3.1: מושגי יסוד בממברנות
- Page ID
- 207658

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)מקור: BiochemFFA_3_1.pdf. ספר הלימוד כולו זמין בחינם מהמחברים בכתובת http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy
דו-שכבות שומנים
קרום המגן סביב התאים מכיל רכיבים רבים, כולל כולסטרול, חלבונים, גליקוליפידים, גליצרופוספוליפידים וספינגוליפידים. השניים האחרונים שבהם, כאשר הם מעורבבים במרץ עם מים, ייצרו באופן ספונטני את מה שנקרא דו שכבת שומנים (איור 3.1), המשמשת גבול מגן לתא שהוא אטום במידה רבה לתנועת רוב החומרים על פניו. למעט החריגים הבולטים של מים, פחמן דו חמצני, פחמן חד חמצני וחמצן, רוב הקוטבים/יוניים דורשים חלבוני הובלה כדי לעזור להם לנווט ביעילות על פני הדו -שכבה. התנועה המסודרת של תרכובות אלה היא קריטית כדי שהתא יוכל 1) להשיג מזון לאנרגיה; 2) חומרי ייצוא; 3) לשמור על איזון אוסמוטי; 4) ליצור שיפועים להובלה משנית; 5) לספק כוח אלקטרומוטורי לאיתות עצבי; ו 6) לאחסן אנרגיה בשיפועים אלקטרוכימיים לייצור ATP (זרחון חמצוני או פוטוסינתזה). במקרים מסוימים נדרשת אנרגיה להעברת החומרים (הובלה פעילה).
דיפוזיה מקלה
במקרים אחרים, אין צורך באנרגיה חיצונית והם נעים על ידי דיפוזיה דרך ערוצים סלולריים ספציפיים. זה מכונה דיפוזיה מקלה. לפני שנדון בתנועה של חומרים על פני ממברנות, ראוי שנדון בהרכב הממברנות התאיות. ממברנות הפלזמה נבדלות מדפנות התא הן בחומרים המרכיבים אותן והן בגמישותם. קירות התא יכוסו לקראת סוף פרק זה.
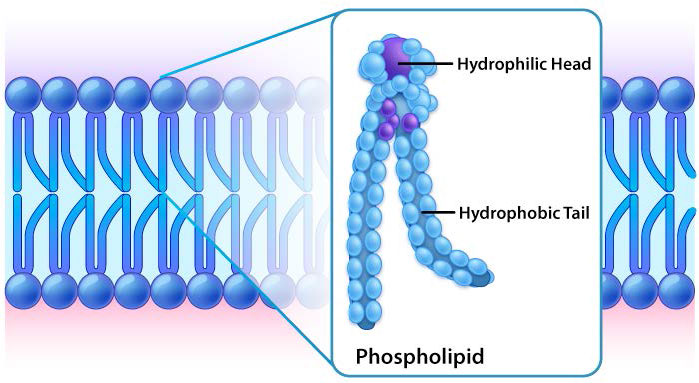
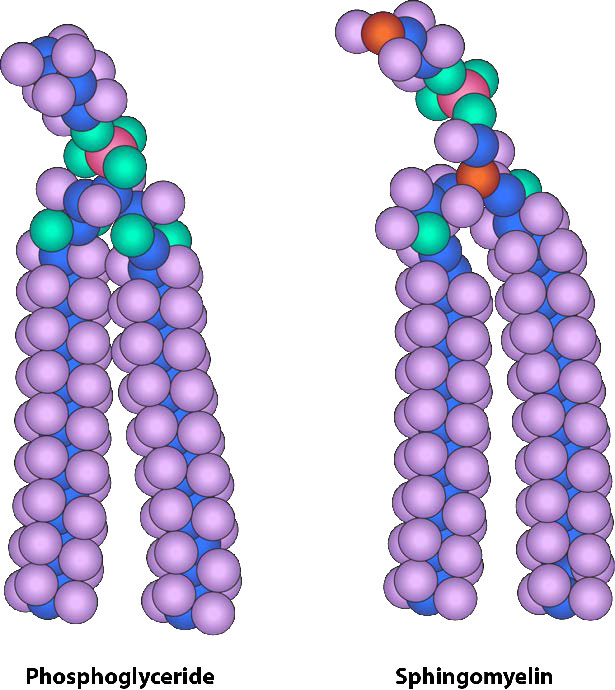
