
1.3: מבוא - מים ומאגרים
- Page ID
- 207814

מקור: BiochemFFA_1_3.pdf. ספר הלימוד כולו זמין בחינם מהמחברים בכתובת http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy

כשזה מגיע למים, אנחנו ממש טובעים בהם, מכיוון שמים הם ללא ספק המרכיב הנפוץ ביותר בכל תא. כדי להבין את החיים, אנו מתחילים את הדיון ביסודות המים, מכיוון שכל מה שקורה בתאים, אפילו תגובות הקבורות עמוק בתוך אנזימים, הרחק מהמים, מושפע מהכימיה של המים.
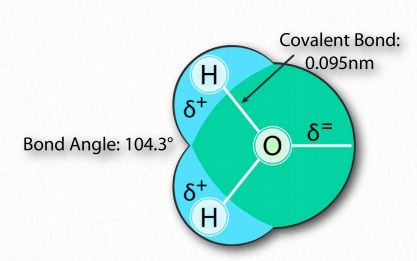
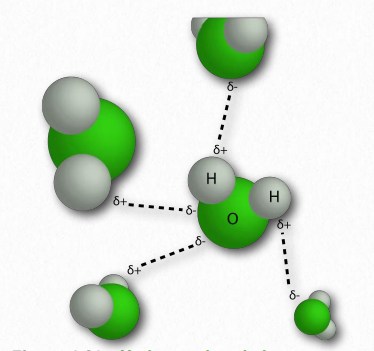
למולקולת המים צורת 'V' רחבה (זווית HO-H היא 104°) עם שיתוף לא אחיד של אלקטרונים בין החמצן לאטומי המימן (איור 1.23). חמצן, עם האלקטרושליליות הגבוהה שלו, מחזיק אלקטרונים קרוב יותר לעצמו מאשר ההידרוגנים. המימנים, כתוצאה מכך, מתוארים כבעלי מטען חיובי חלקי (בדרך כלל מוגדר כ-δ+) ולחמצן יש מטען שלילי חלקי (כתוב כ-δ-). לפיכך, מים הם מולקולה קוטבית מכיוון שמטענים מופצים סביבם בצורה לא אחידה, לא באופן סימטרי.
מים כממס
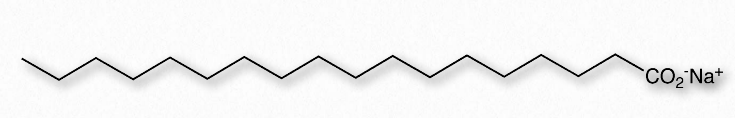
מים (איור 1.23) מתוארים כממס בגלל יכולתו להמיס (להמיס) מולקולות רבות, אך לא כולן. מולקולות שהן יוניות או קוטביות מתמוססות בקלות במים, אך חומרים לא קוטביים מתמוססים בצורה גרועה במים, אם בכלל. שמן, למשל, שאינו קוטבי, נפרד מהמים כאשר הוא מעורבב איתו. מצד שני, נתרן כלורי, המיינן, ואתנול, שהוא קוטבי, מסוגלים ליצור קשרי מימן, ולכן שניהם מתמוססים במים. המסיסות של אתנול במים חיונית עבור מבשלות בירה, ייננים ומזקקים - אך עבור נכס זה לא יהיה יין, בירה או משקאות חריפים. כפי שהוסבר בסעיף קודם, אנו משתמשים במונח הידרופילי לתיאור חומרים המקיימים אינטראקציה טובה עם מים ומתמוססים בהם ואת המונח הידרופובי כדי להתייחס לחומרים שאינם קוטביים ואינם מתמוססים במים. טבלה 1.3 ממחישה כמה חומרים קוטביים ולא קוטביים. מונח שלישי, אמפיפילי, מתייחס לתרכובות שיש להן שתי תכונות. סבונים, למשל, הם אמפיפיליים, המכילים זנב אליפטי ארוך ולא קוטבי וראש שמיינן.

טבלה 1.3 תמונה מאת אליה קים
מסיסות
מסיסות החומרים במים מבוססת על שינויים באנרגיה חופשית, כפי שנמדד על ידי ΔG. זכור, מהכימיה, ש- H הוא האנטלפיה (חום בלחץ קבוע) ו- S היא אנטרופיה. בהתחשב בכך,
\[ΔG = ΔH - TΔS\]
כאשר T היא הטמפרטורה בקלווין. כדי שתהליך יהיה חיובי, ΔG עבורו חייב להיות פחות מאפס.
