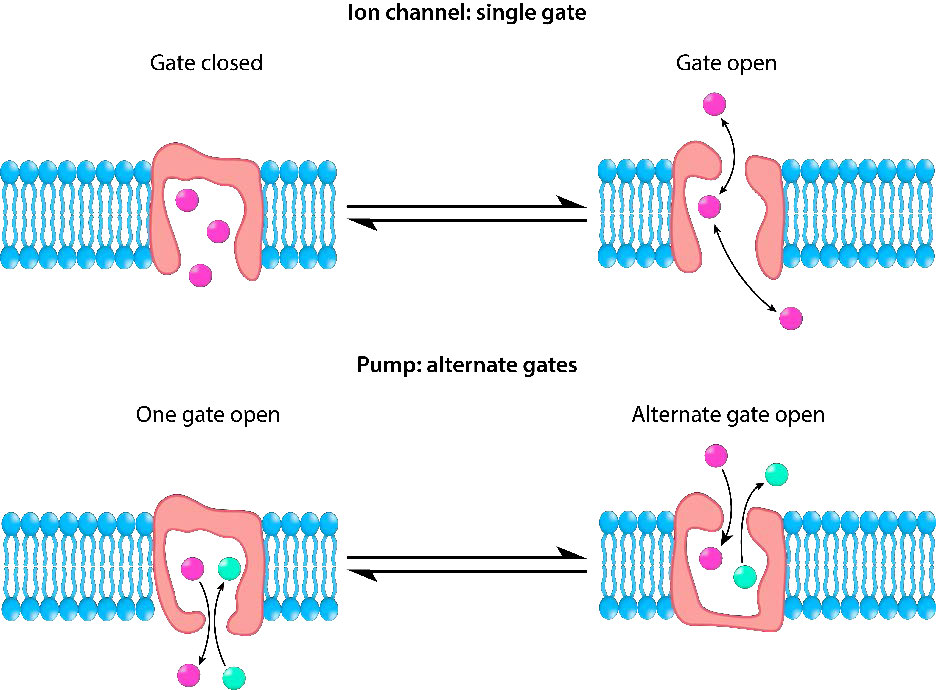
ביחס לתנועת חומרים דרך חלבוני ממברנה, יש הבדל בין תעלות (הנקראות לפעמים נקבוביות) לבין מובילים. ערוצים מספקים במידה רבה פתחים עם ספציפיות מסוימת ומולקולות עוברות דרכם קרוב לקצב הדיפוזיה. בדרך כלל הם כרוכים בתנועה של מים או יונים. דוגמאות לכך הן תעלות הנתרן או האשלגן של תאי עצב. למובילים יש ספציפיות ושיעורי העברה גבוהים שהם איטיים יותר בסדרי גודל. חלבוני הובלה כוללים את משאבת הנתרן-אשלגן, מחליף הנתרן-סידן ופרמאז לקטוז, בין רבים אחרים).
דיפוזיה מקלה
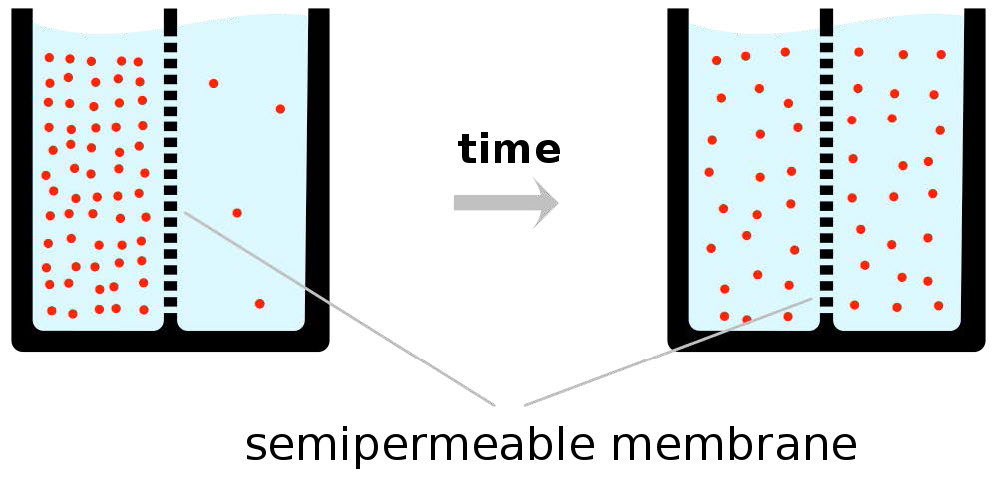
כאמור, הכוח המניע לדיפוזיה קלה הוא ריכוז, כלומר בדיפוזיה קלה, חומרים יעברו רק מריכוז גבוה יותר לריכוז נמוך יותר וכי בסוף התהליך, ריכוז החומרים מכל צד של דו-שכבה יהיה שווה (איור 3.28). זה עשוי לעבוד טוב במקרים רבים.
לדוגמא, ריכוז הגלוקוז בדם גבוה מספיק בכדי שתאי הדם האדומים יוכלו להשתמש בדיפוזיה מקלה כאמצעי לרכישת גלוקוז. תאים אחרים, שהוסרו עוד יותר מאספקת הדם בה ריכוז הגלוקוז נמוך יותר, חייבים להשתמש במנגנוני הובלה פעילים מכיוון שאין ריכוז מספיק של גלוקוז בכדי לספק לתאים את הגלוקוז הדרוש להם.
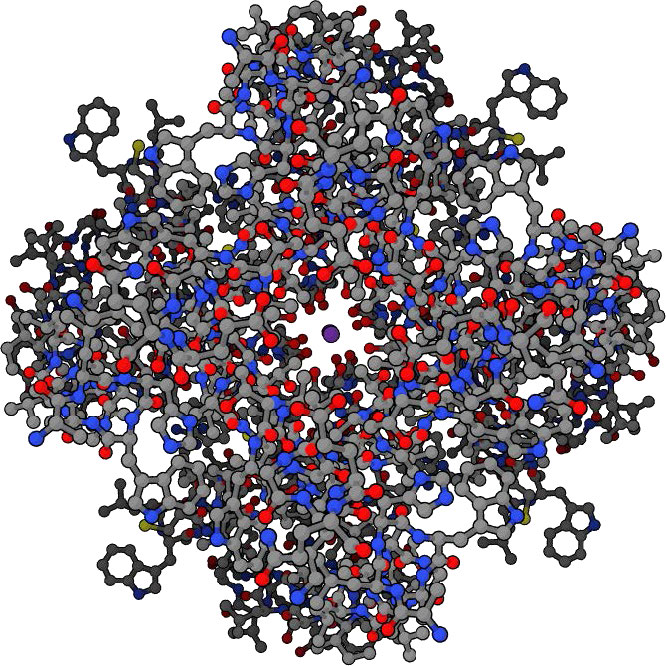
תעלות יונים
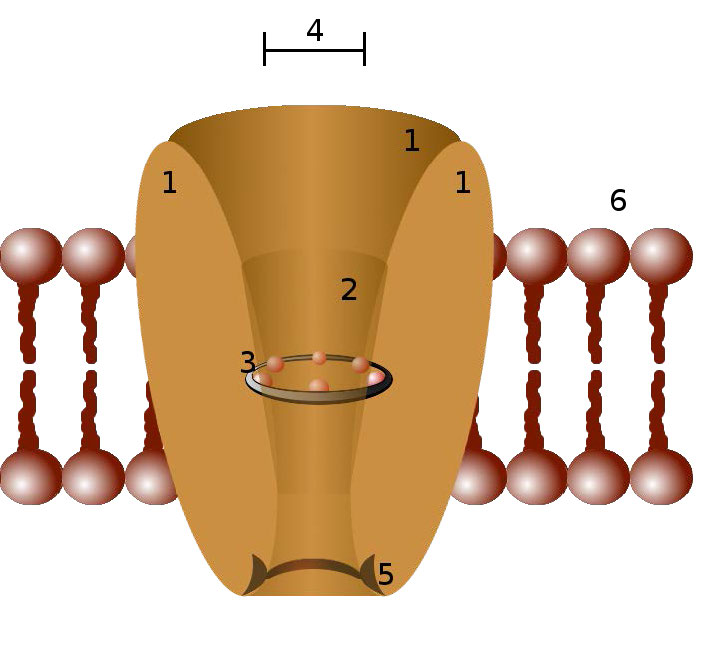
תעלות יונים הן חלבוני ממברנה יוצרי נקבוביות בממברנות של כל התאים המווסתים את התנועה של יונים נבחרים על פני ממברנה (איורים 3.29 & 3.30). הם עוזרים לבסס את פוטנציאל הממברנה המנוחה ולהשפיע על פוטנציאל הפעולה ועל אותות חשמליים אחרים. הם חשובים מאוד בתהליך העברת העצבים. תעלות יונים שולטות בזרימת היונים על פני תאי הפרשה ואפיתל, וכתוצאה מכך מסייעות לווסת את נפח התא על ידי השפעה על הלחץ האוסמוטי.
תעלות יונים הן תכונות חיוניות של כמעט כל התאים, ומתפקדות כ"מנהרות "סלקטיביות המגבילות את התנועה דרכן ליונים בעלי מאפיינים ספציפיים (בדרך כלל גודל). גודל הפתח צר מאוד (בדרך כלל אטום אחד או שניים ברוחב) והוא מסוגל לבחור אפילו כנגד יונים קטנים מדי.
מנגנוני בקרה
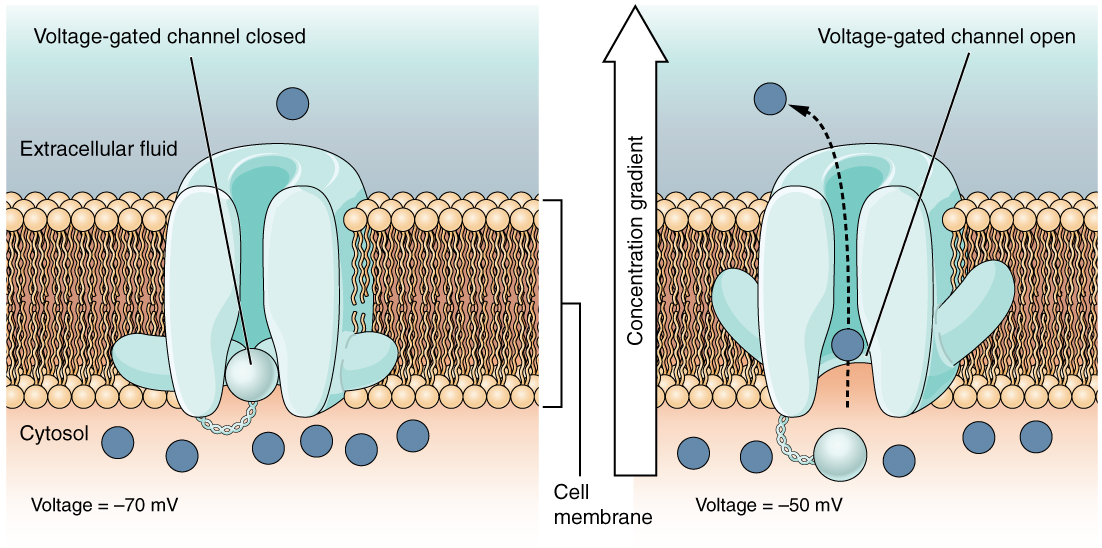
תעלות יונים נשלטות על ידי מנגנונים הכוללים מתח, ליגנדים, אור, טמפרטורה ועיוות מכני (מתיחה מופעלת). תעלות יונים מגודרות ליגנד (LGICs) הן חלבונים טרנסממברניים הנפתחים כדי לאפשר באופן סלקטיבי ליונים כגון Na+, K+, Ca++ או Cl− לעבור דרך הממברנה בתגובה לקשירה של שליח ליגנד.