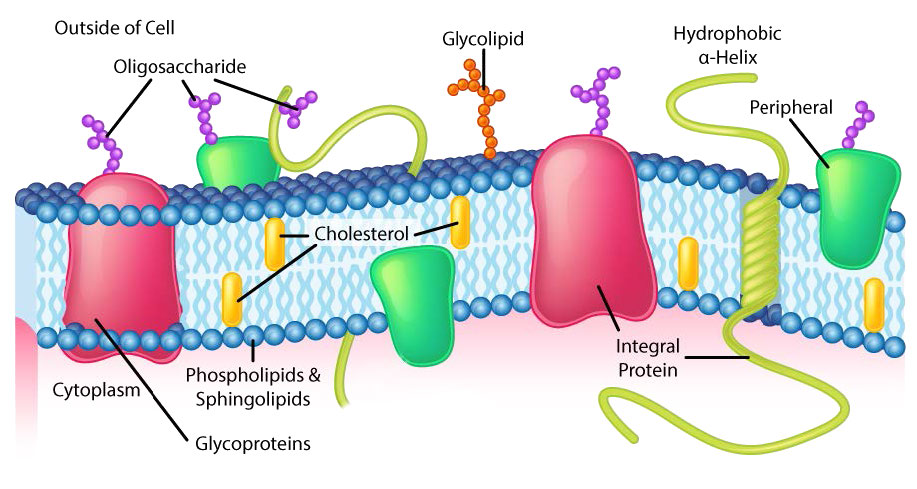
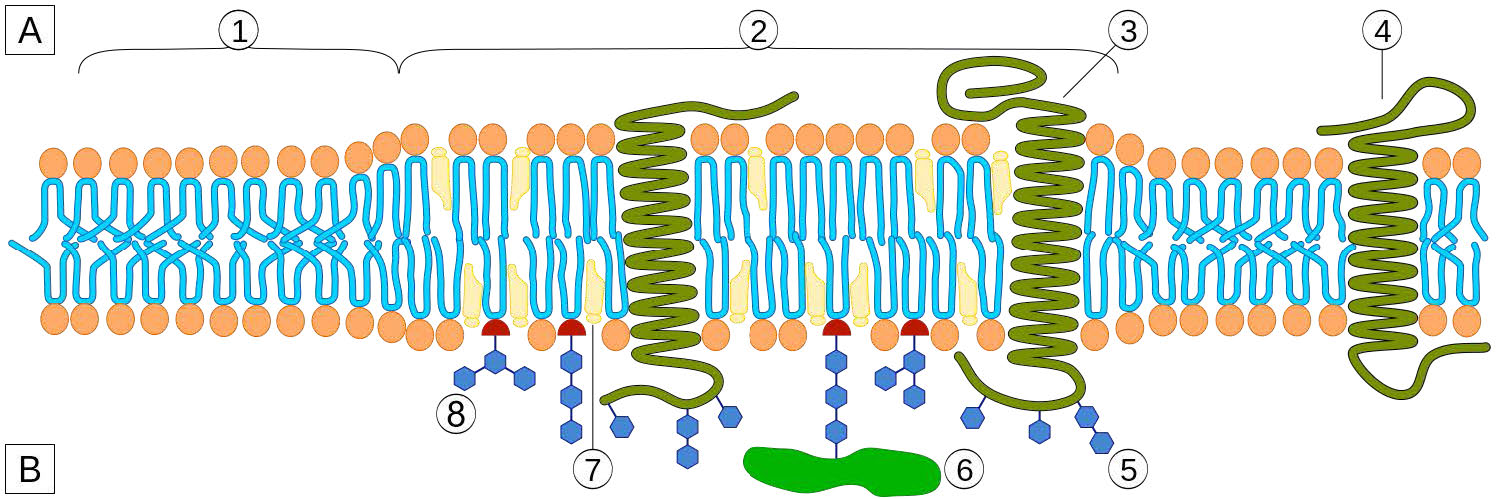
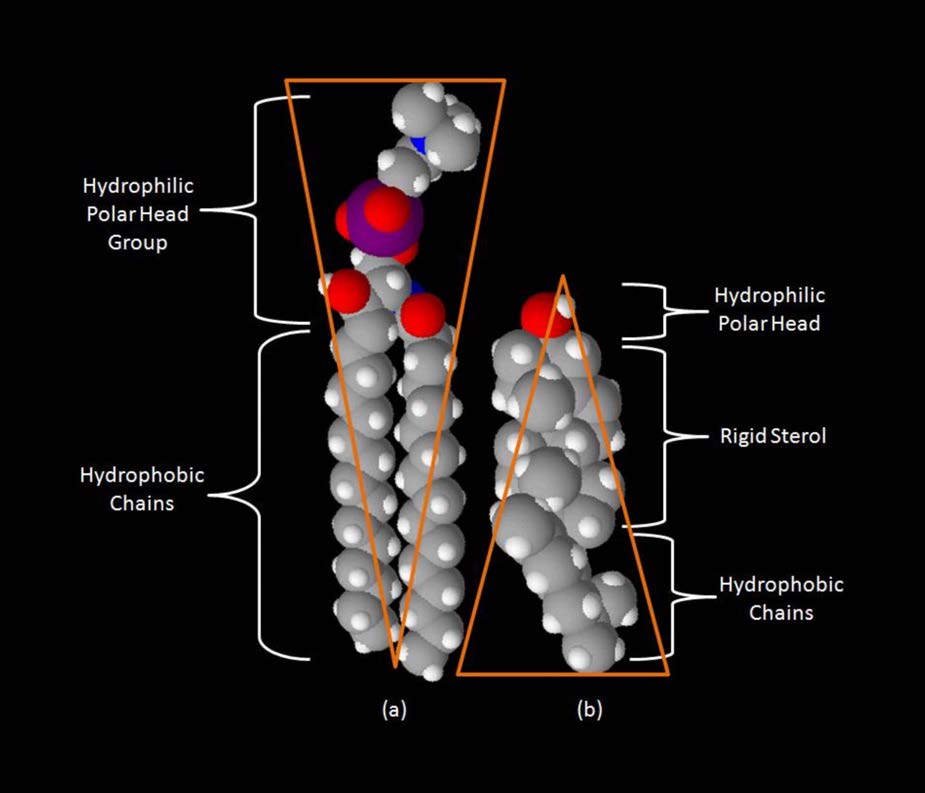
למרות שלחלק מהתאים אין דפנות תאים (תאי בעלי חיים) ולאחרים יש (חיידקים, פטריות וצמחים), יש משותף בין התאים בכך שלכולם יש ממברנות פלזמה. קיימת גם שכיחות במרכיבי הממברנות, אם כי הכמות היחסית של המרכיבים משתנה. איורים 3.1 ו -3.2 ממחישים את המבנה והסביבות של ממברנות הפלזמה. כל ממברנות הפלזמה מכילות כמות משמעותית של חומרים אמפיפיליים הקשורים לחומצות שומן. אלה כוללים את הגליצרופוספוליפידים ואת הספינגוליפידים. חומצות השומן מסומנות כזנבות הידרופוביים באיורים.
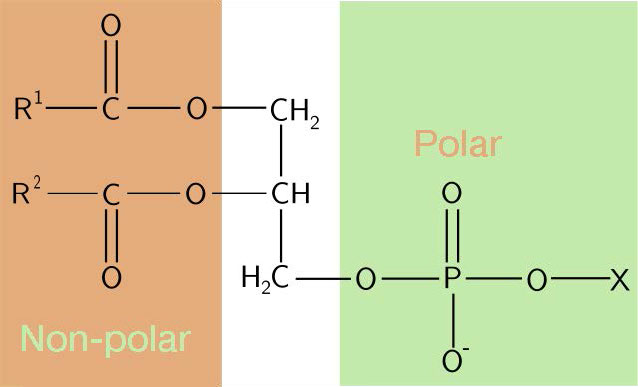
ראשים הידרופיליים
הרכב הראשים ההידרופיליים משתנה במידה ניכרת. בגליצרופוספוליפידים, פוספט תמיד קיים, כמובן, ולעתים קרובות הוא מאסטרי לחומר אחר כדי ליצור פוספטיד (איור 3.3). תרכובות נפוצות המקושרות לפוספט (במיקום X) כוללות סרין, אתנולמין וכולין. אלה משתנים במטענים שלהם כך שבדרך זו ניתן לשלוט על המטען על המשטח החיצוני או הפנימי. לתאים נוטים להיות מטענים שליליים יותר במחצית החיצונית של דו שכבת השומנים (הנקראת העלון החיצוני) ומטענים חיוביים יותר בחצי הפנימי (עלון פנימי).
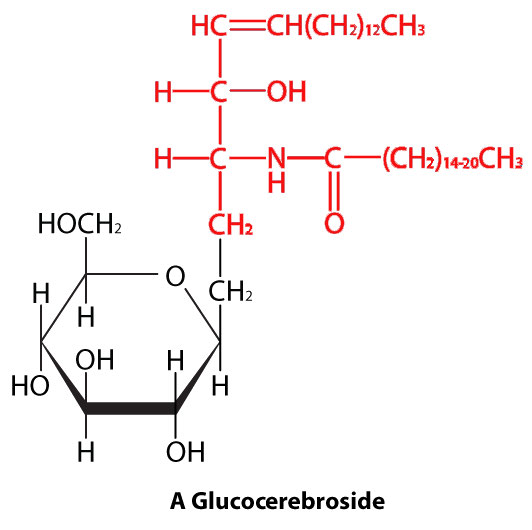
ספינגוליפידים
בספינגוליפידים (איור 3.4), הראש ההידרופילי יכול להכיל פוספט המקושר לאתנולמין או כולין וזה מתאר את מבנה הספינגומיאלין, מרכיב חשוב בממברנות עצביות. לרוב הספינגוליפידים חסר הפוספט ובמקום זאת יש להם ראש הידרופילי של סוכר בודד (cerebrosides) או אוליגוסכריד מורכב (גנגליוזידים).
