
11.2: Replication DNA
- Page ID
- 174737

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Malengo ya kujifunza
- Eleza maana ya replication ya DNA ya semiconservative
- Eleza kwa nini replication DNA ni bidirectional na inajumuisha wote kuongoza na lagging strand
- Eleza kwa nini vipande vya Okazaki vinaundwa
- Eleza mchakato wa replication ya DNA na kazi za enzymes zinazohusika
- Kutambua tofauti kati ya replication DNA katika bakteria na eukaryotes
- Eleza mchakato wa kuiga mzunguko wa mzunguko
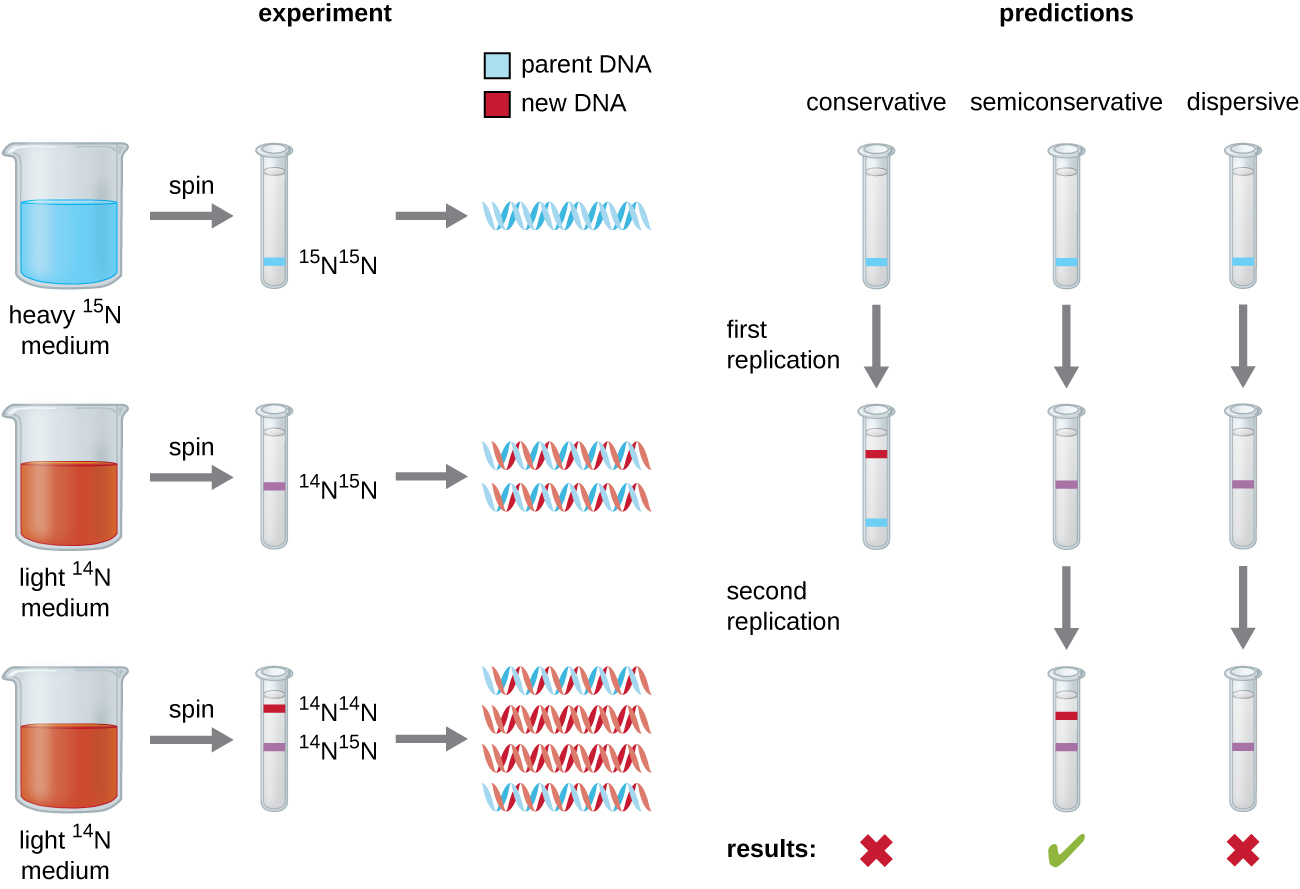
Ufafanuzi wa muundo wa helix mara mbili na James Watson na Francis Crick mwaka 1953 ulitoa hint kuhusu jinsi DNA inakiliwa wakati wa mchakato wa kuiga. Kutenganisha vipande vya helix mara mbili ingeweza kutoa templates mbili kwa ajili ya awali ya vipande vipya vya ziada, lakini hasa jinsi molekuli mpya za DNA zilijengwa bado haijulikani. Katika mfano mmoja, replication ya semiconservative, vipande viwili vya helix mbili tofauti wakati wa replication ya DNA, na kila strand hutumika kama template ambayo strand mpya ya ziada inakiliwa; baada ya kuiga, kila DNA iliyopigwa mara mbili inajumuisha kamba moja ya wazazi au “zamani” na kamba moja “mpya”. Kulikuwa na mifano miwili ya ushindani pia ilipendekeza: kihafidhina na kutawanyika, ambazo zinaonyeshwa kwenye Kielelezo\(\PageIndex{1}\).

Mathayo Meselson (1930—) na Franklin Stahl (1929—) walipanga majaribio katika 1958 mtihani ni ipi ya mifano hii kwa usahihi inawakilisha DNA replication (Kielelezo\(\PageIndex{2}\)). Walikua E. koli kwa vizazi kadhaa katika kati iliyo na isotopu “nzito” ya nitrojeni (15 N) iliyoingizwa katika