גלי קול גורמים לעיוות מכני של תאי שיער באוזן. התוצאה היא פתיחת תעלות יונים והתחלת אות עצבי למוח.
תעלות יון נתרן בלשון לקולטני סוכר נפתחות בתגובה לקשירת סוכרוז, ומאפשרות לריכוז הנתרן בתא העצב לעלות וליזום אות עצבי למוח. במקרה זה, ברירת המחדל של השער היא להיות סגורה והוא נפתח בתגובה לקשירה של ליגנד (סוכרוז).
בתאי חישת אור של העין, שערי סידן פתוחים כברירת מחדל, אך גירוי על ידי אור גורם להם להיסגר, מה שמעורר סדרה של אירועים שגורמים לאות שנשלח למוח על תפיסת האור. כך, במקרה זה, הגירוי (האור) גורם לסגירת ערוץ פתוח.
בהנעת הכיוון השני, אותות עצביים שמקורם במוח עוברים לרקמת השריר ובאמצעות מערכת חילופי דברים מסובכת מביאים לפתיחת שערי סידן של תאי שריר, מגבירים את ריכוז הסידן וממריצים את התכווצות השרירים (ראה כאן).
תעלות מגודרות מתח חיוניות להעברת אותות עצביים, תהליך שנדון לעומק יותר כאן.
תנועת יונים דרך תעלות
היכולת של תעלות יונים לבחור מול יונים גדולים מדי היא אינטואיטיבית - גודל הפתח בתעלת היונים פשוט אינו גדול מספיק כדי שיון גדול יותר ייכנס דרך הפתח. אשלגן, למשל, עובר בתעלות נתרן לעיתים רחוקות מכיוון שהפתח קטן מדי.
תעלות אשלגן שהן סלקטיביות ליוני אשלגן חייבות להיות גדולות מספיק כדי לאפשר כניסת אשלגן, אך אם הגודל היה אמצעי הבחירה היחיד, אז יוני נתרן היו עוברים בקלות גם בתעלות אשלגן, מכיוון שיוני נתרן (0.95 Å) קטנים מיוני אשלגן (1.33 Å). על מנת שתעלות אשלגן יבחרו מול יוני נתרן ויעדיפו יוני אשלגן, שיקולים אחרים נכנסים לתמונה.
מעטפת הידרציה
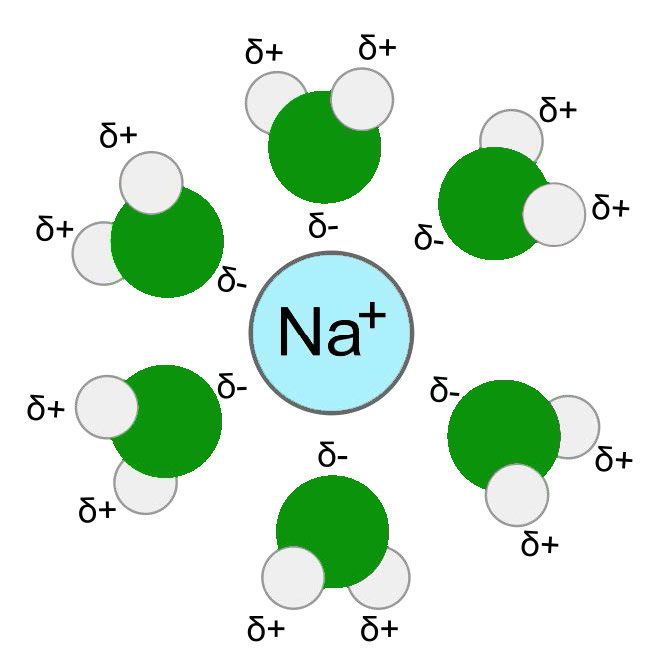
כדי להבין את הסלקטיביות הייחודית הזו, חשוב להבין כיצד יונים נעים בערוצים. לפני שיון יכול לעבור דרך תעלה, יש לנתק אותו תחילה ממולקולות המים בקליפת ההידרציה שלו - מולקולות מים המקיפות יונים בתמיסות מימיות (איור 3.32). תהליך זה דורש קלט של אנרגיה. האנרגיה הראשונית הנדרשת להפשיט את מולקולות המים ממעטפת ההידרציה הושוותה לאנרגיית ההפעלה של תגובה אנזימטית.
דומה לאנזימים
כשם שאנזימים מורידים את אנרגיית ההפעלה של תגובות אנזימטיות ובכך מאפשרים להם להתרחש בקלות רבה יותר, כך גם חלבוני התעלה מורידים את דרישות האנרגיה למולקולה לחצות דו שכבה שומנית. בהיעדר חלבון התעלה, אנרגיית ההתייבשות אוסרת בעיקר על רוב המולקולות הקוטביות להתרחש, כך שמעטים מאוד עוברים אותה על פני שכבת השומנים ללא חלבון התעלה. זו הסיבה שחלבוני תעלת יונים/הובלה חשובים כל כך לתא.
לאחר הפשטת המים, היון יכול לעבור דרך התעלה וכאשר הוא מגיע לצד השני של התעלה, היון המתפזר מתייבש מחדש, ובכך מחזיר לעצמו את האנרגיה שנדרשה בתחילה כדי להסיר את מולקולות המים מהיון.
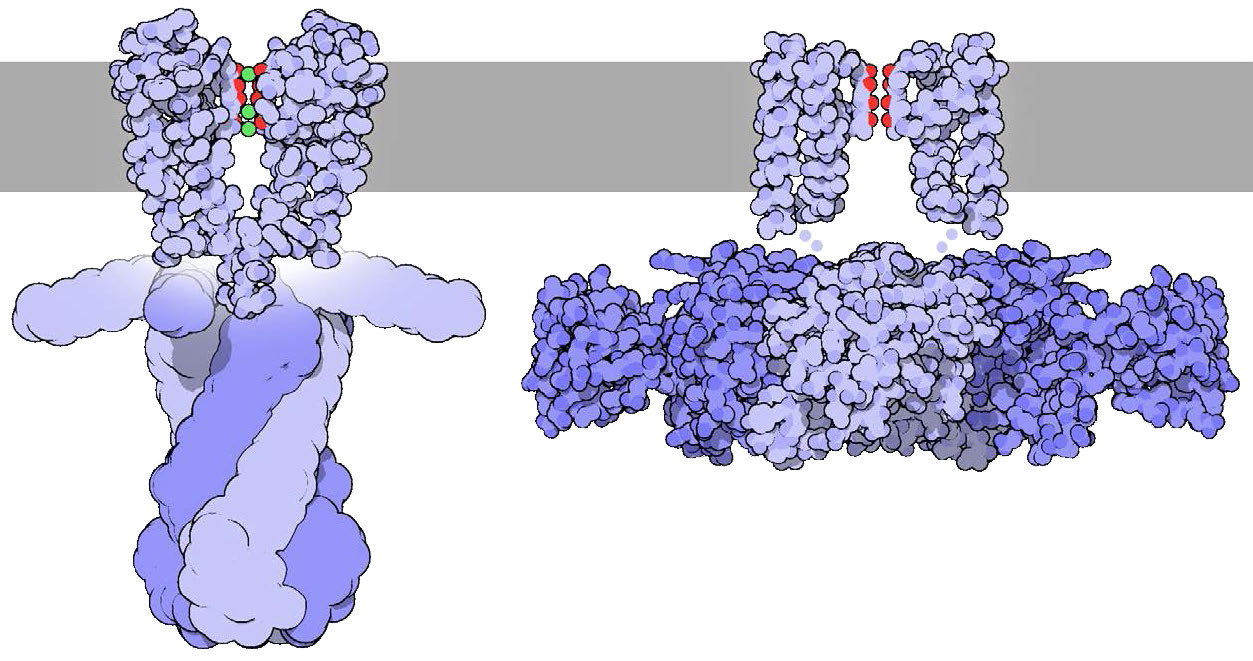
סלקטיביות של תעלת האשלגן
תעלת האשלגן (איור 3.33) משתמשת בממדי יון האשלגן בדיוק כדי לרעות אותו דרך התעלה. יון הנתרן, בעל ממדים שונים מתקשה יותר לעבור אותו דרך התעלה למרות גודלו הקטן יותר. הסיבה לכך נעוצה באנרגיה הנדרשת להתייבשות.
עבור יוני אשלגן, לאחר הפשטת המים, קבוצות קרבוניל הממוקמות במדויק לאורך התעלה עוזרות לייצב את היון תוך כדי תנועה. יון הנתרן, לעומת זאת קטן מדי ואינו יוצר קשרים יעילים עם קבוצות קרבוניל ובכך יש לו דרך קשה יותר. בגלל זה, הפרש האנרגיה בין התייבשות להתייבשות של יון נתרן בתעלת אשלגן אינו חיובי מבחינה אנרגטית (דורש הזנת אנרגיה נטו) אך אותו תהליך עבור יון אשלגן הוא חיובי מבחינה אנרגטית (מביא לרווח נטו של אנרגיה).
סרט 3.1 - גרמידין בויקיפדיה (GIF מונפש, הורד לצפייה)
גורם אנרגיה
לפיכך הבחירה לטובת אשלגן ונגד יוני נתרן בתעלת אשלגן מבוססת על אנרגיה, לא על גודל פיזי, ואילו בבחירת יוני נתרן על פני יוני אשלגן בתעלת נתרן, הגודל הוא השיקול העיקרי.
איזון יונים
תנועת היונים על פני דו שכבה שומנית מווסתת היטב, ומסיבה טובה. שמירה על איזון תקין של יונים בתוך ומחוץ לתאים חשובה לשמירה על איזון אוסמוטי. זה חשוב גם בתוך ומחוץ לאברונים כמו המיטוכונדריה והכלורופלסטים לייצור אנרגיה. אם האיזון היוני של תא מופרע מספיק על ידי יונופור בלתי מבוקר, תא עלול למות.
גרמיצידין
גרמיצידינים (סרט 3.1) הם פוליפפטידים אנטיביוטיים המסונתזים על ידי חיידק האדמה המכונה Bacillus brevis. הפנטדקפפטידים הקטנים הללו (15 חומצות אמינו) מסונתזים על ידי החיידק כדי להרוג חיידקים אחרים.
כאשר הם משוחררים על ידי Bacillus brevis, הגרמיצידינים מכניסים את עצמם לממברנות של חיידקים גראם חיוביים ומאפשרים תנועה של יוני נתרן לתאי המטרה, ובסופו של דבר הורגים אותם. גרמיצידינים יכולים גם לגרום למוליזה בבני אדם ולכן לא ניתן להשתמש בהם באופן פנימי, אלא משתמשים בהם באופן מקומי.