הרחקת מים
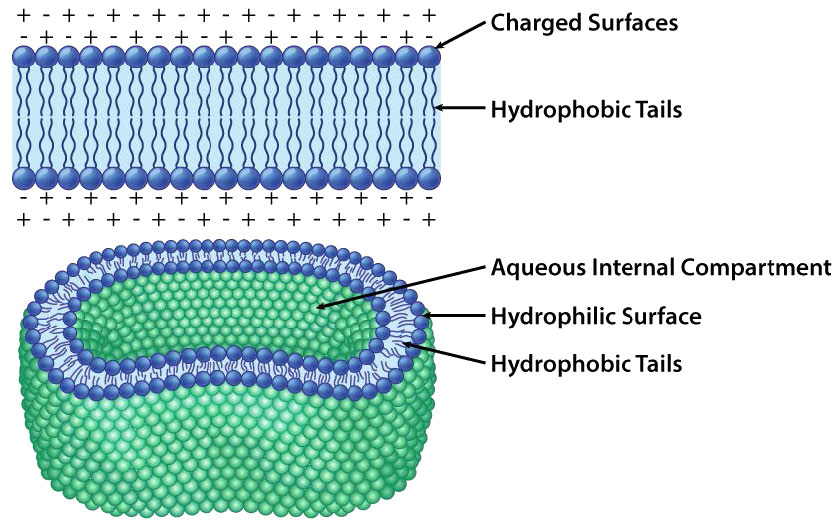
בכל מקרה, לגליצרופוספוליפיד או לספינגוליפיד יש קצה אחד שהוא קוטבי וקצה אחד שאינו קוטבי. כפי שראינו בארגון חומצות אמינו עם שרשראות צד הידרופוביות המתרחשות באופן מועדף בחלק הפנימי של חלבון מקופל כדי לא לכלול מים, כך גם החלקים הלא קוטביים של מולקולות אמפיפיליות אלה מסדרים את עצמם כדי לא לכלול מים. זכור כי הציטופלזמה של תא היא בעיקר מים והחלק החיצוני של התא נשטף בדרך כלל בשכבה מימית. לכן הגיוני לחלוטין שהחלקים הקוטביים של מולקולות הממברנה מסדרים את עצמם כפי שהם עושים - חלקים קוטביים בחוץ באינטראקציה עם מים וחלקים לא קוטביים באמצע השכבה הדו-שכבתית נמנע/לא כולל מים.
הטיית קומפוזיציה
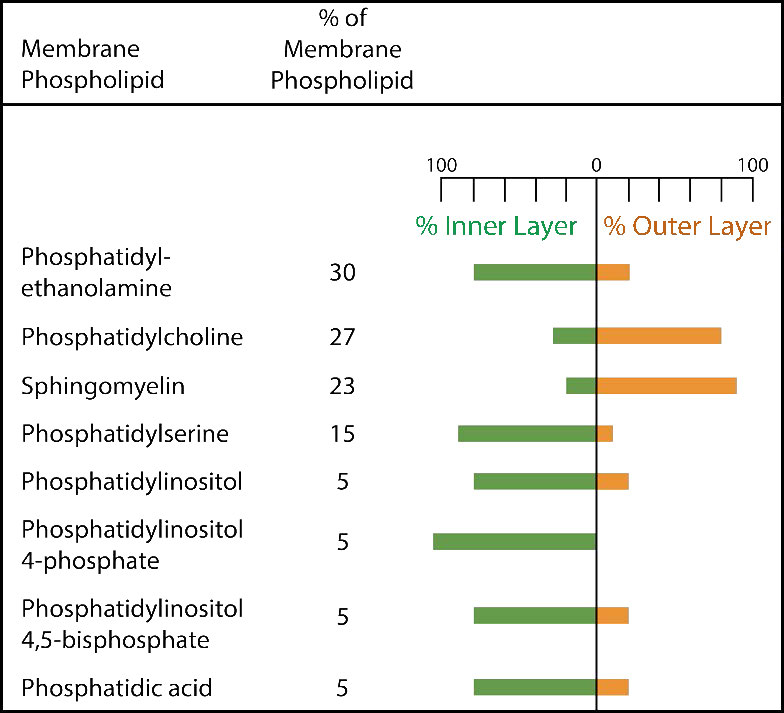
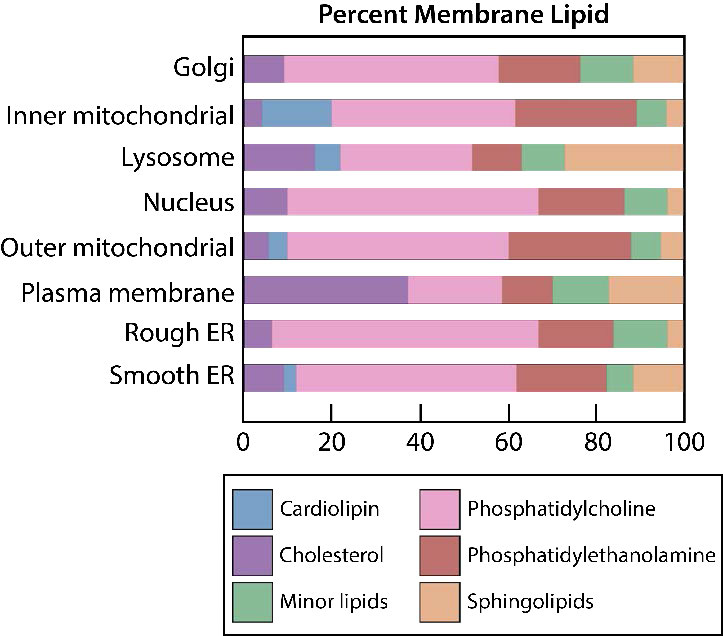
לקרום הפלזמה יש הטיות הרכב מובהקות ביחס לפנים ולחוץ (איור 3.7). ראשית, לגליקוזילציה (של שומנים וחלבונים) יש את קבוצות הסוכר הממוקמות כמעט אך ורק בחלק החיצוני של התא, הרחק מהציטופלזמה (איור 3.8). בין שומני הממברנה, ספינגוליפידים הם הרבה יותר גליקוזילטים מאשר גליצרופוספוליפידים. בנוסף, חלק מהגלירופוספוליפידים נמצאים באופן מועדף בצד זה או אחר (איור 3.7). פוספטידילסרין ופוספטידיל-אתנולמין נמצאים באופן מועדף בתוך העלון הפנימי של קרום הפלזמה, ואילו פוספטידילכולין נוטה להיות ממוקם על העלון החיצוני. בתהליך האפופטוזיס מופיעים הפוספטידילסרינים בעלון החיצוני שם הם משמשים אות למקרופאגים לקשור ולהשמיד את התא. ספינגוליפידים נמצאים באופן מועדף בקרום הפלזמה ונעדרים כמעט לחלוטין מממברנות הרשת המיטוכונדריאלית והאנדופלזמית (איור 3.9).
ממברנות אברון
הטיה בהרכב השומנים קיימת גם ביחס לממברנות האברון. הדיפוספודיגליצרוליפיד הלא שגרתי המכונה קרדיוליפין, למשל, נמצא כמעט רק בממברנות המיטוכונדריה (ראה כאן) וכמו פוספטידילסרין, תנועתו היא צעד חשוב באפופטוזיס. באיתות, פוספטידילינוזיטולים ממלאים תפקידים חשובים המספקים שליחים שניים עם ביקוע (ראה כאן).