מהמשוואה, ערכי ΔG מופחתים יועדפו עם ירידות באנטלפיה ו/או עלייה באנטרופיה. הבה נבחן תחילה מדוע חומרים שאינם קוטביים אינם מתמוססים במים. נוכל לדמיין מצב שבו תהליך ההמסה כרוך ב"סביבה "של כל מולקולה של המומס הלא קוטבי במים, בדיוק כמו כל נתרן וכל יון כלוריד מוקף במולקולות מים כאשר המלח מתמוסס.
ארגון מים
יש הבדל משמעותי, אם כי בין סביב מולקולה לא קוטבית עם מולקולות מים לבין יונים מסביב (או תרכובות קוטביות) עם מולקולות מים.
ההבדל הוא שמכיוון שמולקולות לא קוטביות לא ממש מקיימות אינטראקציה עם מים, המים מתנהגים בצורה שונה מאוד מאשר עם יונים או מולקולות היוצרות קשרי מימן. למעשה, סביב כל מולקולה לא קוטבית, המים מתארגנים מאוד, ומיישרים את עצמם באופן קבוע. כפי שכנראה כל סטודנט לכימיה א' זוכר, אנטרופיה היא מדד לאי סדר, כך שכאשר משהו הופך לסדר, האנטרופיה פוחתת, כלומר ΔS שלילי, כך שהמונח TΔS במשוואה הוא חיובי (שלילי של שלילי).
מכיוון שלערבוב של חומר לא קוטבי עם מים אין בדרך כלל מרכיב חום משמעותי, ה- ΔG חיובי. משמעות הדבר היא, אם כן, כי המסת תרכובת לא קוטבית במים אינה חיובית ואינה מתרחשת במידה משמעותית. יתר על כן, כאשר החומר הלא קוטבי מתחבר לעצמו ולא למים, אז מולקולות המים חופשיות להתערבב, מבלי להיות מסודרות, וכתוצאה מכך עלייה באנטרופיה. אנטרופיה מניעה אפוא את ההפרדה של חומרים לא קוטביים מתמיסות מימיות.

חומרים אמפיפיליים
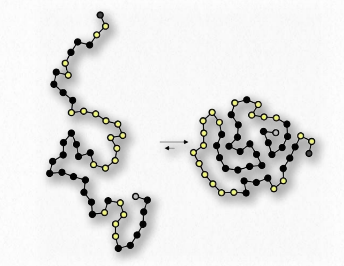
לאחר מכן, אנו שוקלים ערבוב של חומר אמפיפילי, כגון סבון, עם מים (איור 1.24). יוני הנתרן המחוברים לחומצות השומן בסבון יורדים בקלות בתמיסה מימית, ומשאירים אחריהם מולקולה טעונה שלילית בקצה אחד ואזור לא קוטבי בקצה השני. יינון הסבון גורם לעלייה באנטרופיה - שני חלקיקים במקום אחד. החלק הלא קוטבי של יון הסבון הטעון שלילי הוא בעייתי - אם הוא נחשף למים, הוא יגרום להתארגנות של מים ויביא לירידה באנטרופיה ולΔG חיובי.
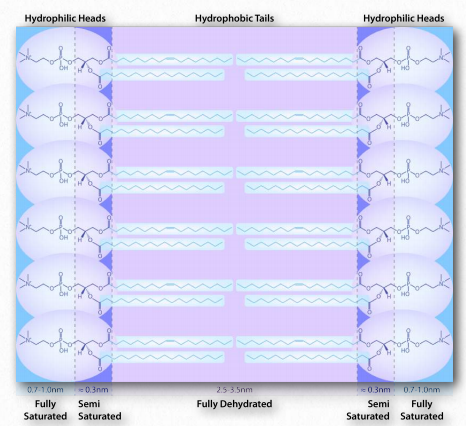
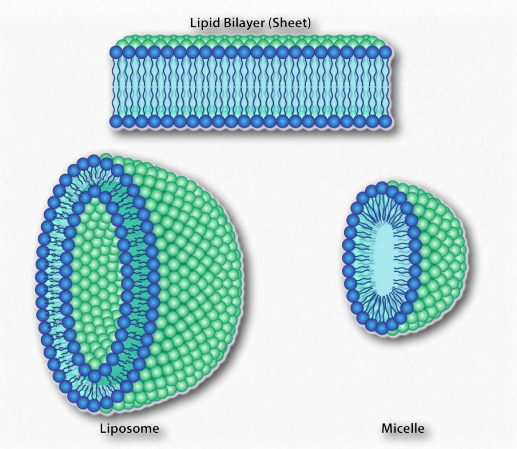
מכיוון שאנו יודעים שחומצות שומן מתמוססות במים, חייב להיות משהו אחר במשחק. יש. בדיוק כמו המולקולות הלא קוטביות בדוגמה הראשונה הקשורות זו לזו ולא למים, כך גם החלקים הלא קוטביים של יוני הסבון מתחברים זה לזה ואינם כוללים מים. התוצאה היא שיוני הסבון מסדרים את עצמם כמיצלות (איור 1.25) כאשר החלקים הלא קוטביים בחלק הפנימי של המבנה הרחק מהמים והחלקים הקוטביים מבחוץ מתקשרים עם מים.