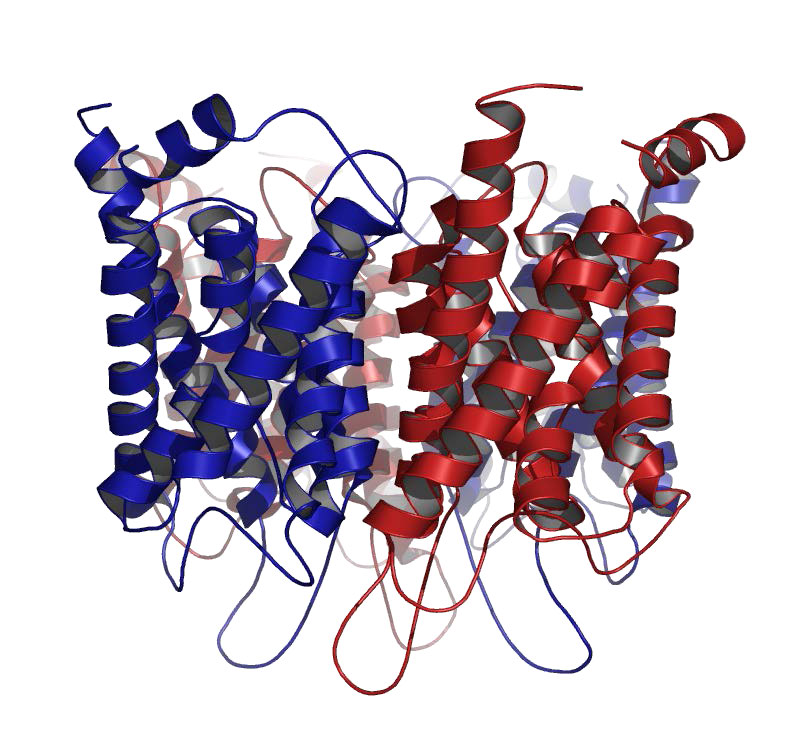
אקוופורינים
אקוופורינים הם חלבוני ממברנה אינטגרליים המכילים נקבוביות המאפשרים באופן סלקטיבי מעבר של מולקולות מים פנימה והחוצה מהתא, תוך מניעת תנועת יונים ומומסים אחרים (איורים 3.34 & 3.35). כמה אקוופורינים הנקראים aquaglyceroporins, מעבירים גם ישויות קטנות אחרות שאינן טעונות, כגון גליצרול, אמוניה, אוריאה ו-CO2, על פני הממברנה,. נקבוביות המים אטומות לחלוטין למולקולות טעונות, כגון פרוטונים, החשובים לשמירה על הפרש הפוטנציאל האלקטרוכימי של הממברנה.
פורינים
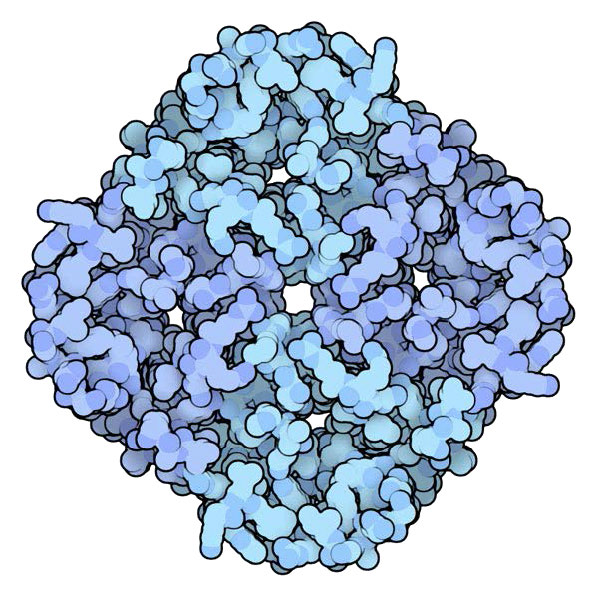
פורינים הם חלבונים המכילים מבנה β-חבית החוצה את קרום התא/דופן ופועל כנקבובה/תעלה שדרכה מתפזרות מולקולות ספציפיות. פורינים נמצאים בקרום החיצוני של חיידקים גראם שליליים וכמה חיידקים גראם חיוביים, מיטוכונדריה וכלורופלסטים.
פורינים בדרך כלל מעבירים רק קבוצה אחת של מולקולות או, במקרים מסוימים, מולקולה ספציפית אחת. אנטיביוטיקה, כגון β-lactam ו- fluoroquinolone עוברים דרך פורינים כדי להגיע לציטוזול של חיידקים גראם שליליים. חיידקים עלולים לפתח עמידות לאנטיביוטיקה אלה כאשר מתרחשת מוטציה לפורין המעורב שגורמת להדרה של האנטיביוטיקה שאחרת תעבור דרכה.
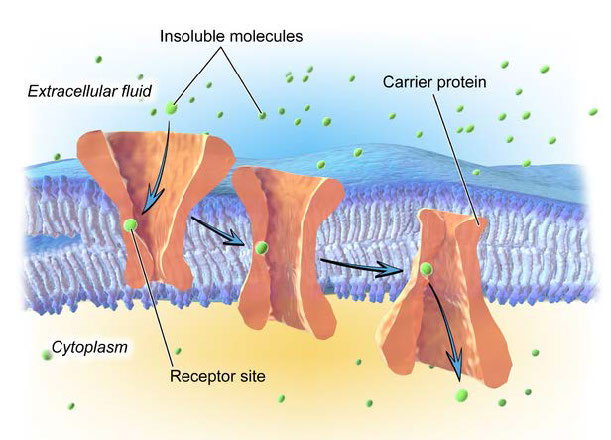
חלבוני טרנספורטר
לא כל ההובלה הקלה מתרחשת דרך חלבוני תעלות יונים. חלבוני טרנספורטר, כפי שצוין קודם לכן (כאן ואיור 3.27) מקלים על תנועת חומרים על פני דו שכבה שומנית, אך הם איטיים יותר מתעלות יונים. איור 3.36 ממחיש חלבון טרנספורטר בפעולה. כפי שניתן לראות, חלבוני טרנספורטר מסתמכים על אתר קולטן ספציפי לצורך זיהוי נכון של המולקולה שיש להזיז.
קישור המולקולה הנכונה גורם לשינוי קונפורמטיבי בצורת החלבון (היפוך) וכתוצאה מכך היפוך הצד הפתוח של החלבון לצד השני של דו שכבת השומנים. בדרך זו, המולקולה מועברת. בדומה לתעלות יונים, חלבוני הטרנספורטר מקלים על תנועת חומרים לכל כיוון, מונעים רק על ידי הפרש הריכוז בין צד אחד לשני.
הובלה פעילה
כל מנגנוני ההובלה שתוארו עד כה מונעים אך ורק על ידי שיפוע ריכוז - נע מריכוזים גבוהים יותר לכיוון ריכוזים נמוכים יותר. תנועות אלו יכולות להתרחש בכל כיוון, וכאמור, לגרום לריכוזים שווים משני צידי הדו -שכבה, אם מותר להם לעבור להשלמה. אולם פעמים רבות התאים חייבים להזיז חומרים כנגד שיפוע ריכוז וכאשר זה קורה, נדרש מקור אנרגיה נוסף. תהליך זה ידוע בשם תחבורה פעילה.
הגדרה טובה להובלה פעילה היא שבהובלה פעילה, לפחות מולקולה אחת מועברת כנגד שיפוע ריכוז. מקור אנרגיה נפוץ, אך לא בלעדי, הוא ATP (ראה Na+/K+ ATPase), אך משתמשים גם במקורות אנרגיה אחרים. לדוגמה, טרנספורטר הנתרן-גלוקוז משתמש בשיפוע נתרן ככוח להובלה פעילה של גלוקוז לתא. לפיכך, חשוב לדעת שלא כל הובלה פעילה משתמשת באנרגיית ATP.