דיפוזיה לרוחב
תנועה של שומנים בתוך כל עלון של דו שכבת השומנים מתרחשת בקלות ובמהירות עקב נזילות הממברנה. סוג זה של תנועה נקרא דיפוזיה לרוחב וניתן למדוד אותו בטכניקה הנקראת FRAP (איור 3.10, ראה גם כאן). בשיטה זו, לייזר מכה ומכתים קטע משכבת השומנים של התא, ומשאיר נקודה כפי שמוצג ב ב. עם הזמן, הכתם מתפזר בסופו של דבר על פני כל שכבת השומנים הדו-שכבתית, בדומה לטיפת דיו שתתפזר לאורך כל הדרך כאשר מוסיפים לכוס מים. מדידה של קצב הדיפוזיה נותנת אינדיקציה לנזילות הממברנה.
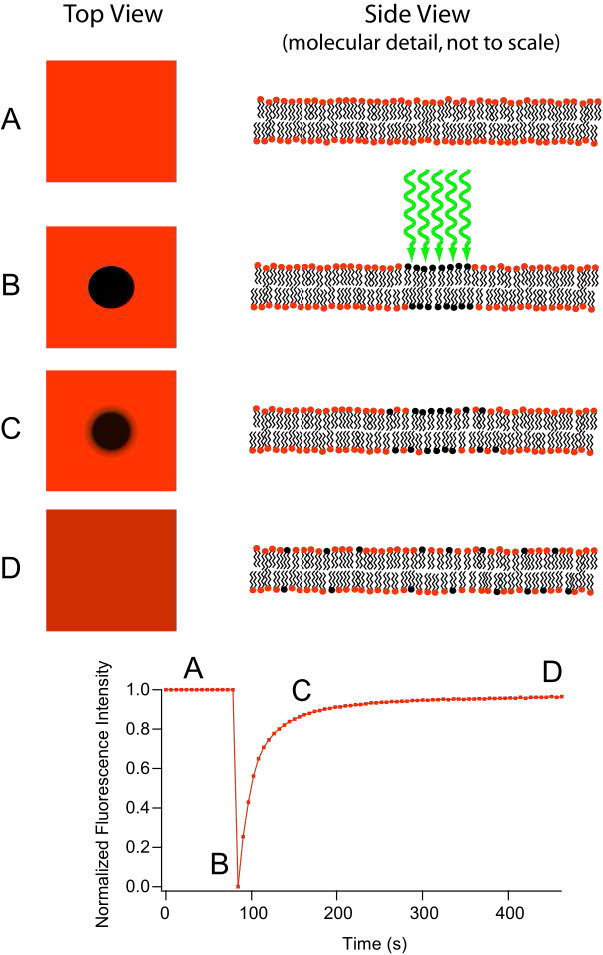
דיפוזיה רוחבית
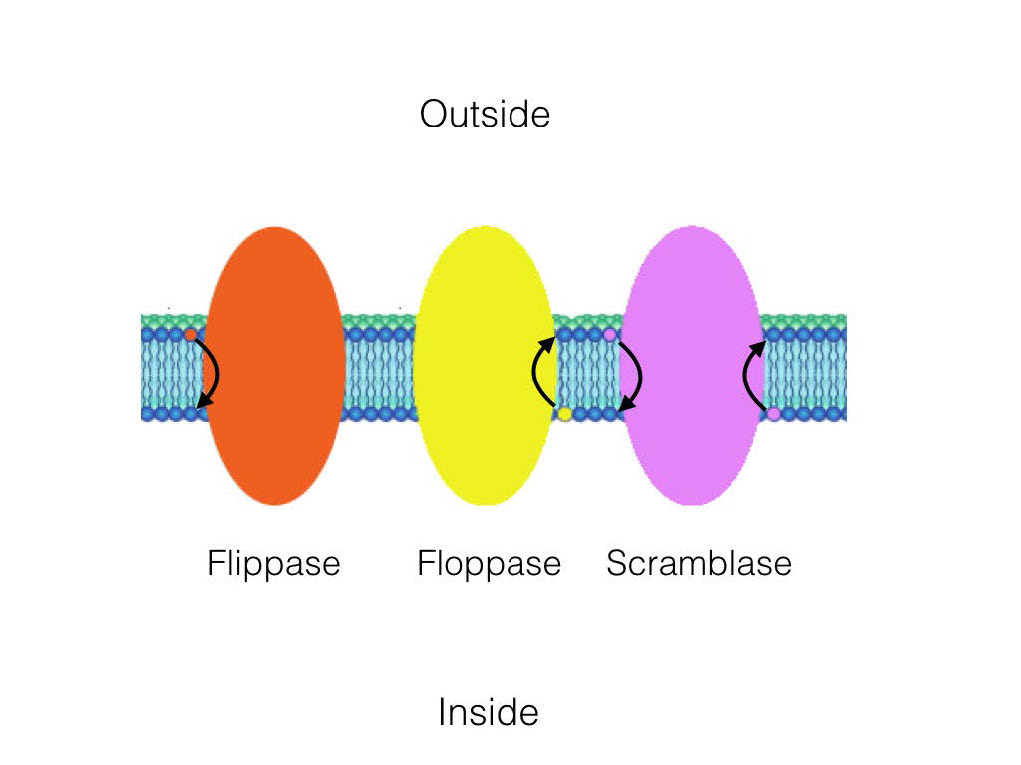
בעוד שהתנועה בדיפוזיה לרוחב מתרחשת במהירות, תנועת מולקולות מעלון אחד לעלון השני מתרחשת הרבה יותר לאט. סוג זה של תנועה מולקולרית נקרא דיפוזיה רוחבית וכמעט ואינו קיים בהיעדר פעולת אנזים. זכור כי יש הטיה של הפצה של מולקולות בין עלונים של הממברנה, כלומר משהו חייב להזיז אותם.
ישנם שלושה אנזימים המזרזים תנועה של תרכובות בדיפוזיה רוחבית. פליפאזות מעבירות גליצרופוספוליפידים/ספינגוליפידים ממברנה מהעלון החיצוני לעלון הפנימי (הצד הציטופלזמי) של התא. פלופאזות מזיזות שומנים ממברנה בכיוון ההפוך. ערבוביות נעות לשני הכיוונים.