besi za nitrojeni na, hatimaye, ndani ya DNA. Hii kinachoitwa DNA ya wazazi. Utamaduni wa E. koli kisha ulibadilishwa kuwa kati iliyo na 14 N na kuruhusiwa kukua kwa kizazi kimoja. Seli zilivunwa na DNA ikatengwa. DNA ilitenganishwa na ultracentrifugation, wakati ambapo DNA iliunda bendi kulingana na wiani wake. DNA iliyopandwa katika 15 N itatarajiwa kuunda bendi katika nafasi ya juu ya wiani kuliko ile iliyopandwa katika 14 N. Meselson na Stahl alibainisha kuwa baada ya kizazi kimoja cha ukuaji katika 14 N, bendi moja iliyoonekana ilikuwa kati katika nafasi kati ya DNA ya seli zilizopandwa peke katika 15 N au 14 N. hii ilipendekeza ama semiconservative au kutawanyika mode ya replication. Baadhi ya seli ziliruhusiwa kukua kwa kizazi kimoja zaidi katika 14 N na kuzunguka tena. DNA iliyovunwa kutoka seli zilizopandwa kwa vizazi viwili katika 14 N iliunda bendi mbili: bendi moja ya DNA ilikuwa katika nafasi ya kati kati ya 15 N na 14 N, na nyingine ilifanana na bendi ya 14 N DNA. Matokeo haya yanaweza kuelezewa tu ikiwa DNA inaiga kwa namna ya semiconservative. Kwa hiyo, mifano mingine miwili ilitolewa nje. Kama matokeo ya jaribio hili, sasa tunajua kwamba wakati wa replication ya DNA, kila moja ya vipande viwili vinavyotengeneza helix mara mbili hutumika kama template ambayo vipande vipya vinakiliwa. Kipande kipya kitakuwa cha ziada kwa mkondo wa wazazi au “wa zamani”. Molekuli za DNA zinazosababisha zina mlolongo sawa na zinagawanywa sawa katika seli mbili za binti.
Zoezi\(\PageIndex{1}\)
Nini ingekuwa hitimisho la majaribio ya Meselson na Stahl ikiwa, baada ya kizazi cha kwanza, walipata bendi mbili za DNA?