האינטראקציה של ראשי הקוטב עם מים מחזירה את המים למצבם המופרע יותר. עלייה זו באי סדר, או אנטרופיה, מניעה את היווצרותם של מיצלות. כפי שנראה בדיון על דו שכבת השומנים, אותם כוחות מניעים גליצרופוספוליפידים וספינגוליפידים ליצור באופן ספונטני דו שכבות שבהן החלקים הלא קוטביים של המולקולות מתקשרים זה עם זה כדי לא לכלול מים והחלקים הקוטביים מסדרים את עצמם בצד החיצוני של הדו -שכבה (איור 1.28).
דוגמה נוספת נראית בקיפול חלבונים כדוריים בציטופלזמה. חומצות אמינו לא קוטביות נמצאות בחלק הפנימי של החלבון (לא כולל מים). אינטראקציה של חומצות האמינו הלא קוטביות מתגלה ככוח מניע לקיפול חלבונים שכן הם מיוצרים בתמיסה מימית.
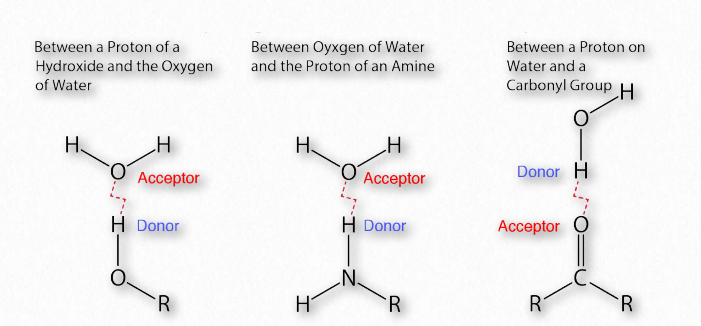
קשרי מימן
קשה להפריז בחשיבותם של קשרי מימן בביוכימיה (איור 1.30). לינוס פאולינג עצמו אמר,
".... אני מאמין שככל ששיטות הכימיה המבנית מיושמות עוד יותר על בעיות פיזיולוגיות, יימצא כי המשמעות של קשר המימן לפיזיולוגיה גדולה מזו של כל תכונה מבנית אחת אחרת".
בשנת 2011, קבוצת משימות IUPAC נתנה הגדרה מבוססת ראיות של קשר מימן הקובעת,
"קשר המימן הוא אינטראקציה אטרקטיבית בין אטום מימן ממולקולה או שבר מולקולרי X — H שבו X הוא אלקטרונגטיבי יותר מ-H, לבין אטום או קבוצת אטומים באותה מולקולה או אחרת, שבה יש עדויות ליצירת קשר."
חיובים חלקיים
ההבדל באלקטרושליליות בין מימן למולקולה שאליה הוא קשור קוולנטית מוליד מטענים חלקיים כמתואר לעיל. מטענים זעירים אלה (δ+ ו- δ-) גורמים ליצירת קשרי מימן, המתרחשים כאשר המטען החיובי החלקי של אטום מימן נמשך לשלילה החלקית של מולקולה אחרת. במים, זה אומר שהמימן של מולקולת מים אחת נמשך לחמצן של אחרת (איור 1.31). מכיוון שמים הם מולקולה א-סימטרית, המשמעות היא גם שהמטענים אינם סימטריים. חלוקה לא אחידה כזו היא שעושה דיפול. מולקולות דו-קוטביות חשובות לאינטראקציות עם מולקולות דו-קוטביות אחרות ולהמסת חומרים יוניים (איור 1.32).
קשרי מימן אינם בלעדיים למים. למעשה, הם כוחות חשובים המחזיקים יחד מקרומולקולות הכוללות חלבונים וחומצות גרעין. קשרי מימן מתרחשים בתוך ובין מקרומולקולות.