רכיבים אחרים של דו שכבתי שומנים
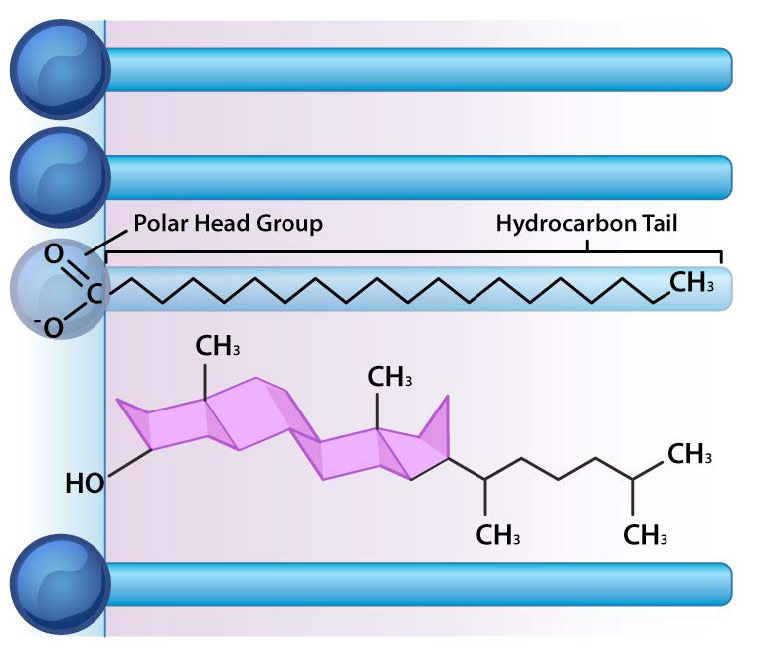
מלבד גליצרופוספוליפידים וספינגוליפידים, ישנם חומרים אחרים הנמצאים בדרך כלל בשכבות דו-שומניות של ממברנות התא. שני בולטים חשובים הם כולסטרול (איור 3.13) וחלבונים. מלבד המשמש כמבשר מטבולי של הורמונים סטרואידים וחומצות המרה, תפקידו העיקרי של הכולסטרול בתאים הוא בממברנות. השטיחות וההידרופוביות של טבעות הסטרול מאפשרות לכולסטרול לקיים אינטראקציה עם החלקים הלא קוטביים של דו שכבת השומנים בעוד שקבוצת ההידרוקסיל בקצה יכולה לקיים אינטראקציה עם החלק ההידרופילי.
נזילות ממברנה
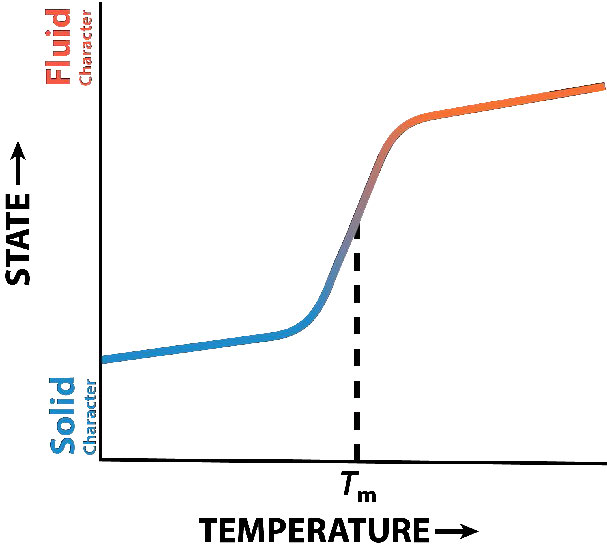
תפקוד הכולסטרול בשכבת השומנים הוא מורכב (איור 3.13). זה משפיע על נזילות הממברנה. איור 3.14 מציג את מעבר הפאזה של קרום כשהוא מחומם, ועובר מדמות "קפואה" יותר לזו של "נוזל" יותר ככל שהטמפרטורה עולה. נקודת האמצע של מעבר זה, המכונה Tm, מושפעת מהרכב חומצות השומן של התרכובות הדו-שכבתיות השומניות. חומצות שומן ארוכות ורוויות יותר יעדיפו ערכי Tm גבוהים יותר, ואילו חומצות שומן בלתי רוויות וקצרות יעדיפו ערכי Tm נמוכים יותר. מסיבה זו, לדגים, החיים בסביבות קרירות, יש רמה גבוהה יותר של חומצות שומן בלתי רוויות - לשימוש לייצור ליפידים ממברנה שיישארו נוזלים בטמפרטורות האוקיאנוס. מעניין לציין כי כולסטרול אינו משנה את ערך ה- Tm, אלא מרחיב את טווח המעבר בין צורות קפואות ונוזליות של הממברנה, ומאפשר לו טווח נזילות רחב יותר.
רפסודות ליפידים
כולסטרול נמצא בשפע גם במבני קרום הנקראים רפסודות שומנים. מתואר באיור 3.15, רפסודות שומנים הן מבנים מאורגנים בתוך הממברנה המכילים בדרך כלל מולקולות איתות וחלבוני ממברנה אינטגרליים אחרים. רפסודות שומנים משפיעות על נזילות הממברנה, העברה עצבית וסחר בקולטנים וחלבוני ממברנה.
תכונות
מאפייני ההבחנה של הרפסודות הם שהם מסודרים יותר מהדו-שכבות המקיפות אותם, המכילים יותר חומצות שומן רוויות (אריזה הדוקה יותר ופחות ארגון, כתוצאה מכך) ועד פי 5 כולסטרול. הם גם עשירים יחסית בספינגוליפידים, עם עד 50% כמויות גדולות יותר של ספינגומיאלין מאשר האזורים הסובבים את הדו-שכבה. הריכוז הגבוה יותר של כולסטרול ברפסודות עשוי לנבוע מיכולתו הגדולה יותר לקשר עם ספינגוליפידים (איור 3.16). קבוצות מסוימות, כגון חלבונים prenylated, כמו RAS, עשויות להיות מחוץ לרפסודות שומנים.
רפסודות שומנים עשויות לספק פלטפורמות ריכוז לאחר שקולטני חלבון בודדים נקשרים לליגנדים באיתות. לאחר הפעלת הקולטן מתרחשת ברפסודה שומנית, קומפלקס האיתות יספק הגנה מפני אנזימים שאינם רפסטיים שעלולים להשבית את האות. לדוגמה, תכונה נפוצה של מערכות איתות היא זרחון, כך שרפסודות שומנים עשויות לספק הגנה מפני דה-פוספורילציה על ידי אנזימים הנקראים פוספטאזות. נראה כי רפסודות שומנים מעורבות בתהליכי העברת אותות רבים, כגון איתות קולטן אנטיגן של תאי T, איתות קולטן אנטיגן של תאי B, איתות קולטן EGF, איתות אימונוגלובולין E, איתות קולטן אינסולין ואחרים. למידע נוסף על איתות, ראה כאן.
מחסום
הובלת חומרים על פני ממברנות חיונית לקיום תא. דו שכבת השומנים מהווה מחסום יעיל לכניסת רוב המולקולות וללא אמצעי לאפשר למולקולות מזון להיכנס לתא, היא תמות. המולקולות הראשוניות הנעות בחופשיות על פני דו שכבת השומנים הן קטנות ולא טעונות, כגון H2O, CO2, CO ו- O2, כך שמולקולות גדולות יותר, כמו גלוקוז, שהתא זקוק לאנרגיה, לא ייכללו ביעילות אם לא היו חלבונים כדי להקל על תנועתו על פני הממברנה.