Replication DNA katika Bakteria
Replication ya DNA imejifunza vizuri katika bakteria hasa kwa sababu ya ukubwa mdogo wa jenomu na mutants zinazopatikana. E. koli ina jozi za msingi milioni 4.6 (Mbp) katika kromosomu moja ya mviringo na zote zinaigwa katika takriban dakika 42, kuanzia asili moja ya kuiga na kuendelea kuzunguka mduara bidirectionally (yaani, katika pande zote mbili). Hii ina maana kwamba takriban nyukleotidi 1000 huongezwa kwa pili. Mchakato huu ni wa haraka sana na hutokea kwa makosa machache.
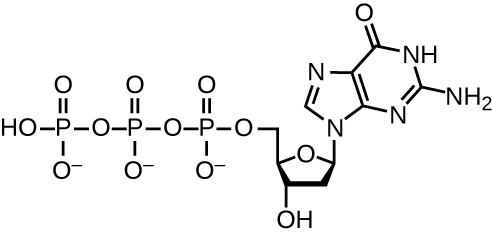
Replication ya DNA hutumia idadi kubwa ya protini na enzymes (Jedwali\(\PageIndex{1}\)). Mmoja wa wachezaji muhimu ni enzyme DNA polymerase, pia inajulikana kama DNA pol. Katika bakteria, aina tatu kuu za polima za DNA zinajulikana: DNA pol I, DNA pol II, na DNA pol III. Sasa inajulikana kuwa DNA pol III ni enzyme zinazohitajika kwa ajili ya awali ya DNA; DNA pol I na DNA pol II kimsingi zinahitajika kwa ajili ya ukarabati. DNA pol III anaongeza deoxyribonucleotides kila nyongeza kwa nyukleotidi kwenye kamba ya template, moja kwa moja kwa kundi la 3'-OH la mlolongo wa DNA unaoongezeka. Kuongezea kwa nucleotides hizi inahitaji nishati. Nishati hii iko katika vifungo vya makundi matatu ya phosphate yaliyounganishwa na kila nucleotide (nucleotide ya triphosphate), sawa na jinsi nishati iliyohifadhiwa katika vifungo vya phosphate ya adenosine triphosphate (ATP) (Kielelezo\(\PageIndex{3}\)). Wakati dhamana kati ya phosphates ni kuvunjwa na kutolewa diphosphate, nishati iliyotolewa inaruhusu malezi ya covalent phosphodiester dhamana na upungufu wa maji mwilini awali kati ya nyukleotidi zinazoingia na bure 3'-OH kundi juu ya kukua DNA strand.
Uanzishwaji
Kuanzishwa kwa replication hutokea katika mlolongo maalum wa nucleotide inayoitwa asili ya replication, ambapo protini mbalimbali hufunga ili kuanza mchakato wa kuiga. E. koli ina asili moja ya kuiga (kama vile prokaryotes nyingi), inayoitwa ORIC, kwenye kromosomu yake moja. Asili ya kuiga ni takriban jozi 245 za msingi kwa muda mrefu na ni matajiri katika utaratibu wa adenine-thymine (AT).
Baadhi ya protini zinazofunga kwa asili ya kuiga ni muhimu katika kufanya mikoa moja-stranded ya DNA kupatikana kwa replication. DNA ya chromosomal ni kawaida amefungwa karibu histones (katika eukaryotes na archaea) au histone-kama protini (katika bakteria), na ni supercoiled, au sana amefungwa na inaendelea yenyewe. Ufungaji huu hufanya habari katika molekuli ya DNA haipatikani. Hata hivyo, enzymes zinazoitwa topoisomerases zinabadilisha sura na supercoiling ya kromosomu. Kwa replication ya DNA ya bakteria kuanza, kromosomu ya supercoiled imetulia na topoisomerase II, pia huitwa DNA gyrase. Enzyme inayoitwa helixase kisha hutenganisha vipande vya DNA kwa kuvunja vifungo vya hidrojeni kati ya jozi za msingi za nitrojeni. Kumbuka kwamba Utaratibu wa AT una vifungo vichache vya hidrojeni na, kwa hiyo, huwa na mwingiliano dhaifu kuliko utaratibu wa guanine-cytosine (GC). Enzymes hizi zinahitaji hidrolisisi ya ATP. Kama DNA inafungua, miundo yenye umbo la Y inayoitwa uma za replication hutengenezwa. Mbili uma replication ni sumu katika asili ya replication, kuruhusu kwa bidirectional replication na malezi ya muundo ambayo inaonekana kama Bubble wakati kutazamwa na microscope ya elektroni maambukizi; Matokeo yake, muundo huu huitwa Bubble replication. DNA karibu na kila uma replication ni coated na protini moja stranded kisheria ili kuzuia DNA single-stranded kurejea katika helix mara mbili.