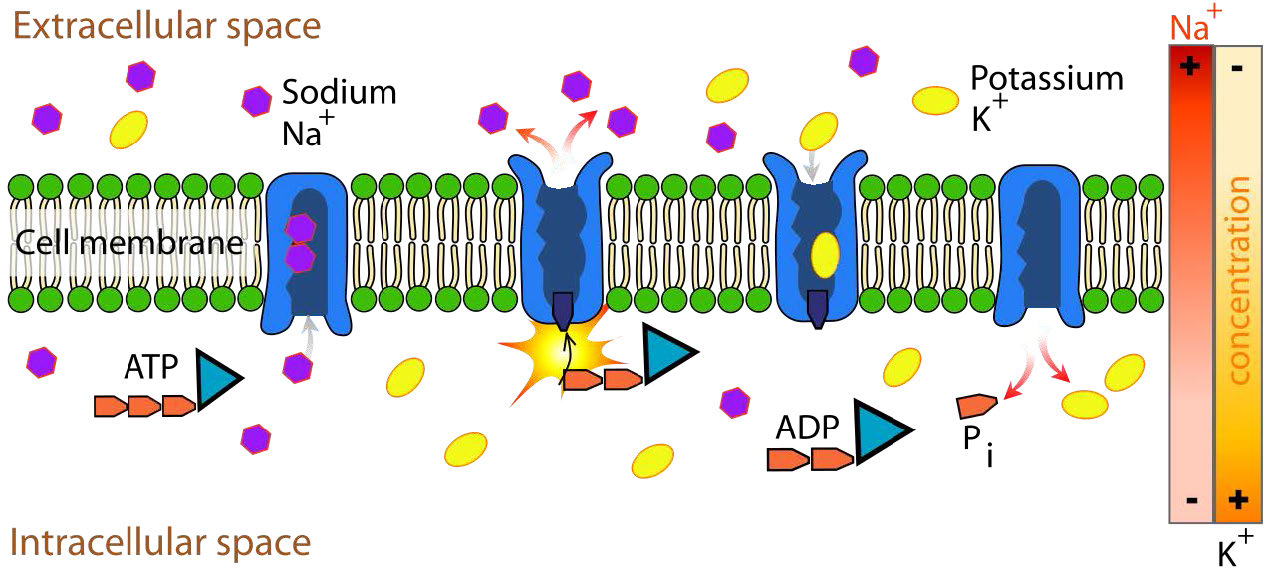
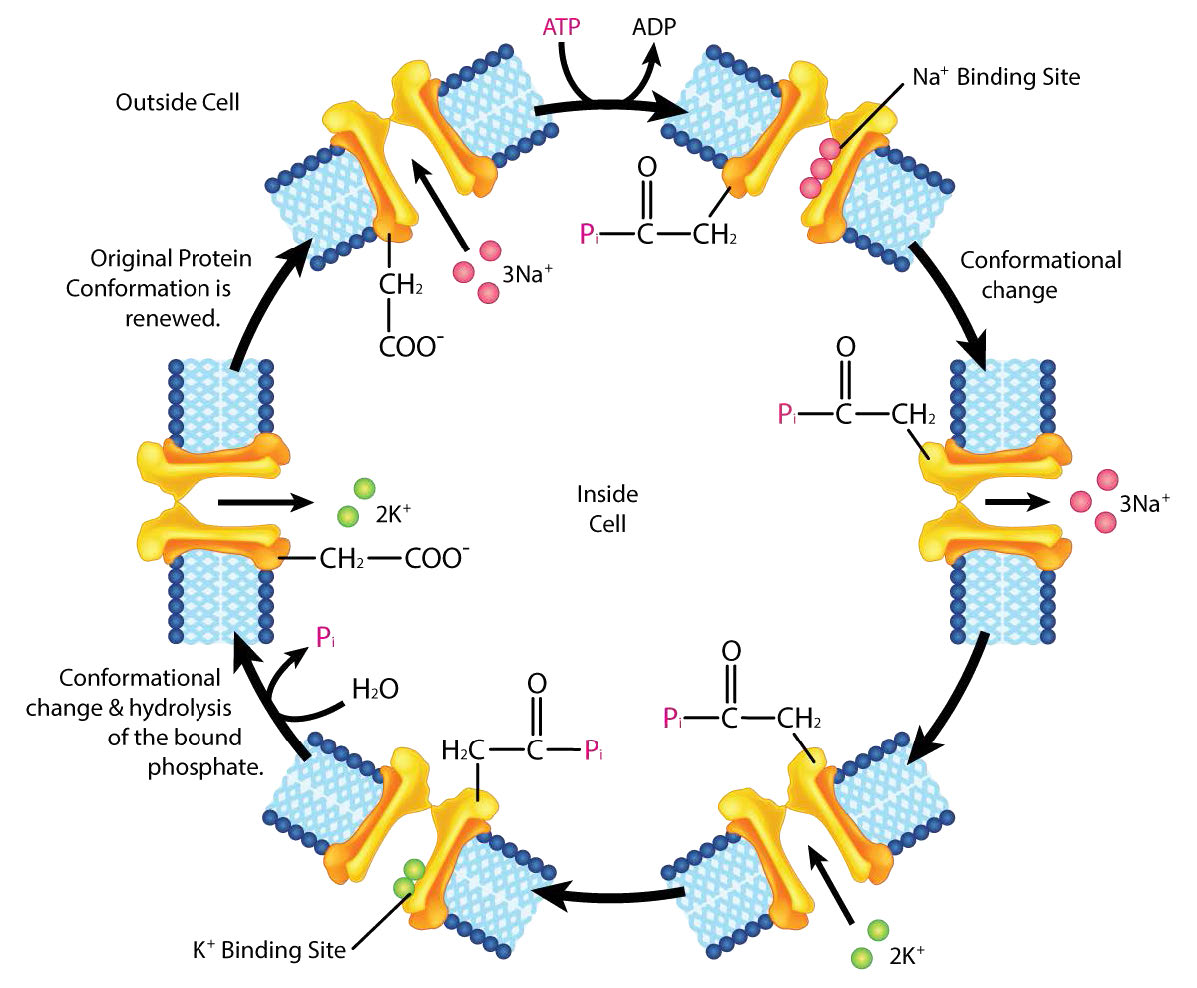
Na+/K+ ATPase
חלבון אינטגרלי חשוב להובלת ממברנה הוא אנטי-פורט Na+/K+ ATPase (איורים 3.37 ו-3.38), המעביר שלושה יוני נתרן אל מחוץ לתא ושני יוני אשלגן לתוך התא בכל מחזור פעולה. בכל מקרה, תנועת היונים היא כנגד שיפוע הריכוז. מכיוון ששלושה מטענים חיוביים מועברים החוצה עבור כל שני מטענים חיוביים המועברים פנימה, המערכת היא אלקטרוגנית.
החלבון משתמש באנרגיה של ATP ליצירת שיפועי יונים החשובים הן בשמירה על לחץ אוסמוטי תאי והן (בתאי עצב) ליצירת שיפועי הנתרן והאשלגן הדרושים להעברת אותות. כשל בתפקוד המערכת גורם לנפיחות של התא עקב תנועת מים לתא באמצעות לחץ אוסמוטי. הטרנספורטר מוציא כחמישית מאנרגיית ה- ATP של תאי בעלי חיים. מחזור הפעולה מתרחש כדלקמן:
- משאבה קושרת ATP ואחריה קישור של 3 יוני Na+ מציטופלזמה של התא
- הידרוליזה של ATP גורמת לזרחון של שאריות אספרטט של המשאבה. ADP משוחרר
- משאבה זרחנית עוברת שינוי קונפורמטיבי כדי לחשוף יוני Na+ לחלק החיצוני של התא. יוני Na+ משתחררים.
- המשאבה קושרת 2 יוני K+ חוץ תאיים.
- משאבת דה-פוספורילטים גורמת לו לחשוף יוני K+ לציטופלזמה כאשר המשאבה חוזרת לצורה המקורית.
- משאבה קושרת 3 יוני Na+, קושרת ATP ומשחררת 2 יוני K+ כדי להפעיל מחדש את התהליך
ה- ATPase Na+/K+ מסווג כ- ATPase מסוג P. קטגוריה זו של משאבה בולטת כבעלת תוצר ביניים אספרטט זרחני והיא קיימת ברחבי הממלכות הביולוגיות - חיידקים, ארכאים ואיקריוטים.
סוגי ATPase
ל-ATPases יש תפקידים בסינתזה או בהידרוליזה של ATP ומגיעים בכמה צורות שונות.
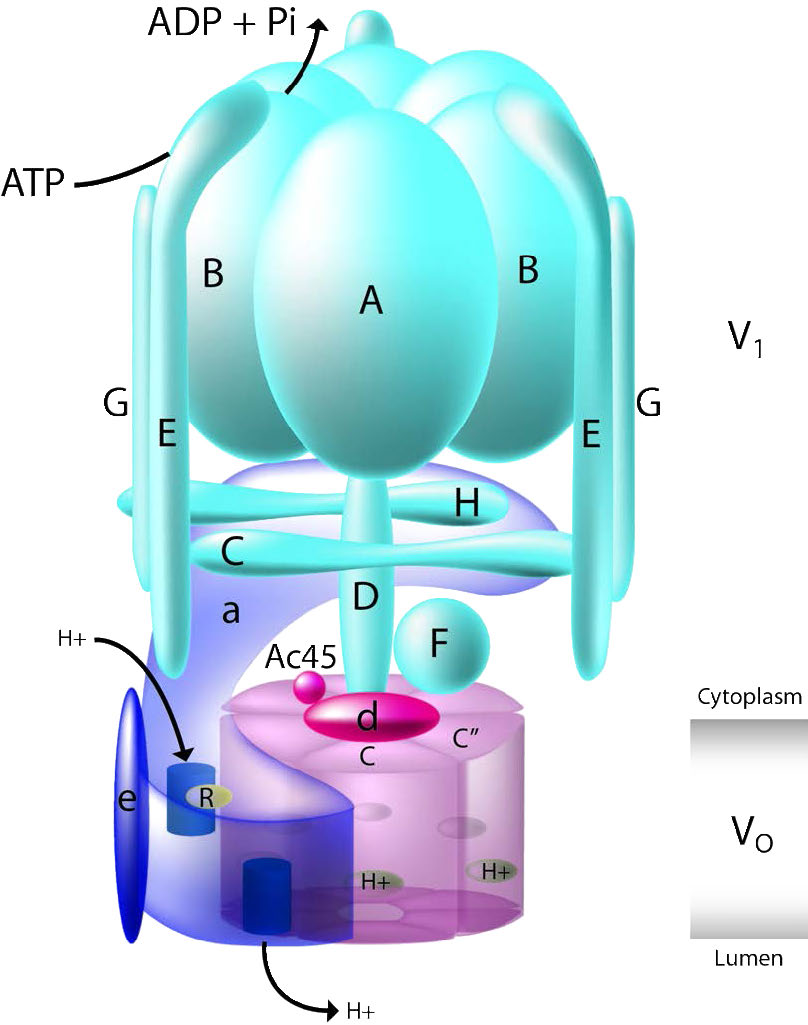
- F-ATPases (F1FO-ATPases) קיימים במיטוכונדריה, כלורופלסטים וממברנות פלזמה חיידקיות והם הסינתיסייזרים העיקריים של ATP למערכות אלו. כל אחד מהם משתמש בשיפוע פרוטונים כמקור האנרגיה שלו לייצור ATP. קומפלקס V של המיטוכונדריון הוא ATPase מסוג F.
- V-ATPases (V1VO-ATPases) נמצאים בעיקר בוואקום של אוקריוטים. הם מנצלים אנרגיה מהידרוליזה של ATP להובלת מומסים ופרוטונים לתוך vacuoles וליזוזומים, ובכך מורידים את ערכי ה-pH שלהם.
ה- ATPases מסוג V ו- F דומים מאוד במבנה. סוג V (איור 3.39) משתמש ב-ATP כדי לשאוב פרוטונים לתוך ואקואולים וליזוזומים, בעוד שסוגי F משתמשים בשיפועי פרוטונים של המיטוכונדריה והכלורופלסטים לייצור ATP.
- A-ATPases (A1AO-ATPases) נמצאים בארכאים ודומים לתפקוד F-ATPases.
- P-ATPases (E1E2-ATPases) נמצאים בחיידקים, פטריות ובממברנות פלזמה אוקריוטיות ואברונים. הם מעבירים מגוון של יונים על פני ממברנות. לכל אחד מהם מנגנון פעולה משותף הכולל אוטופוספורילציה של שרשרת צד של חומצה אספרטית שמורה בתוכו. דוגמאות ל-ATPases מסוג P כוללות את ה-Na+/K+ ATPase ואת משאבת הסידן.
- E-ATPases הם אנזימים הנמצאים על פני התא. הם מבצעים הידרוליזה של מגוון טריפוספטים נוקלאוזידים חוץ-תאיים, כולל ATP.
העברת עצבים
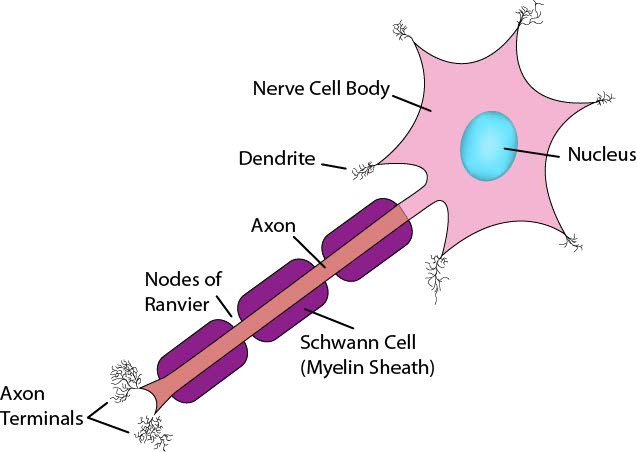
כעת, לאחר שראית כיצד Na+/K+ ATPase מתפקד, ראוי לדון כיצד תאי עצב משתמשים בשיפועי יונים שנוצרו איתו כדי ליצור ולהעביר אותות עצביים. נוירונים הם תאים של מערכת העצבים המשתמשים באותות כימיים וחשמליים כדי להעביר מידע במהירות על פני הגוף (איור 3.40). מערכת העצבים החושית מקשרת בין קולטנים לראייה, שמיעה, מגע, טעם וריח למוח לצורך תפיסה. נוירונים מוטוריים עוברים מחוט השדרה לתאי שריר. לנוירונים אלה יש גוף תא והרחבה ארוכה ודקה מאוד הנקראת אקסון, הנמתחת מגוף התא בחוט השדרה עד לשרירים שהם שולטים בהם. דחפים עצביים נעים במורד האקסון כדי לעורר התכווצות שרירים.