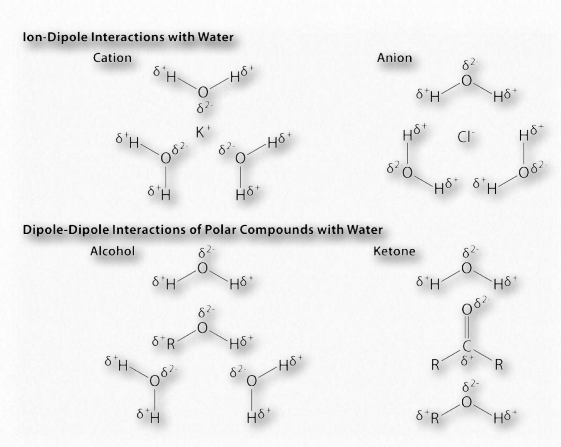
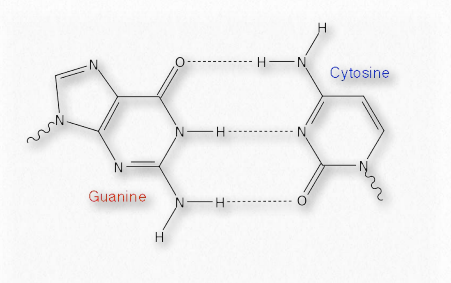
הזיווג המשלים המתרחש בין בסיסים בגדילים מנוגדים של DNA, למשל, מבוסס על קשרי מימן. כל קשר מימן חלש יחסית (בהשוואה לקשר קוולנטי, למשל - טבלה 1.4), אך ביחד הם יכולים להיות חזקים למדי.

טבלה 1.4 תמונה מאת אליה קים
היתרונות של אינטראקציות חלשות
חולשתם, לעומת זאת, למעשה מועילה למדי לתאים, במיוחד בכל הנוגע לחומצות גרעין (איור 1.33). גדילי ה- DNA, למשל, חייבים להיות מופרדים על פני מתיחות קצרות בתהליכי השכפול והסינתזה של RNA. מכיוון שצריך להפריד רק כמה זוגות בסיסים בכל פעם, האנרגיה הנדרשת לשם כך קטנה והאנזימים המעורבים בתהליכים יכולים לפרק אותם בקלות, לפי הצורך. קשרי מימן ממלאים תפקידים גם בקשירת מצעים לאנזימים, קטליזה ואינטראקציה בין חלבון לחלבון, כמו גם סוגים אחרים של קישור, כגון חלבון-DNA או נוגדן-אנטיגן.
כאמור, קשרי מימן חלשים יותר מקשרים קוולנטיים (טבלה 1.4) וחוזקם משתנה בצורה חלשה מאוד (1-2 קילו-ג'יי/מול) לחזקים למדי (29 קילו-ג'יי/מול). קשרי מימן מתרחשים רק למרחקים קצרים יחסית (2.2 עד 4.0 Å). ככל שמרחק קשר המימן רחוק יותר, כך הקשר חלש יותר.
חוזק הקשר ב- kJ/mol מייצג את כמות החום שיש להכניס למערכת כדי לשבור את הקשר - ככל שהמספר גדול יותר, כך חוזק הקשר גדול יותר. קשרי מימן נשברים בקלות באמצעות חום. הרתחת מים, למשל, מחייבת שבירת קשרי H. כאשר מבנה ביולוגי, כגון חלבון או מולקולת DNA, מתייצב על ידי קשרי מימן, שבירת קשרים אלה מערערת את המבנה ועלולה לגרום לדנטורציה של החומר - אובדן מבנה. מסיבה זו רוב החלבונים וכל ה- DNA מאבדים את המבנים המקומיים שלהם, או המקופלים, כאשר הם מחוממים לרתיחה.
תמונה על ידי אליה קים טבלה 1.5
עבור מולקולות DNA, דנטורציה גורמת להפרדה מוחלטת של הגדילים זה מזה. עבור מרבית החלבונים המשמעות היא אובדן המבנה התלת מימדי האופייני שלהם ואיתו אובדן התפקוד שביצעו. למרות שכמה חלבונים יכולים בקלות לשקם את המבנה המקורי שלהם כאשר הפתרון שבו הם נמצאים מקורר, רובם לא יכולים. זו אחת הסיבות שאנחנו מבשלים את האוכל שלנו. חלבונים חיוניים לחיים, ולכן דנטורציה של חלבונים חיידקיים גורמת למוות של כל מיקרואורגניזמים המזהמים את המזון.