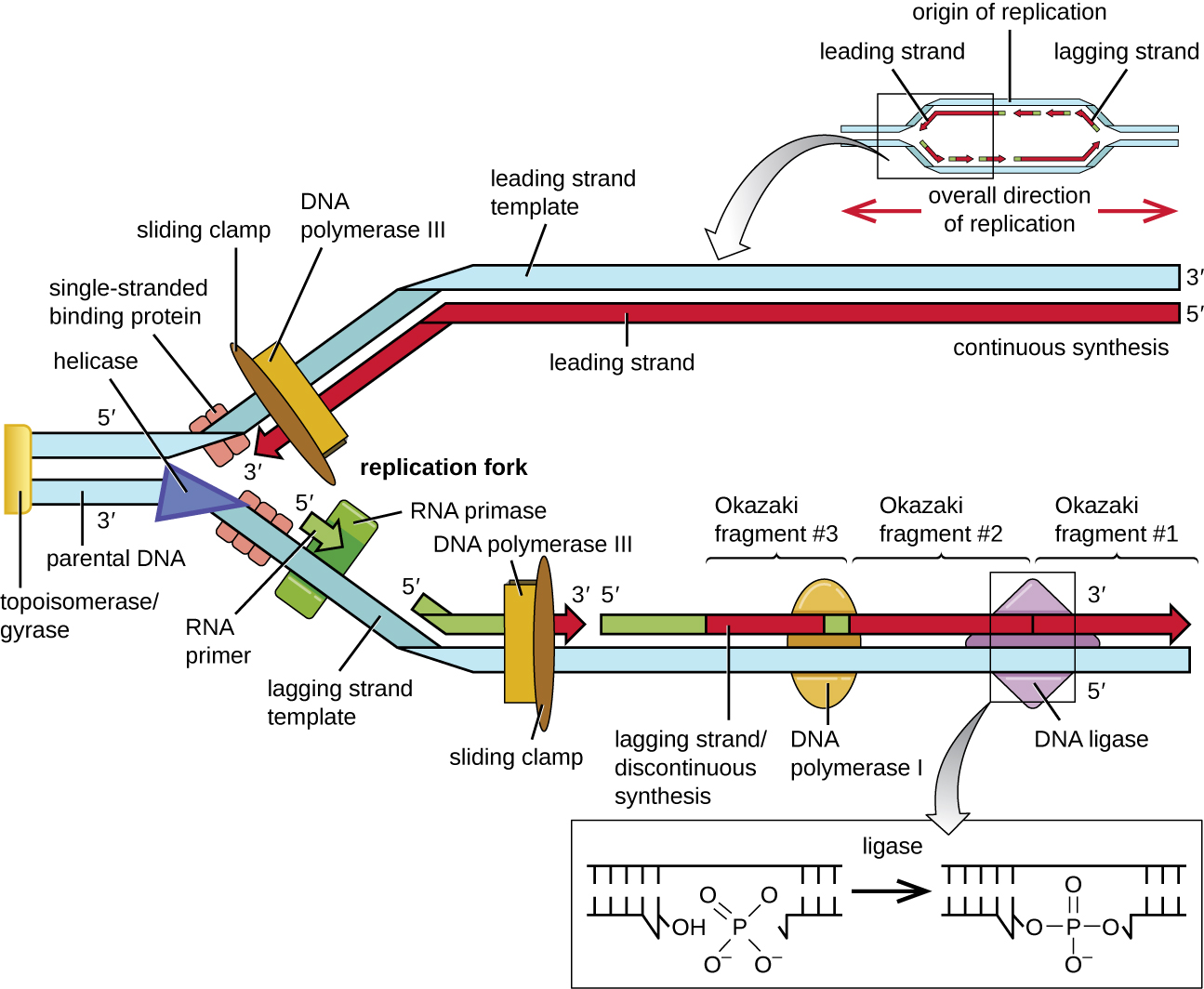
Mara baada ya DNA moja-stranded inapatikana katika asili ya replication, DNA replication inaweza kuanza. Hata hivyo, DNA pol III ina uwezo wa kuongeza nucleotidi tu katika mwelekeo wa 5' hadi 3' (kamba mpya ya DNA inaweza kupanuliwa tu katika mwelekeo huu). Hii ni kwa sababu DNA polymerase inahitaji bure 3'-OH kundi ambayo inaweza kuongeza nyukleotidi kwa kutengeneza covalent phosphodiester dhamana kati ya mwisho 3'-OH na 5' phosphate ya nucleotide ijayo. Hii pia inamaanisha kuwa haiwezi kuongeza nyukleotidi ikiwa kikundi cha bure cha 3'-OH hakipatikani, ambacho ndicho kesi kwa strand moja ya DNA. Tatizo linatatuliwa kwa msaada wa mlolongo wa RNA ambao hutoa mwisho wa bure wa 3'-OH. Kwa sababu mlolongo huu inaruhusu kuanza kwa awali ya DNA, inaitwa ipasavyo primer. The primer ni nucleotides tano hadi 10 kwa muda mrefu na inayoongezea DNA ya wazazi au template. Inatengenezwa na RNA primase, ambayo ni polymerase ya RNA. Tofauti na polymerases ya DNA, polymerases ya RNA hazihitaji kikundi cha bure cha 3'-OH ili kuunganisha molekuli ya RNA. Sasa kwa kuwa primer hutoa kikundi cha bure cha 3'-OH, DNA polymerase III sasa inaweza kupanua primer hii ya RNA, na kuongeza nucleotides ya DNA moja kwa moja ambayo ni ya ziada kwa kamba ya template (Kielelezo\(\PageIndex{1}\)).
Elongation