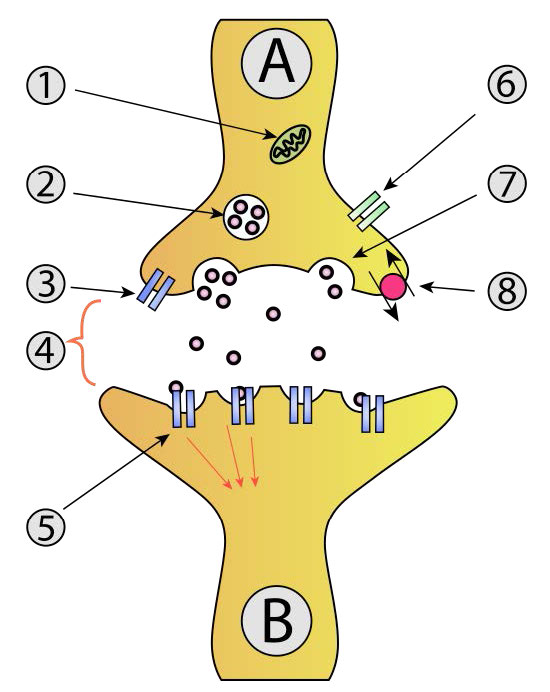
אותות עוברים דרך נוירונים, ובסופו של דבר מגיעים לצמתים עם תאי עצב אחרים או תאי מטרה כגון תאי שריר. שימו לב שנוירונים אינם יוצרים מגע פיזי זה עם זה או עם תאי שריר. החלל הזעיר בין שני נוירונים או בין נוירון לתא שריר נקרא שסע סינפטי. בשסע הסינפטי, הנוירון משחרר נוירוטרנסמיטורים היוצאים מתא העצב ועוברים על פני הצומת לתא מקבל שבו נוצרת תגובה. תגובה זו עשויה ליצור אות עצבי נוסף, אם התא הסמוך הוא תא עצב או שהוא עשוי להיות התכווצות שרירים אם הנמען הוא תא שריר (איור 3.41).
בבחינת תנועת מידע דרך תאי עצב, אם כן, נדון בשני שלבים - 1) יצירה והתפשטות של אות בתא עצב ו 2) פעולה של נוירוטרנסמיטורים היוצאים מתא עצב ועוברים צומת סינפטי.
מקור אות
יצירת אות עצבי מתחילה בגירוי לתא העצב. במקרה של התכווצות שרירים, קליפת המוח המוטורית של המוח שולחת אותות לנוירונים המוטוריים המתאימים, וממריצה אותם ליצור דחף עצבי. כיצד נוצר דחף כזה?
פוטנציאל מנוחה
במצב הלא מגורה, לכל התאים, כולל תאי עצב, יש הפרש מתח קטן (הנקרא פוטנציאל מנוחה) על פני קרום הפלזמה, הנובע משאיבה לא שוויונית של יונים על פני הממברנה. ה-Na+/K+ ATPase, למשל, שואב יוני נתרן מהתא ויוני אשלגן לתאים. מכיוון ששלושה יוני נתרן נשאבים החוצה על כל שני יוני אשלגן שנשאבים פנימה, נוצר מטען ושיפוע כימי. שיפוע המטען הוא שמוליד את פוטנציאל המנוחה.
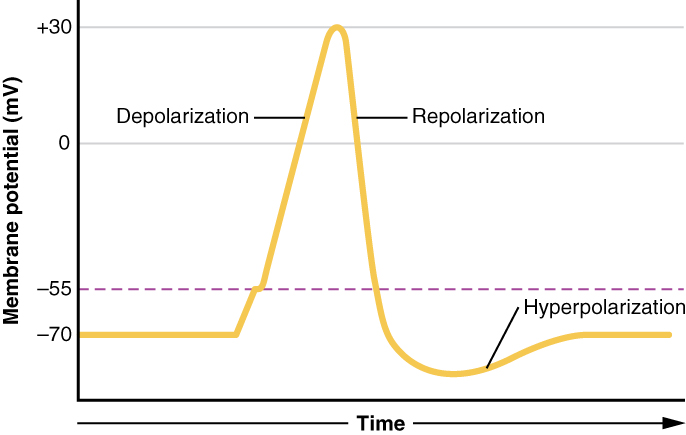
שינוי שיפועי היונים על פני הממברנות מספק את הכוח המניע לאותות עצביים. זה קורה כתוצאה מפתיחה וסגירה של תעלות יונים מגודרות. פתיחת שערים כדי לאפשר ליונים לעבור דרך הממברנה משנה במהירות את האיזון היוני על פני הממברנה וכתוצאה מכך הפרש מתח חדש הנקרא פוטנציאל הפעולה. פוטנציאל הפעולה הוא הדחף להעברת העצבים.
ייזום האות
האות שנוצר על ידי נוירון מוטורי מתחיל בפתיחת תעלות נתרן בקרום גוף תאי העצב וגורם לזרימה מהירה של יוני נתרן לתא העצב. שלב זה, הנקרא דפולריזציה (איור 3.42), מעורר אות אלקטרוכימי - פוטנציאל הפעולה. זכור כי Na+/K+ ATPase יצר שיפוע נתרן גדול, כך שיוני נתרן ממהרים לתא כאשר תעלות נתרן נפתחות. לאחר הדפולריזציה הראשונית, שערי תעלות אשלגן, המגיבים לדפולריזציה, נפתחים ומאפשרים ליוני אשלגן להתפזר במהירות אל מחוץ לתא (זכור שיוני K+ נמצאים בשפע יותר בתוך התא). שלב זה נקרא שלב הקיטוב מחדש ובמהלכו שערי הנתרן נסגרים.
היציאה המהירה של יוני אשלגן גורמת להפרש המתח "להתגבר" על פוטנציאל המנוחה ושערי האשלגן נסגרים. לאחר מכן מה שנקרא תקופת עקשן, כאשר Na+/K+ ATPase מתחיל בעבודתו לבסס מחדש את התנאים המקוריים על ידי שאיבת יוני נתרן החוצה ויוני אשלגן לתא העצב. בסופו של דבר, המערכת מתאוששת ופוטנציאל המנוחה מתבסס מחדש. הקצה היוזם של תא העצב מוכן לאחר מכן לאות נוסף.
הפצת פוטנציאל פעולה
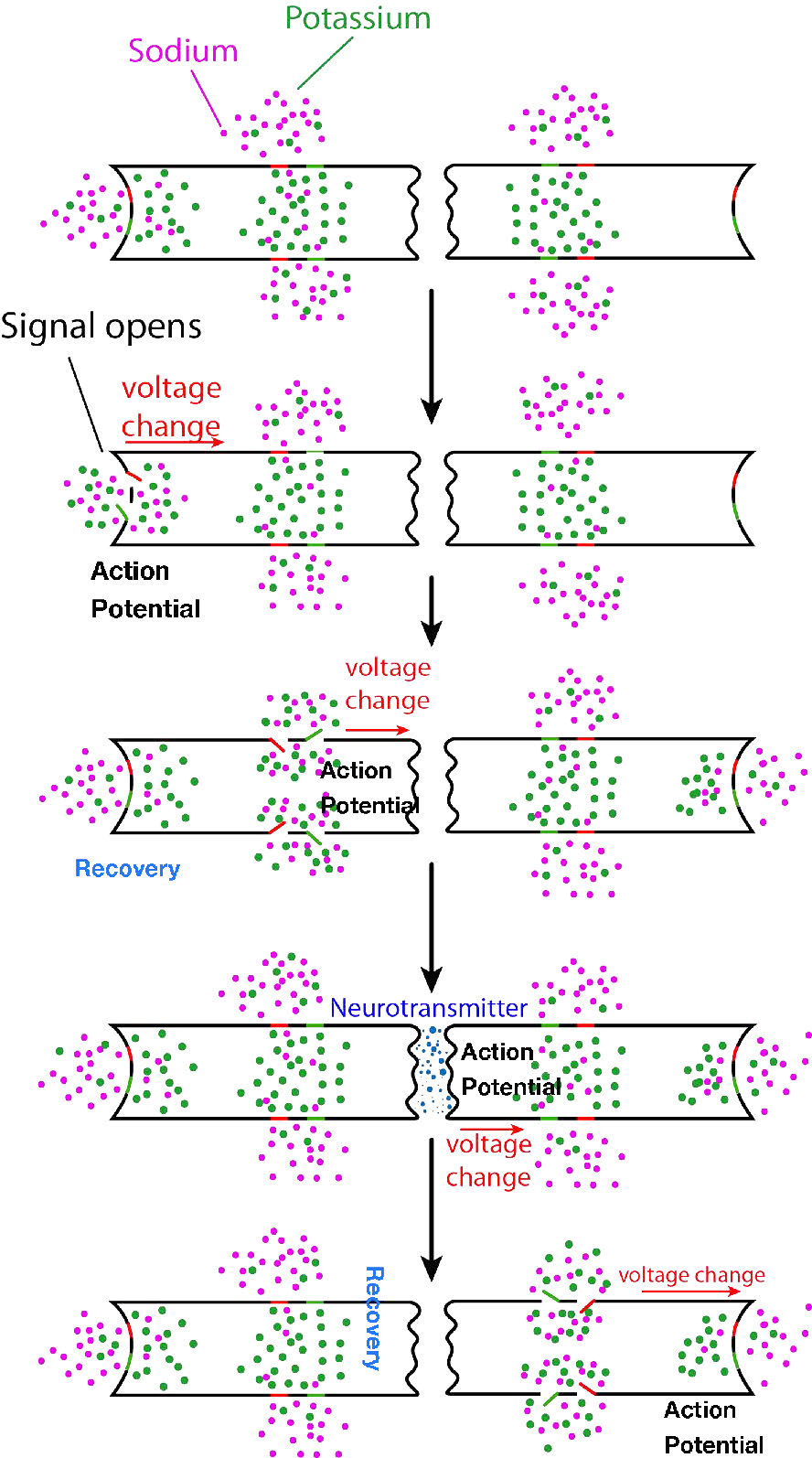
מה שתיארנו כאן הוא רק התחלת האות העצבי בחלק אחד של תא העצב. כדי שהאות יתקבל, פוטנציאל הפעולה חייב לנוע לאורך כל תא העצב (האקסון) ולגרום לשחרור אות כימי לתוך השסע הסינפטי כדי להגיע למטרה שלו. הפצת אות העצב (פוטנציאל הפעולה) בתא העצב המקורי היא הפונקציה של כל שאר תעלות היונים המגודרות (איור 3.43) הממוקמות בצידי תא העצב. שערי הנתרן והאשלגן המעורבים בהפצת האות פועלים כולם בתגובה לשינויי מתח שנוצרו על ידי השיפוע האלקטרוכימי הנע במורד תא העצב (איור 3.44). זכור כי פתיחת השערים הראשוניים בתחילת האות יצרה זרם של יוני נתרן וזרם של יוני אשלגן.