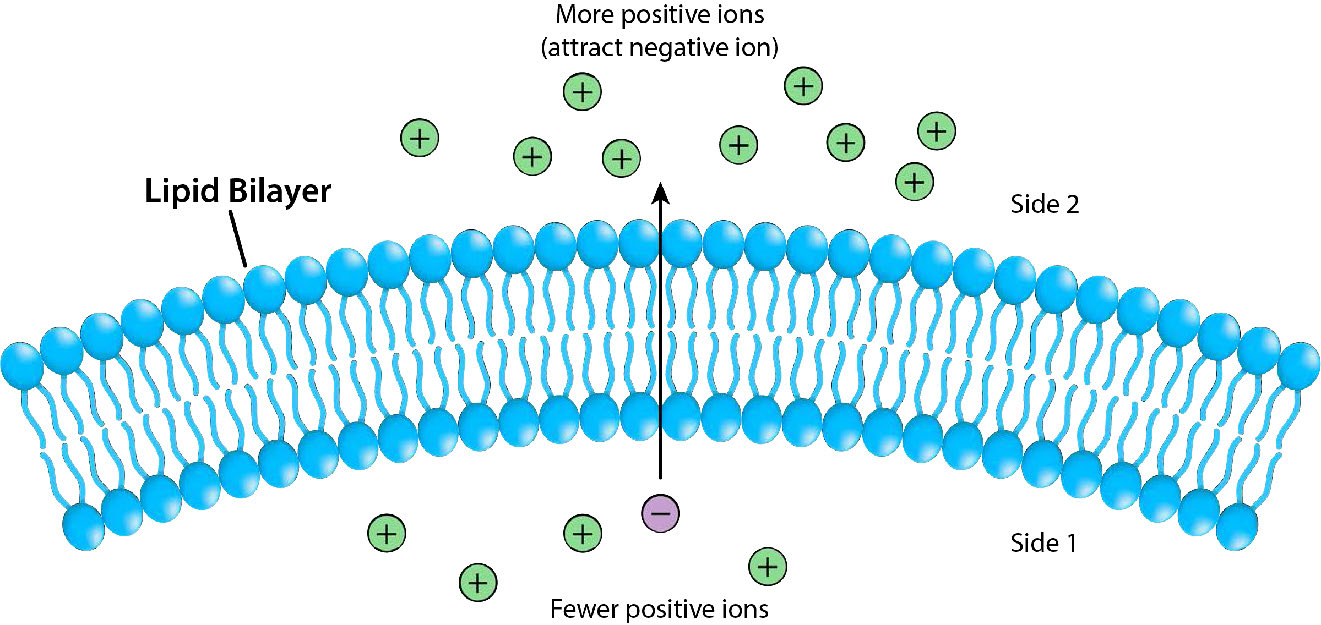
איור 3.17 מתאר את המחסום שדו-שכבת השומנים מספקת לתנועה על פניו ואת הלחצים (משיכה יונית, במקרה זה) שיכולים להשפיע על התנועה. אנרגיה פוטנציאלית מהפרשי מטען וריכוז נקצרת על ידי תאים למטרות הכוללות סינתזה של ATP, והזזת חומרים כנגד שיפוע ריכוז בתהליך הנקרא הובלה פעילה.
חלבוני ממברנה
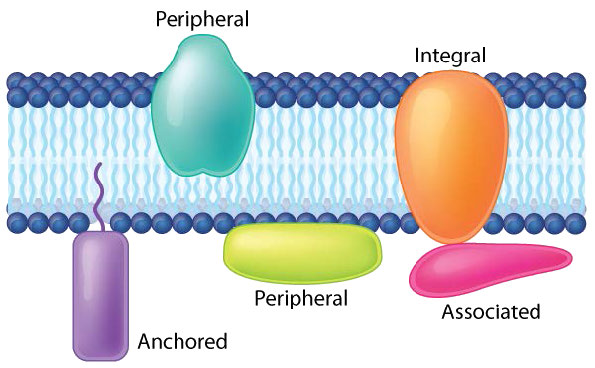
חלבונים בשכבה דו-שומנית יכולים להשתנות בכמות עצומה, בהתאם לממברנה. תכולת החלבון לפי משקל של ממברנות שונות נעה בדרך כלל בין 30 ל 75% לפי משקל. בחלק מהממברנות המיטוכונדריות יכולות להיות עד 90% חלבון. חלבונים המקושרים לממברנות וקשורים אליהם מגיעים במספר סוגים.
חלבונים טרנסממברניים
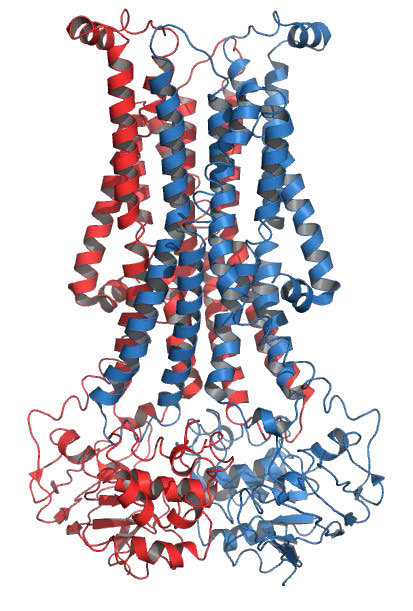
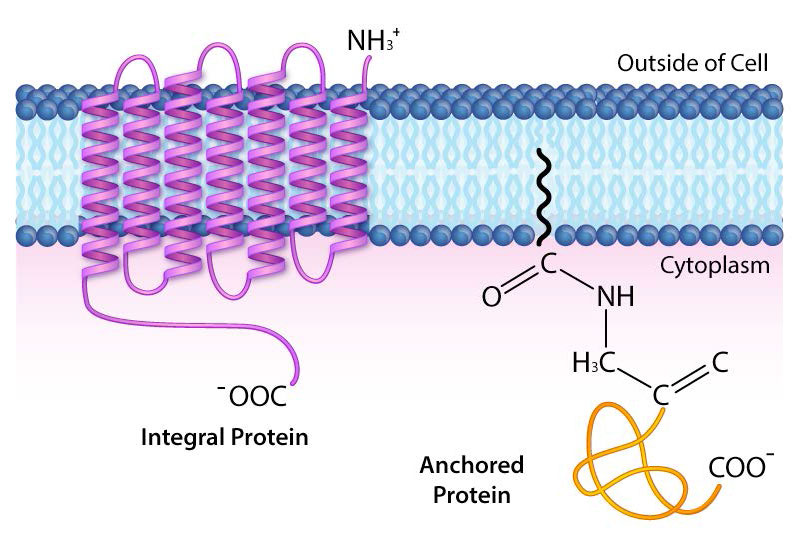
חלבונים טרנסממברניים הם חלבוני ממברנה אינטגרליים המשתרעים לחלוטין מצד אחד של קרום ביולוגי לצד השני ומוטבעים היטב בממברנה (איור 3.18). חלבונים טרנסממברניים יכולים לתפקד כאתרי עגינה להתקשרות (למטריצה החוץ -תאית, למשל), כקולטנים במערכת האיתות התאית, או להקל על הובלה ספציפית של מולקולות לתוך התא או החוצה ממנו.
דוגמה לחלבונים משולבים/טרנסממברניים כוללים את אלה המעורבים בהובלה (למשל, Na+/K+ ATPase), תעלות יונים (למשל, תעלת אשלגן של תאי עצב) והעברת אותות על פני דו שכבת השומנים (למשל, קולטנים מצמידים GProtein).