החשיבות של מאגרים
מים יכולים ליינן במידה קלה (10-7 M) ליצירת H+ (פרוטון) ו- OH- (הידרוקסיד). אנו מודדים את ריכוז הפרוטונים של תמיסה עם pH, שהוא היומן השלילי של ריכוז הפרוטונים.
pH = -יומן [H+]
אם ריכוז הפרוטונים, [H+] = 10-7 M, אז ה- pH הוא 7. באותה קלות נוכל למדוד את ריכוז ההידרוקסיד עם ה- pOH על ידי המשוואה המקבילה,
PoH = -יומן [הו -]
במים טהורים, דיסוציאציה של פרוטון יוצרת בו זמנית הידרוקסיד, כך שה- PoH של מים טהורים הוא 7, גם כן. זה גם אומר ש
PH + פוח = 14
כעת, מכיוון שפרוטונים והידרוקסידים יכולים להתחבר ליצירת מים, כמות גדולה של אחד תגרום לכמות קטנה של השנייה. מדוע זה המקרה? במילים פשוטות, אם אני זורק 0.1 שומות של H+ לתמיסת מים טהורים, ריכוז הפרוטונים הגבוה יגיב עם הכמות הקטנה יחסית של הידרוקסידים ליצירת מים, ובכך יפחית את ריכוז ההידרוקסיד. באופן דומה, אם אני זורק עודף הידרוקסיד (כמו NaOH, למשל) למים טהורים, ריכוז הפרוטונים נופל מאותה סיבה.
חומצות לעומת בסיסים
כימאים משתמשים במונח "חומצה" כדי להתייחס לחומר שיש בו פרוטונים שיכולים להתנתק (לרדת) כשהוא מומס במים. הם משתמשים במונח "בסיס" כדי להתייחס לחומר שיכול לספוג פרוטונים כשהוא מומס במים. גם חומצות וגם בסיסים מגיעים בצורות חזקות וחלשות. (דוגמאות לחומצות חלשות מוצגות בטבלה 1.5.) חומצות חזקות, כגון HCl, מתנתקות לחלוטין במים. אם נוסיף 0.1 שומות (6.02x1022 מולקולות) של HCl לתמיסה לייצור ליטר, יהיו לו 0.1 שומות של H+ ו- 0.1 שומות של מולקולות Cl- או 6.02x1022 מכל אחת מהן. לא יישאר HCl כשזה יקרה. בסיס חזק כמו NaOH גם מתנתק לחלוטין ל- Na+ ו- OH-.

חומצות חלשות
חומצות ובסיסים חלשים שונים מעמיתיהם החזקים. כאשר מכניסים שומה אחת של חומצה אצטית (hAc) למים טהורים, רק אחוז זעיר ממולקולות ה- hAc מתנתקות ל- H+ ו- Ac-. ברור שחומצות חלשות שונות מאוד מחומצות חזקות. בסיסים חלשים מתנהגים באופן דומה, אלא שהם מקבלים פרוטונים, במקום לתרום אותם. מכיוון שאנו יכולים לראות הכל כצורה של חומצה חלשה, לא נשתמש כאן במונח בסיס חלש.

התלמידים תמהים לעתים קרובות ומצפים ש- [H +] = [A -] מכיוון שמשוואת הדיסוציאציה מציגה אחד מכל אחד מ- HA. זה, למעשה, נכון רק כאשר מותר ל- HA להתנתק במים טהורים. בדרך כלל ה- HA ממוקם בתמיסה שיש בה פרוטונים והידרוקסידים כדי להשפיע על דברים. אותם פרוטונים ו/או הידרוקסידים משנים את ה- H+ ואת הריכוז באופן לא שווה, מכיוון ש- A- יכול לספוג חלק מהפרוטונים ו/או HA יכולים לשחרר את H+ כאשר הם מושפעים מה- OH- בתמיסה. לכן, יש לחשב את ריכוז הפרוטונים מה-pH באמצעות משוואת הנדרסון האסלבלך.
\[pH = pKa + log ([Ac- ]/[HAc])\]

תמונה על ידי אליה קים טבלה 1.6
אתם עשויים לתהות מדוע אכפת לנו מחומצות חלשות. יתכן שמעולם לא חשבת הרבה על חומצות חלשות כשהיית בכימיה כללית. המדריך שלך תיאר אותם כמאגרים וכנראה שיננת בצייתנות את העובדה ש"מאגרים הם חומרים המתנגדים לשינוי ב- pH "מבלי ללמוד באמת מה פירוש ניקוי בלבול - פירוש הדבר. מאגרים חשובים מכדי לחשוב עליהם בדרך זו.
עליות
חומצות חלשות הן קריטיות לחיים מכיוון שהזיקה שלהן לפרוטונים גורמת להן להתנהג כמו UPS. איננו מתייחסים ל- UPS שהוא שירות החבילות המאוחד, אלא במקום זאת, למערכות גיבוי הסוללה העטופות למחשבים הנקראים ספקי כוח ללא הפרעה הבועטים כדי לשמור על הפעלת מחשב במהלך הפסקת חשמל. הסוללה במחשב נייד היא UPS, למשל.