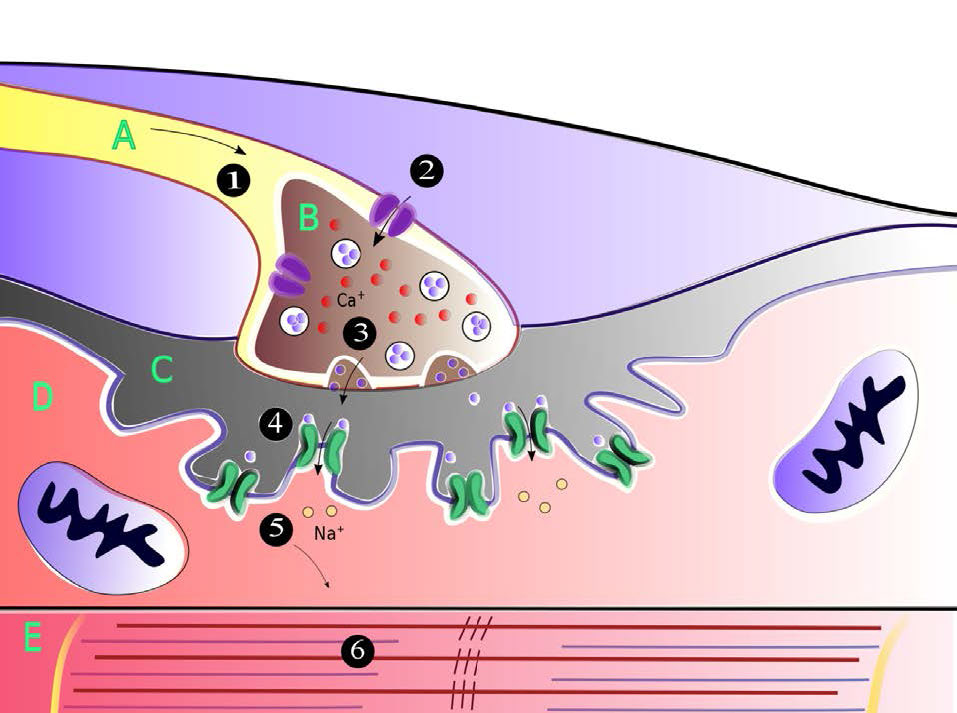
אות נע
שינוי כימי וחשמלי זה שיוצר את פוטנציאל הפעולה משאיר את קצה תא העצב בו התחיל ונוסע במורד האקסון לכיוון הקצה השני של תא העצב. לאורך הדרך הוא נתקל יותר בתעלות מגודרות נתרן ואשלגן. בכל מקרה, אלה מגיבים פשוט לשינוי המתח של פוטנציאל הפעולה ונפתחים ונסגרים, בדיוק באותו אופן שבו נפתחו השערים להפעלת האות. לפיכך, גל מהיר של יוני נתרן הולכים וגדלים ויוני אשלגן יורדים נע לאורך תא העצב, מתפשט (ומוגבר) על ידי שערים הנפתחים ונסגרים כאשר היונים והמטענים נעים במורד תא העצב. בסופו של דבר, גל הגאות היוני מגיע לקצה תא העצב (מסוף האקסון) הפונה לשסע הסינפטי.
חוצה את השסע הסינפטי
כדי שהאות יתקבל על ידי המטרה המיועדת (תא פוסט -סינפטי) מהנוירון שמקורו (נוירון פרסינפטי), עליו לחצות את השסע הסינפטי ולעורר את התא השכן (איור 3.45). העברת מידע על פני שסע סינפטי היא תפקידם של נוירוטרנסמיטורים. מדובר במולקולות קטנות המסונתזות בתאי עצב הארוזות בשלפוחית קרום הנקראות שלפוחיות סינפטיות בתא העצב. נוירוטרנסמיטורים מגיעים בכל הצורות והצורות הכימיות, מכימיקלים קטנים כמו אצטילכולין ועד פפטידים כמו נוירופפטיד Y. המוליך העצבי הנפוץ ביותר הוא גלוטמט, הפועל בלמעלה מ -90% מהסינפסות במוח האנושי.
סרט 3.2 - תנועה של פוטנציאל פעולה במורד תא עצב - ויקיפדיה
לתוך השסע
כאשר פוטנציאל הפעולה בנוירון הפרסינפטי מתקרב לקצה האקסון, שלפוחיות סינפטיות מתחילות להתמזג עם הממברנה ותכולת הנוירוטרנסמיטר שלהן נשפכת לתוך השסע הסינפטי. ברגע שהם נמצאים בשסע, הנוירוטרנסמיטורים מתפזרים, חלקם מגיעים לקולטנים בתא הפוסט-סינפטי. קשירה של הנוירוטרנסמיטר לקולטנים על קרום התא הפוסט-סינפטי מעוררת תגובה.
עבור נוירונים מוטוריים, התא הפוסט -סינפטי יהיה תא שריר, והתגובה תהיה התכווצות/הרפיה של השרירים. בשלב זה, תא העצב שמקורו עשה את עבודתו והעביר את המידע שלו למטרה המיידית שלו. אם התא הפוסט -סינפטי הוא תא עצב, התהליך חוזר על עצמו בתא זה עד שהוא מגיע ליעדו.
טרנספורטר Na+/גלוקוז
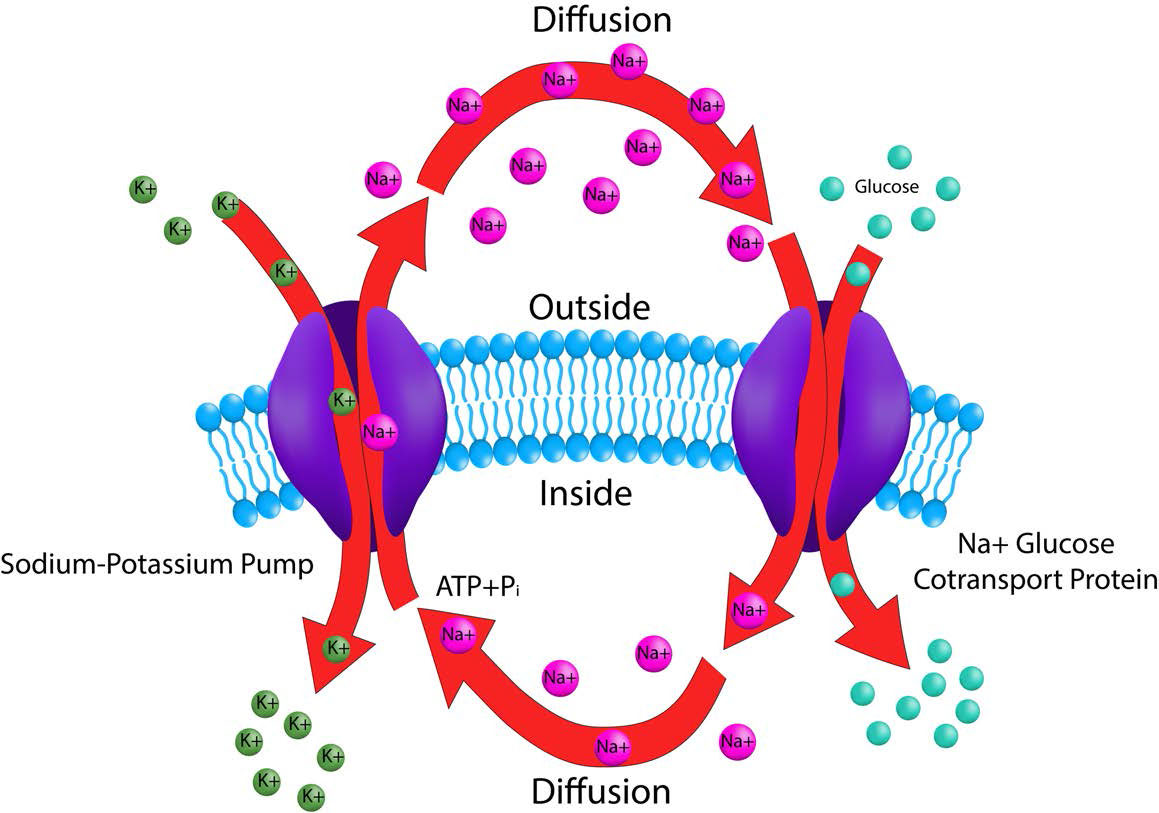
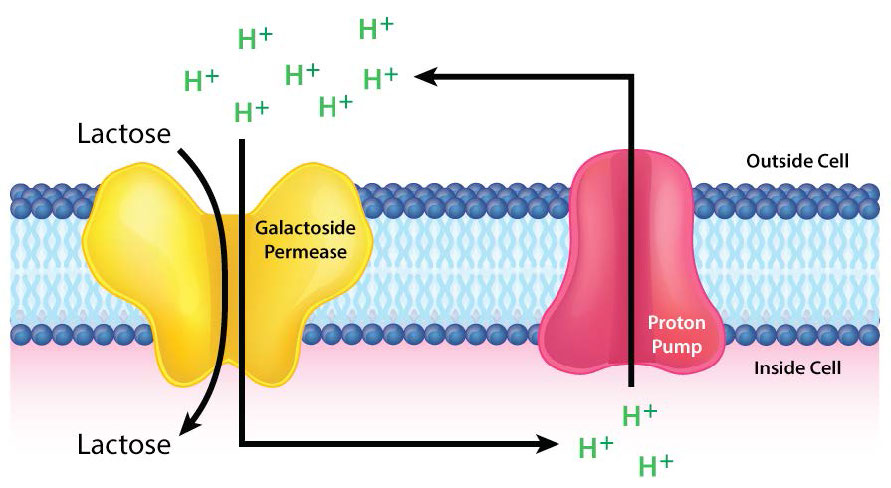
קליטת חומרים מזינים ממערכת העיכול נחוצה לחיי בעלי חיים. חלבון הובלת הנתרן/גלוקוז הוא סימפורטר אלקטרוגני המעביר גלוקוז לתאי מעיים. הוא נמצא ברירית המעי ובצינורית הפרוקסימלית של הנפרון של הכליה. מערכת הובלת הנתרן/גלוקוז מתפקדת באחרונה כדי לקדם ספיגה חוזרת של גלוקוז.
המשאבה פועלת בשילוב עם מערכת התחבורה Na+/K+. שיפוע יוני הנתרן שנבנה על ידי משאבת Na+/K+ משמש כמקור אנרגיה להנעת תנועת הגלוקוז לתאים (ראה איור 3.38). שימוש בשיפוע יונים שהוקם על ידי משאבה נפרדת ידוע בשם הובלה פעילה משנית. עבור רירית המעי, המשאבה מעבירה גלוקוז אל מחוץ למעי ולתאי המעיים. מאוחר יותר, הגלוקוז מיוצא מהצד השני של תאי המעיים לחלל הביניים לשימוש בגוף.