חלבוני ממברנה היקפית מקיימים אינטראקציה עם חלק מהדו-שכבה (בדרך כלל אינו כרוך באינטראקציות הידרופוביות), אך אינם מקרינים דרכו. דוגמה טובה היא פוספוליפאז A2, אשר מבקע חומצות שומן מגליצרופוספוליפידים בממברנות. לחלבוני ממברנה קשורים בדרך כלל אין אזורים הידרופוביים חיצוניים, ולכן הם אינם יכולים להטמיע בחלק משכבת השומנים, אלא נמצאים בקרבתם. קשר כזה עשוי להיווצר כתוצאה מאינטראקציה עם חלבונים או מולקולות אחרות בשכבת השומנים הדו-שכבתית. דוגמה טובה היא ריבונוקלאז.
חלבוני ממברנה מעוגנים
חלבוני ממברנה מעוגנים אינם מוטבעים בעצמם בשכבת השומנים הדו-שכבתית, אלא מחוברים למולקולה (בדרך כלל חומצת שומן) המוטמעת בממברנה (איור 3.19). משפחת החלבונים האונקוגנים המכונה ras הם דוגמאות טובות. חלבונים אלה מעוגנים לדו-שכבת השומנים על ידי התקשרות לקבוצות פרנזיל לא קוטביות המזרזות על ידי האנזים farnesyltransferase.
סיווג עדין יותר
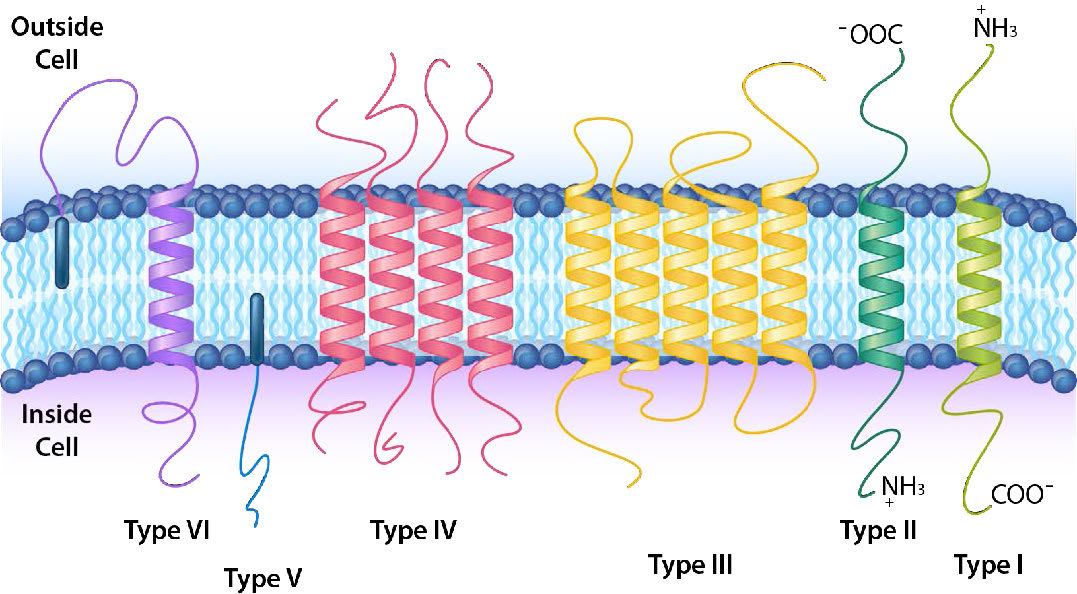
תכנית סיווג מפורטת יותר מסווגת עוד יותר את החלבונים האינטגרליים והמעוגנים לשישה סוגים שונים (איור 3.20). לסוג I ולסוג II יש רק חלק אחד מהחלבון שעובר דרך הממברנה. הם נבדלים בכיוון של קצה האמין והקרבוקסיל ביחס לבפנים/חוץ. לחלבונים טרנסממברניים מסוג I יש את קצה האמינו מבחוץ וקצה קרבוקסי מבפנים, ואילו לחלבונים מסוג II זה הפוך. חלבונים מסוג III הם שרשרת פוליפפטיד אחת שיש לה אזורים מרובים שלה חוצים הלוך ושוב על פני הממברנה, לעתים קרובות ליצירת תעלה. סוג IV הוא חלבון רב-פוליפפטיד בעל מעברים מרובים של הממברנה. לחלבונים טרנסממברניים מסוג V אין חלק מהם שחוצה את הממברנה, אך הם מעוגנים לממברנה על ידי ליפיד (כגון חומצת שומן) המוטמע בשכבה השומנית. לחלבונים טרנסממברניים מסוג VI יש לשניהם חלק מהם שחוצה את הממברנה והם מחוברים לשומנים המוטבעים בשכבת השומנים הדו-שכבתית.
סוגי דם
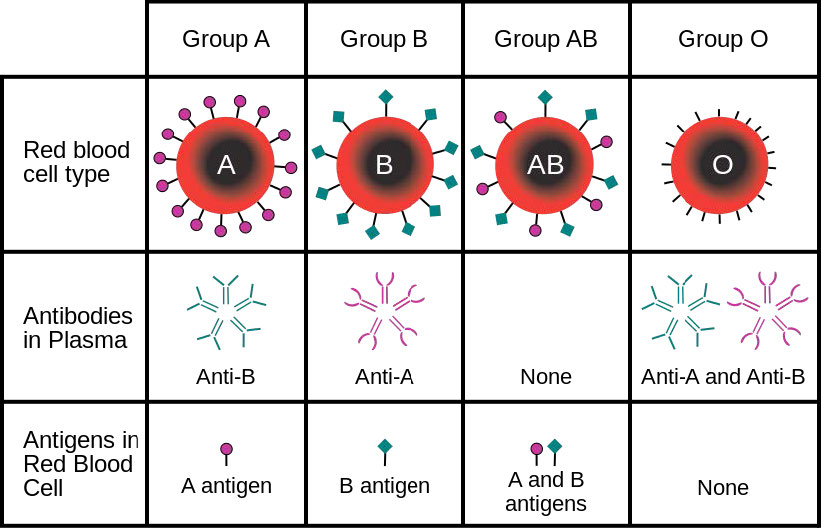
לתאים יש מאות אלפי חלבוני ממברנה והרכב החלבון של הממברנה משתנה בהתאם לתפקודו ומיקומו. גליקופרוטאינים המוטבעים בממברנות ממלאים תפקידים חשובים בזיהוי הסלולר. סוגי הדם, למשל, נבדלים זה מזה במבנה שרשראות הפחמימות היוצאות מעל פני הגליקופרוטאין בממברנות שלהן (איור 3.21).
לחץ אוסמוטי
ממברנות מספקות מחסומים/גבולות לרוב המולקולות, אך חדירות המים על פני דו שכבה שומנית יוצרת משתנה שיש לקחת בחשבון. המשתנה כאן הוא לחץ אוסמוטי. לחץ אוסמוטי (באופן רופף) מתייחס לנטייה של תמיסה לקלוט מים בתהליך האוסמוזה. באיור 3.22 ניתן לראות ייצוג חזותי של מושג הלחץ.