אנו יכולים לחשוב על חומצות חלשות כספקי פרוטון ללא הפרעה בטווחי pH מסוימים, המספקים (או סופגים) פרוטונים לפי הצורך. חומצות חלשות עוזרות אפוא לשמור על ריכוז H+ (ובכך ה- pH) של התמיסה בה הן קבועות יחסית.
שקול את מערכת הביקרבונט/חומצה פחמנית. איור 1.35 מראה מה קורה כאשר H 2 CO 3 מתנתק. הוספת יוני הידרוקסיד (על ידי הוספת בסיס חזק כמו NaOH) לתמיסה גורמת ליוני H+ להגיב עם יוני OH ליצירת מים. כתוצאה מכך, ריכוז יוני H+ יירד וה- pH יעלה.
עם זאת, בניגוד למצב עם פתרון של מים טהורים, יש מקור גיבוי של H+ זמין בצורה של H 2 CO 3. כאן נכנסת פונקציית UPS. כאשר פרוטונים נלקחים על ידי יוני ההידרוקסיל הנוספים (מייצרים מים), הם מוחלפים בחלקם בפרוטונים מה- H 2 CO 3. זו הסיבה שחומצה חלשה היא חיץ. הוא מתנגד לשינויים ב- pH על ידי שחרור פרוטונים כדי לפצות על אלה ש"נוצלו "בתגובה עם יוני ההידרוקסיל.

הנדרסון-האסלבלך
כדאי להיות מסוגל לחזות את התגובה של מערכת H 2 CO 3 לשינויים בריכוז H+. משוואת הנדרסון-האסלבלך מגדירה את הקשר בין pH ליחס של HCO 3 - ו- H 2 CO 3. זה הוא
pH = pKa + יומן ([HCO 3 -]/[H 2 CO 3])
משוואה פשוטה זו מגדירה את הקשר בין ה- pH של תמיסה לבין היחס בין HCO 3 - ו- H 2 CO 3 בה. המונח החדש, הנקרא pKa, מוגדר כ
pKa = -יומן K a,
בדיוק כמו
pH = -יומן [H +].
ה- Ka הוא קבוע ניתוק החומצה ומהווה מדד לחוזקה של חומצה. עבור חומצה כללית, HA, שמתנתקת כ
חה ח + א -, ק א = [H +] [א -]/[חה]
לפיכך, ככל שהחומצה חזקה יותר, כך הפרוטונים יתנתקו ממנה יותר כאשר יתווספו למים וככל שיהיה לערך ה- Ka שלה גדול יותר. ערכים גדולים של Ka מתורגמים לערכים נמוכים יותר של pKa. כתוצאה מכך, ככל שערך ה- pKa נמוך יותר עבור חומצה נתונה, כך החומצה החלשה חזקה יותר.
קבוע pKa
שים לב כי pKa הוא קבוע עבור חומצה נתונה. ה- pKa לחומצה פחמנית הוא 6.37. לשם השוואה, ה- pKa לחומצה פורמית הוא 3.75. חומצה פורמית היא אפוא חומצה חזקה יותר מחומצה אצטית. לחומצה חזקה יותר יהיו יותר פרוטונים המנותקים ב- pH נתון מאשר חומצה חלשה יותר.
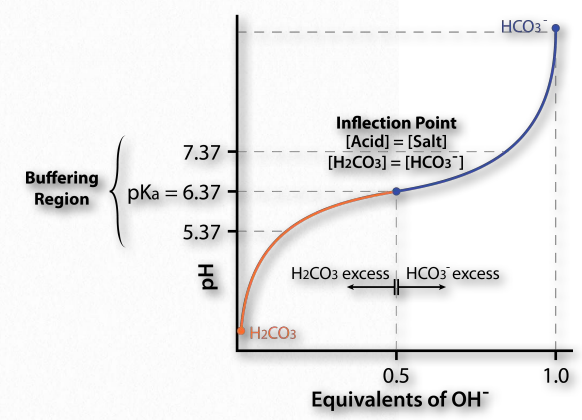
עכשיו, איך זה מתורגם לייצוב pH? איור 1.35 מציג עקומת טיטרציה. בעקומה זו, הטיטרציה מתחילה בתנאים בפינה השמאלית התחתונה (pH נמוך מאוד). ב- pH זה, צורת H 2 CO 3 שולטת, אך ככל שמתווספים יותר ויותר OH- (עוברים ל- 45 מדוע אכפת לנו מ- pH? מכיוון שמולקולות ביולוגיות יכולות, במקרים מסוימים, להיות רגישות להפליא לשינויים בה. ככל שה-pH של תמיסה משתנה, המטענים של מולקולות בתמיסה יכולים להשתנות, כפי שתראו. שינוי מטענים על מולקולות ביולוגיות, במיוחד חלבונים, יכול להשפיע באופן דרסטי על אופן פעולתן ואפילו אם הן פועלות בסדר), ה- pH עולה, כמות ה- HCO3- עולה ו (בהתאמה), כמות H 2 CO 3 יורדת. שימו לב שהעקומה "משתטחת" ליד ה- pKa (6.37).