משאבות סידן
יוני סידן נחוצים להתכווצות שרירים וממלאים תפקידים חשובים כמולקולות איתות בתוך התאים. בנוסף, כאשר ריכוזי הסידן עולים גבוה מדי, ה- DNA בכרומוזומים יכול לזרז. לכן ריכוז הסידן בתאים מנוהל בזהירות. הוא נשמר נמוך מאוד בציטופלזמה כתוצאה מפעולה של משאבות, הן בקרום הפלזמה, השואבות סידן החוצה מהציטופלזמה והן באברונים, כגון הרשתית האנדופלזמית (רשתית סרקופלסמית של תאי שריר), השואבים סידן מתוך הציטופלזמה ואל האברונים הללו.
פתיחת תעלות סידן, אם כן, מגבירה את ריכוז הסידן במהירות בציטופלזמה וכתוצאה מכך תגובה מהירה, בין אם הכוונה היא איתות או התכווצות של שריר. לאחר יצירת התגובה, הסידן נשאב חזרה מהציטופלזמה על ידי משאבות הסידן המתאימות.
חלק ממשאבות הסידן משתמשות ב- ATP כמקור אנרגיה להעברת סידן ואחרות משתמשות בשיפועי יונים, כגון נתרן לאותה מטרה.
טרנספורטר Na+/Ca++
משאבת סידן אחת מעניינת משתמשת בשיפוע הנתרן כמקור אנרגיה. זוהי משאבת הנתרן/סידן. מערכת אנטי-פורט אלקטרוגנית זו משתמשת בתנועת הנתרן לתא ככוח מניע להעברת סידן מהתא, אם כי כיוונו יכול להתהפך בנסיבות מסוימות. המשאבה היא מערכת בעלת קיבולת גבוהה להעברת הרבה סידן במהירות, נעה עד 5000 יוני סידן בשנייה ונמצאת ברקמות רבות עם פונקציות רבות.
דיגיטליס
פונקציה חשובה אחת של משאבת Na+/Ca++ מתרחשת בתאי לב. Ca++ חשוב להתכווצות שריר הלב. זרימת סידן מהתאים היא הפעולה הרגילה של המשאבה, אולם במהלך עליית המחזור ישנה תנועה גדולה של יוני נתרן לתא הלב. כאשר זה קורה, המשאבה מתהפכת ושואבת Na+ החוצה ו- Ca++ פנימה בקצרה. מכיוון שסידן מסייע בהמרצת התכווצות שריר הלב, הדבר יכול לעזור לחזק את פעימות הלב והוא מוקד השימוש בדיגיטליס לטיפול באי ספיקת לב.
Digitalis חוסם את ATPase הנתרן-אשלגן ומפריע לשיפוע יון הנתרן. כפי שצוין לעיל, כאשר שיפוע Na+ מכוון בכיוון הלא נכון, סידן נשאב פנימה. Digitalis משמש אפוא לטיפול באי ספיקת לב מכיוון שהוא מגביר את ריכוז הסידן בתאי הלב, ומעדיף פעימות חזקות יותר.
מובילי ABC
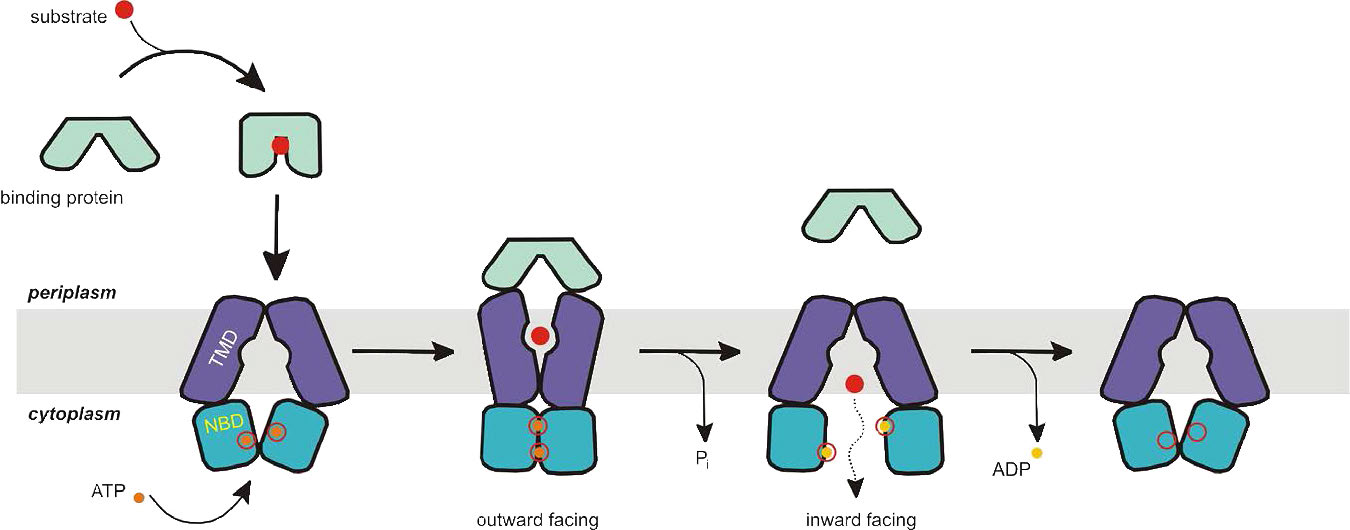
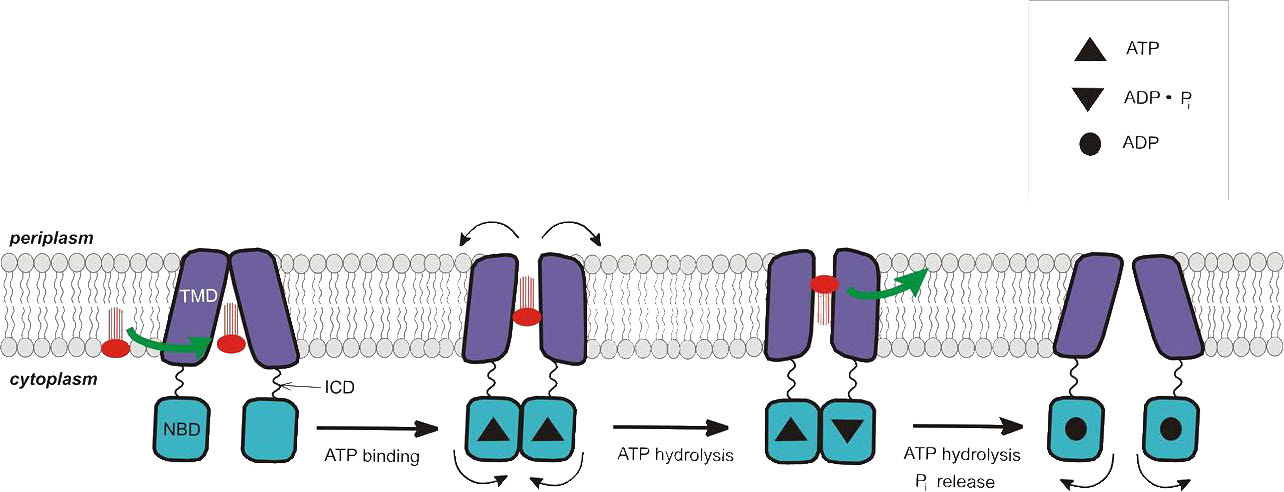
טרנספורטרים של ABC הם סוג נוסף של חלבונים טרנסממברניים המשתמשים באנרגיית ATP כדי להעביר דברים כנגד שיפועי ריכוז (איורים 3.47 & 3.48). משפחת העל החלבונית הזו כוללת מאות חלבונים (48 בבני אדם בלבד) ומשתרעת על כל הפילות הקיימות מפרוקריוטים לבני אדם. חלבונים אלה מתפקדים לא רק בהובלת ממברנה, אלא גם בתהליכים הכוללים תיקון DNA ותהליך התרגום.
תחבורה
חומרים שמעברי ABC נעים על פני ממברנות כוללים מוצרים מטבוליים, שומנים, סטרולים ותרופות. טרנספורטרים של ABC מתפקדים בעמידות מרובת תרופות של תאים רבים, ומספקים עמידות לאנטיביוטיקה בחיידקים וכן עמידות לכימותרפיה בתאים גבוהים יותר על ידי ייצוא תרופות המשמשות לטיפול בשני סוגי התאים הללו.
טרנספורטרים של ABC מחולקים לשלוש קבוצות עיקריות - 1) יבואנים (פרוקריוטים בלבד); 2) יצואנים (פרוקריוטים ואיקריוטים), ו -3) לא מובילים בעלי תפקידים בתיקון ותרגום DNA. לכל חלבוני ההובלה של ABC יש ארבעה תחומי חלבון - שניים שהם ציטופלסמיים ושניים קשורים לממברנה. הם פתוחים לסירוגין לאזורים הציטופלסמיים או החוץ-תאיים (או הפריפלסמיים) וזה נשלט על ידי הידרוליזה של ATP.
מחלה
למובילי ABC יש תפקידים בסיסטיק פיברוזיס ומחלות אנושיות תורשתיות אחרות. הם מעורבים מאוד בפיתוח עמידות לתרופות מרובות על ידי קבוצה מגוונת של תאים. טרנספורטרים של ABC מספקים עמידות מרובת תרופות על ידי גירוש תרופות מהתאים. חלבון ABCB1, למשל, שואב תרופות לדיכוי גידולים מהתא. טרנספורטר ABC אחר המכונה Pgp מעביר תרכובות קטיוניות או ניטרליות אורגניות.
סיסטיק פיברוזיס
סיסטיק פיברוזיס (CF) היא הפרעה גנטית אוטוזומלית רצסיבית הנובעת ממוטציות בשני העותקים של הגן לחלבון מווסת המוליכות הטרנסממברנית של סיסטיק פיברוזיס (CFTR). מערכת טרנספורטר ABC זו, המעבירה יוני כלוריד ותיוציאנט על פני ממברנות רקמת האפיתל מפעילה את השפעתה בעיקר בריאות, אך הלבלב, הכבד, הכליות והמעי מושפעים ממנה.
פונקציה
ל- CFTR תפקידים בייצור נוזלי זיעה, ריר ועיכול. גילויי המחלה כוללים קשיי נשימה וייצור יתר של ריר בריאות. כאשר CFTR מתפקד, נוזלים אלה בדרך כלל דקים, אך כאשר הגן אינו מתפקד, הם נעשים עבים בהרבה ומהווים נקודות זיהום.
CFTR מכיל שני תחומים הידרוליזה של ATP ושני תחומים חוצים קרום תאים עם 6 סלילי α כל אחד. זה יכול להיות מופעל על ידי זרחון על ידי קינאז חלבון תלוי cAMP. קצה הקרבוקסיל של CFTR מקושר לשלד הציטו על ידי תחום PDZ.