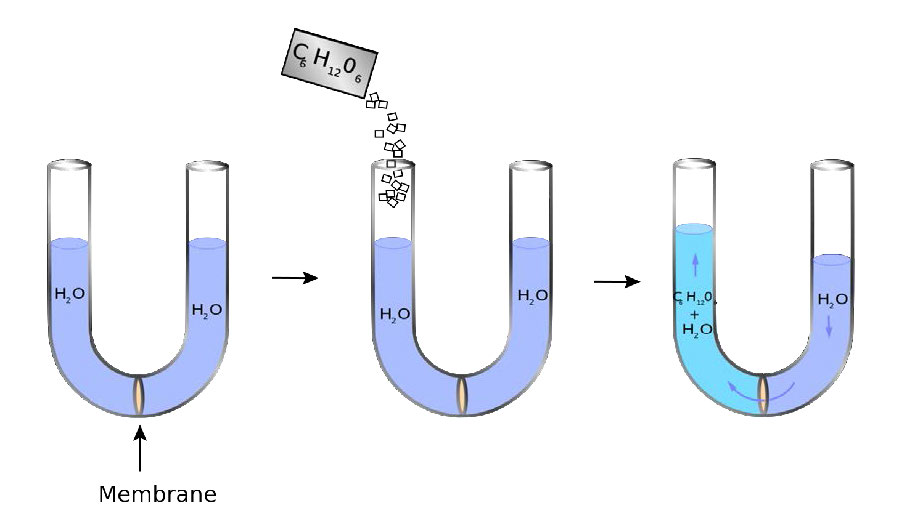
צינור בצורת U יש בתחתית קרום חדיר למחצה. מים יכולים לעבור דרך הממברנה, אך מולקולות סוכר (C6ח12O6) אינן יכולות. בצד שמאל מוסיפים סוכר ויוצרים הפרש ריכוז בין החדרים הימניים והשמאליים. מים מתפזרים על פני הממברנה מימין לשמאל בניסיון להשוות את הריכוזים, מה שגורם לרמת הצד הימני לרדת ולצד שמאל לעלות. הלחץ הנובע מהבדלי הגובה מורגש בקרום.
השוואת ריכוזים
הנוזל מימין אינו זז לגמרי שמאלה, כפי שניתן לצפות אם הכוח היחיד המעורב הוא השוואת ריכוז הסוכר על פני הממברנה (ללא סוכר מימין = ללא מים). במקום זאת, מגיעים לשיווי משקל של מיני מפלסי מים למרות שהריכוזים אינם שווים. הלחץ הקיים בממברנה אז מהבדלי הרמה תואם את הלחץ האוסמוטי של התערובת. הלחץ האוסמוטי של תמיסה הוא הפרש הלחץ הדרוש כדי לעצור את זרימת הממס על פני קרום חדיר למחצה. ניתן לחשוב על לחץ אוסמוטי גם כלחץ הנדרש להתמודדות עם אוסמוזה. המחיר האוסמוטי- איור 3.21 - סוגי דם נובעים מגליקופרוטאינים משטח התא איור 3.22 - לחץ אוסמוטי. מים מתפזרים שמאלה כדי לנסות להשוות את ריכוז המומסים. הלחץ המתממש בממברנה באיור הנכון הוא הלחץ האוסמוטי הבטוח של תמיסה מדוללת מתנהג מתמטית כמו חוק הגז האידיאלי
\[P_{osmotic} = \dfrac{nRT}{V}\]
כאשר n הוא מספר השומות, R הוא קבוע הגז, T הוא הטמפרטורה בקלווין, ו- V הוא הנפח.
זה נוח יותר בפתרונות לעבודה עם מולאריות, כך
\[P_{osmotic}= MR^* T\]
כאשר M הוא המולאריות של המולקולות המומסות, R* הוא קבוע הגז המתבטא ב- (L atm)/(K mol), ו- T היא הטמפרטורה. האות היוונית Π משמשת להתייחסות למונח האוסמוטי P, כך
\[Π = MR^* T\]
זכור בעת חישוב המולאריות כדי לכלול את המולאריות של כל חלקיק. לדוגמה, כאשר ממיסים סוכרוז בתמיסה, הוא אינו מתפצל לחלקיקים קטנים יותר, כך
\[Molarity_{Particles} = Molarity_{Sucrose}\]
עם זאת, עבור מלחים, כמו KOH, היוצר שני יונים בתמיסה (K+ ו- OH-),
חלקיקי מולריות = 2* מולאריות KOH.
שיקול משמעותי
לחץ אוסמוטי הוא שיקול משמעותי לתאים. קחו בחשבון את העובדה שמים יכולים לנוע בחופשיות על פני ממברנות התא, אך רוב תכולת התא, כגון חלבונים, DNA, יונים, סוכרים וכו ', אינה יכולה. שנית, ריכוז התרכובות הללו בתוך התא שונה מריכוזן מחוץ לתא. שלישית, מכיוון שמים יכולים לנוע דרך דו שכבת השומנים, הם נוטים לנוע בכיוון שיטה להשוות את המומסים.
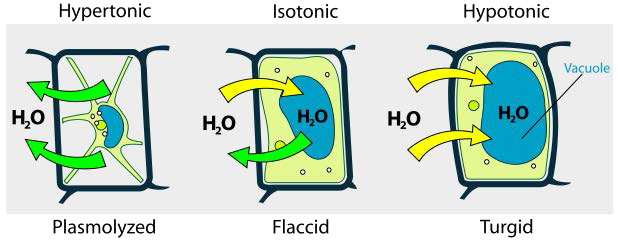
היפוטוני, היפרטוני, איזוטוני
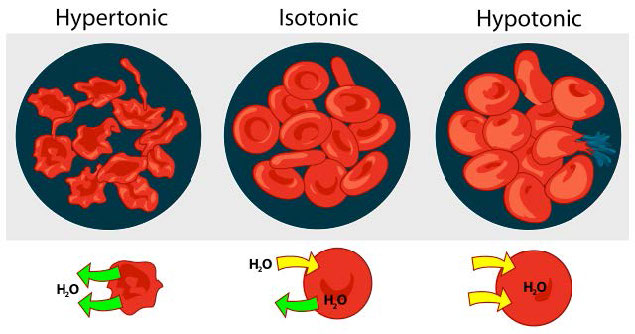
אנו שוקלים שלושה מצבים (איור 3.23). ראשית, אם ריכוז המומסים גדול יותר בתוך התא מאשר בחוץ, מים נוטים לנוע לתוך התא ולגרום לתא להתנפח. מצב זה נקרא היפוטוני. לעומת זאת, אם ריכוז המומסים גדול יותר מחוץ לתא מאשר בתוכו, מים ייצאו מהתא והתא בכיווץ. זהו מצב היפרטוני. לבסוף, אם ריכוזי המומסים לתא ומחוצה לו שווים, זה נקרא פתרון איזוטוני. כאן, לא מתרחשת תנועה של מים על פני קרום התא והתא שומר על גודלו.
אם הלחץ האוסמוטי גדול מהכוחות המחזיקים יחד קרום תאי, התא ייקרע. בגלל זה, חלק מהתאים בנו הגנות למניעת בעיות. לתאי צמחים, למשל, יש דופן תא נוקשה למדי המתנגד להתרחבות בתמיסות היפוטוניות (איור 3.24). לחיידקים יש גם דופן תא המספקת הגנה.