אזור חציצה
שיטוח העקומה אומר לנו שה- pH לא משתנה הרבה (לא עולה מהר) כפי שעשה קודם לכן כאשר נוספה אותה כמות הידרוקסיד. המערכת מתנגדת לשינוי ב-pH (לא עוצרת את השינוי, אלא מאטה אותו) באזור של יחידת pH אחת מעל ויחידת pH אחת מתחת ל-pKa. לפיכך, אזור החציצה של מאגר החומצה הפחמנית/ביקרבונט הוא בערך 5.37 עד 7.37. הוא חזק ביותר ב- pH של 6.37.
עכשיו זה מתחיל להתברר איך המאגר עובד. HA יכול לתרום פרוטונים כאשר יש צורך בתוספות (כגון כאשר OH- מתווסף לפתרון על ידי תוספת של NaOH). באופן דומה, A- יכול לקבל פרוטונים כאשר H+ נוספים מתווספים לתמיסה (הוספת HCl, למשל). היכולת המרבית לתרום או לקבל פרוטונים מגיעה מתי
[A -] = [חה]
זה עולה בקנה אחד עם משוואת הנדרסון האסלבלך ועקומת הטיטרציה. כאשר [A -] = [HA], pH = 6.37+ יומן (1). מאז יומן (1) = 0, pH = 6.37 = pKa עבור חומצה פחמנית. לפיכך עבור כל מאגר, למאגר יהיה חוזק מרבי ושיטוח תצוגה של עקומת הטיטרציה שלו כאשר [A -] = [HA] ומתי pH = pKa. אם למאגר יש יותר מ- pKa אחד (איור 1.36), כל אזור pKa יציג את ההתנהגות.
נאגר לעומת לא נאגר
כדי להבין עד כמה מאגר מגן מפני שינויים ב-pH, שקול את ההשפעה של הוספת .01 שומות של HCl ל-1.0 ליטר מים טהורים (ללא שינוי נפח) ב-pH 7, בהשוואה להוספתו ל-1.0 ליטר של חיץ אצטט 1M ב-pH 4.76. מכיוון ש- HCl מתנתק לחלוטין, ב- 0.01M (10-2 M) HCl יהיה לך 0.01M H +. עבור המים הטהורים, ה-pH יורד מ-7.0 למטה ל-2.0 (pH = -log (0.01M)).
לעומת זאת, ה- pH של מאגר האצטט לאחר הוספת אותה כמות של HCl הוא 4.74. לפיכך, תמיסת המים הטהורים רואה את ה-pH שלה יורד מ-7 ל-2 (5 יחידות pH), בעוד שהתמיסה שנאגרה ראתה את ה-pH שלה יורד מ-4.76 ל-4.74 (0.02 יחידות pH). ברור שהמאגר ממזער את ההשפעה של הפרוטונים שנוספו בהשוואה למים הטהורים.
קיבולת מאגר
חשוב לציין שלמאגרים יש יכולות מוגבלות על ידי ריכוזם. בואו נדמיין שבפסקה הקודמת הוספנו את 0.01 שומות HCl למאגר אצטט בעל ריכוז של 0.01M וכמויות שוות של Ac- ו- hAc. כאשר אנו מנסים לעשות את המתמטיקה במקביל לחישוב הקודם, אנו רואים שיש 0.01M פרוטונים, אך רק 0.005M A- לקלוט אותם. יכולנו לדמיין ש 0.005M מהפרוטונים ייספגו, אבל זה עדיין ישאיר 0.005M של פרוטונים ללא אגירה. לפיכך, ה- pH של תמיסה זו יהיה בערך
pH = -יומן (0.005 מ') = 2.30
חריגה מיכולת החיץ הורידה את ה- pH באופן משמעותי בהשוואה להוספת אותה כמות של פרוטונים למאגר אצטט 1M. כתוצאה מכך, כאשר בוחנים מאגרים, חשוב להכיר בכך שהריכוז שלהם מציב את גבולותיהם. גבול נוסף הוא טווח ה- pH בו מקווים לשלוט בריכוז הפרוטונים.