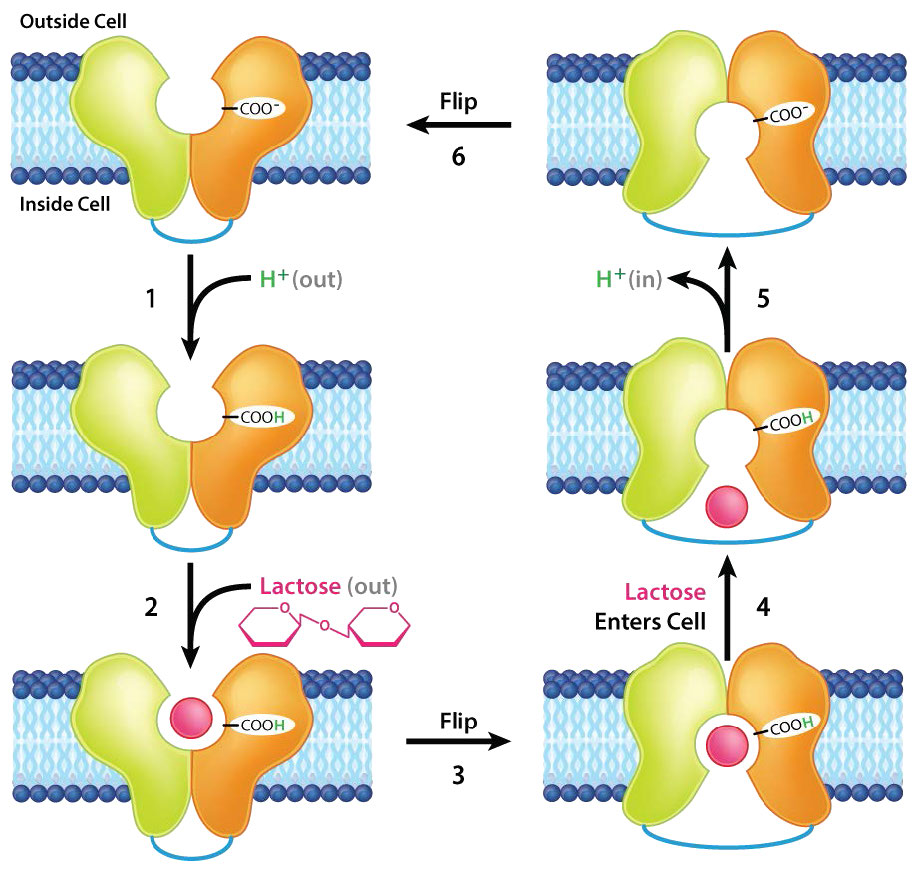
פרמיאז לקטוז
חלבון ממברנה אינטגרלי נוסף המבצע הובלה פעילה הוא פרמיאז לקטוז. זה מקל על תנועת הסוכר לקטוז על פני שכבת השומנים הדו-שכבתית של קרום התא (איורים 3.49- 3.51). מנגנון ההובלה מסווג כהובלה פעילה משנית מכיוון שהוא מנצל את שיפוע האלקטרוכימי H+ המכוון פנימה כמקור אנרגיה. כאשר לקטוז מועבר לתאים, הוא מתפרק לסוכרים החד-סוכרים המחליפים שלו - גלוקוז וגלקטוז - ליצירת אנרגיה.
האנזים המזרז תגובה זו ידוע בשם לקטאז ומחסור בו בבני אדם מוביל לאי סבילות ללקטוז (ראה כאן).
GLUTs
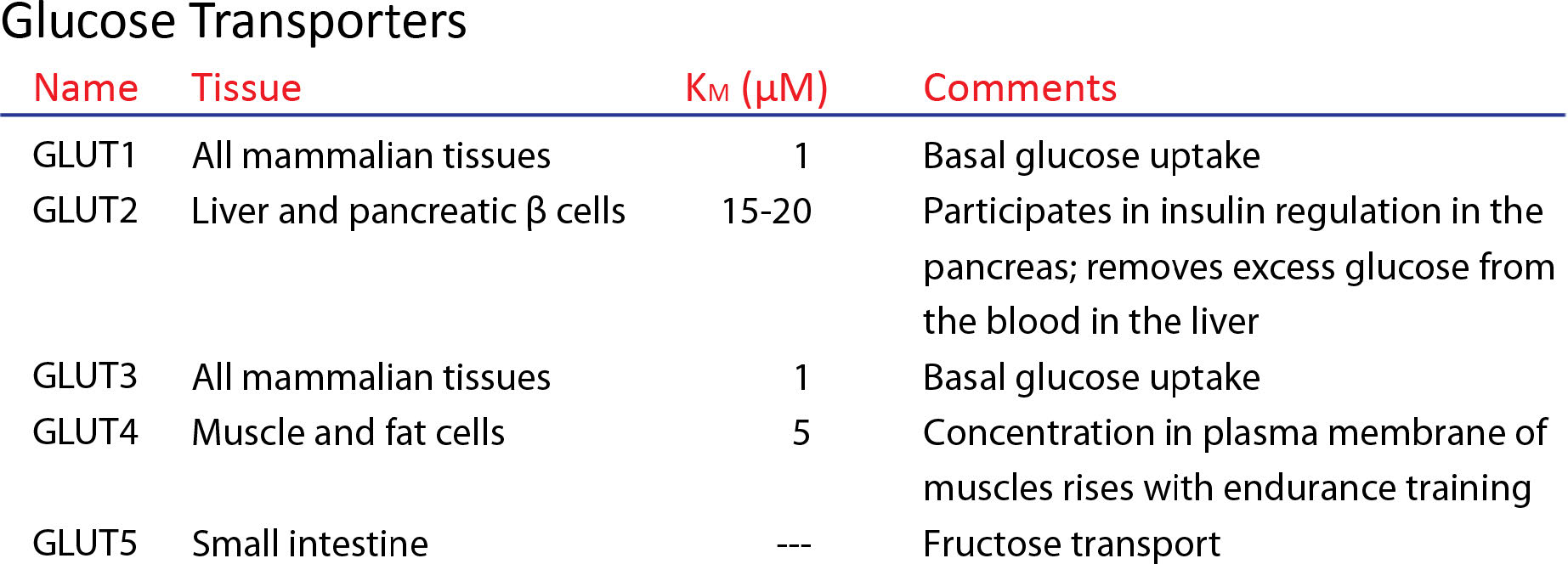
GLUTs (חלבוני הובלת גלוקוז) הם חלבוני ממברנה אינטגרליים מסוג III שאינם פורטים המשתתפים בהובלת גלוקוז על פני ממברנות לתאים. GLUTs נמצאים בכל הפילות ונמצאים בשפע בבני אדם, עם 12 גנים של GLUT. GLUT1, באריתרוציטים נלמד היטב. באמצעות GLUT 1, גלוקוז נכנס ועובר דרכו באמצעות דיפוזיה קלה בקצב הגבוה ב -50,000 מאשר בהיעדרו. GLUTs מסוגים שונים נמצאים בתאים שונים בגוף. זה בתאי הדם האדומים ידוע בשם GLUT 1 ויש לו 12 סלילים הידרופוביים המשתרעים על קרום.
למרות שהמבנה של GLUT 1 אינו ידוע, משערים כי 12 הסלילים יוצרים תא המסוגל ליצור קשרים הידרופיליים עם גלוקוז כדי להקל על מעברו.
רמות GLUT 1 באריתרוציטים עולות ככל שרמות הגלוקוז יורדות ויורדות כאשר רמות הגלוקוז יורדות. GLUT 1 יכול גם להעביר אסקורבט (ויטמין C) בנוסף לגלוקוז ביונקים (כמו בני אדם) שאינם מייצרים ויטמין C משלהם.
גלוט 4
GLUT 4 מוסדר על ידי אינסולין ונמצא בעיקר ברקמת שריר שומנית ומפוספסת. אינסולין משנה מסלולי סחר תוך תאיים בתגובה לעלייה ברמת הסוכר בדם כדי להעדיף תנועה של חלבוני GLUT שונים (כולל GLUT 4) מהשלפוחיות התוך-תאיות לקרום התא, ובכך מגרה את ספיגת הגלוקוז. GLUT 4 נמצא גם בהיפוקמפוס שבו, אם הסחר מופרע, התוצאה יכולה להיות התנהגות דיכאונית ותפקוד קוגניטיבי.
עבור כל חלבוני GLUT, מפתח לשמירה על הגלוקוז בתא הוא זרחון שלו על ידי האנזים הגליקוליזה, הקסוקינאז, בציטופלזמה. מולקולות זרחניות אינן יכולות להיכנס ל- GLUTs ואין להן אמצעי קל לצאת מהתא.
מרחק אד
למנגינה של "מר אד"
אתר מנגינות מטבוליות כאן
קורס הוא מקור,
כמובן, כמובן
מכל הידע שאנו תומכים בו
כוח מרכזי לטוב/לרע הוא מרחק הקמפוס אד
זה נכון למיקור חוץ של קורס במכללה
יש כמה סטנדרטים שיש לאכוף
את הארוכים והקצרים אנו מחזקים את מרחק הקמפוס אד
גשר
כיתה בכיתה נפגשת כל שבוע באותה שעה בכל יום
אבל המרחק אד הוא הייחודי ביותר - לוח הזמנים הגמיש שלו בסדר
קורס אלקטרוני הוא מקור, כמובן, כמובן
סיוע מקוון לדוחות מעבדה
אתה לא רשום לקורס מקוון?
ואז הירשם לזה!
"אתה תאהב את אד המרחק"
הקלטה מאת דיוויד סימונס
מילים על ידי קווין אהרן
הקלטה על ידי דוד סימונס מילים על ידי קווין אהרן
313
השעה אחת
ואהרן מדבר על
הנדרסון
והאסלבלך ו-
PKa ו-
Buffers אני צריך לדעת השיר הזה הוא עבור BB שלוש חמש הו
אני מקווה שאולי
הוא יחשוב שהדרך בה
כתבנו את התשובות שלנו לא
הייתה מטורפת
אני באמת צריכה את הקרדיט
החלקי — אז השיר
הזה הוא ל-BB three five oh
זה ממש מגניב
שזה משפר אותי
צפייה בהרצאות
בסרטי Quicktime
אני באמת צריך
ללכת ולהוריד את
הפודקאסטים האלה עבור BB שלוש חמש הו
השיר הזה עבור BB 3-5-0
לצלילי "הארץ הזאת היא הארץ שלי"
אתר מנגינות מטבוליות כאן
אני מרגיש מאני
אני בפאניקה
כדאי שאלמד את האורגני הישן
שלי יש
לו תגובות
שאני צריך לדעת השיר
הזה הוא ל-BB three five הו
אני יודע שהוא אמר את זה
בגלל זה אני מפחד
כי דילגתי על
תוספת הקרדיט של יום שישי,
טוויל רודף אותי בכיתת ז-רו הנמוכה
הזו ב- BB שלוש חמש
זה יכול להיות סטרי
או אזוטרי
שהפחמנים נעשים כל כך
אנומריים
אני היסטרי מדי
עדיף שתעזוב את זה השיר
הזה בשביל BB שלוש חמש הו
הקלטה מאת טים קרפלוס
מילים על ידי קווין אהרן
הקלטה על ידי טים Karplus מילים על ידי קווין אהרן