קבוצות מיוננות מרובות
עכשיו, מה קורה אם למולקולה יש שתי קבוצות מיוננות (או יותר)? מסתבר, באופן לא מפתיע, שלכל קבוצה יהיה pKa משלה וכתוצאה מכך יהיו מספר אזורי חציצה.
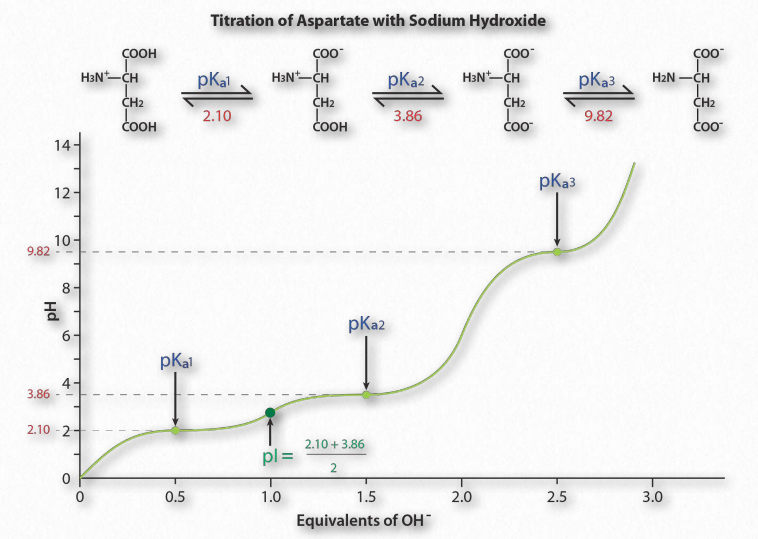
איור 1.36 מציג את עקומת הטיטרציה של חומצת האמינו אספרטית. שים לב שבמקום שיטוח יחיד של העקומה, כפי שנראה עבור חומצה אצטית, עקומת הטיטרציה של חומצה אספרטית מציגה שלושה אזורים כאלה. אלו הם אזורי חציצה בודדים, שכל אחד מהם מרוכז בערכי pKa המתאימים לקבוצת הקרבוקסיל ולקבוצת האמין.
לחומצה אספרטית יש ארבעה מטענים אפשריים: +1 (קבוצת α-carboxyl, קבוצת α-amino ו- Rלקבוצת קרבוקסיל לכל אחד יש פרוטון), 0 (קבוצת α- קרבוקסיל חסרה פרוטון, לקבוצת α- יש פרוטון, לקבוצת R יש פרוטון), -1 (קבוצת α-carboxyl וקרבוקסיל מקבוצת R חסרים כל אחד פרוטון, קבוצת α-amino שומרת על פרוטון), -2 (α קבוצות קרבוקסיל, קרבוקסיל מקבוצת R וקבוצות α-אמינו חסרות פרוטון נוסף).
חיזוי
כיצד ניתן לחזות את המטען של חומצת אמינו ב-pH נתון? כלל אצבע טוב להערכת מטען הוא שאם ה-pH נמצא יותר מיחידה אחת מתחת ל-pKa עבור קבוצה (קרבוקסיל או אמינו), הפרוטון פועל. אם ה-pH הוא יותר מיחידה אחת מעל ה-pKa עבור הקבוצה, הפרוטון כבוי. אם ה- pH אינו יותר מיחידת pH אחת או פחות מ- pKa, הנחה פשוטה זו לא תעבוד.
יתר על כן, חשוב להכיר בכך שכללי אצבע אלה הם הערכות בלבד. ה-pI (pH שבו המטען של מולקולה הוא אפס) הוא ערך מדויק המחושב כממוצע של שני ערכי pKa משני צידי אזור האפס. זה מחושב בממוצע של שני ערכי pKa סביב הנקודה שבה המטען של המולקולה הוא אפס. עבור חומצה אספרטית, זה מתאים pK a1 ו pK a2.
הפניות
- http://www.lpi.usra.edu/lunar/missions/apollo/ אפולו_12/ניסויים/מודד/
- ארונן, אלנגנן; Desiraju, Gautam R.; קליין, רוג'ר א '; סדליי, ג'ואנה; שיינר, סטיב; אלקורטה, איבון; קלרי, דייוויד סי; קראבטרי, רוברט ה'; דננברג, ג'וזף ג'יי; הובזה, פאבל; קיירגארד, הנריק ג '; לגון, אנתוני סי; מננוצ'י, בנדטה; נסביט; נסביט, דייוויד ג'יי (2011). "הגדרת קשר המימן". Appl טהור. כימיה 83 (8): 1637—1641. דוי:10.1351/פאק-רק-10-01-02