Wakati wa upungufu katika replication ya DNA, kuongeza ya nucleotides hutokea kwa kiwango chake cha juu cha nucleotides 1000 kwa pili. DNA polymerase III inaweza tu kupanua katika mwelekeo 5' hadi 3', ambayo inaleta tatizo katika uma replication. Heli mbili ya DNA ni antiparallel; yaani strand moja inaelekezwa katika mwelekeo wa 5' hadi 3' na nyingine inaelekezwa katika mwelekeo wa 3' hadi 5' (tazama Muundo na Kazi ya DNA). Wakati wa replication, strand moja, ambayo ni nyongeza kwa 3' kwa 5' wazazi DNA strand, ni synthesized kuendelea kuelekea replication uma kwa sababu polymerase inaweza kuongeza nucleotides katika mwelekeo huu. Kamba hii inayoendelea synthesized inajulikana kama strand inayoongoza. Kamba nyingine, inayoongezea DNA ya wazazi wa 5' hadi 3', inakua mbali na uma ya kuiga, hivyo polymerase inapaswa kurudi nyuma kuelekea njia ya kuiga ili kuanza kuongeza besi kwa primer mpya, tena katika mwelekeo mbali na uma ya kuiga. Inafanya hivyo mpaka inakabiliwa na kamba iliyotengenezwa hapo awali na kisha inarudi tena (Kielelezo\(\PageIndex{4}\)). Hatua hizi huzalisha vipande vidogo vya mlolongo wa DNA vinavyojulikana kama vipande vya Okazaki, kila ikitenganishwa na utangulizi wa RNA. Vipande vya Okazaki vinatajwa baada ya timu ya utafiti wa Kijapani na wanandoa wa ndoa Reiji na Tsuneko Okazaki, ambao waligundua kwanza mwaka 1966. The strand na vipande Okazaki inajulikana kama strand lagging, na awali yake inasemekana kuwa discontinuous.
Kamba inayoongoza inaweza kupanuliwa kutoka kwa primer moja peke yake, wakati strand lagging inahitaji primer mpya kwa kila moja ya vipande mfupi Okazaki. Mwelekeo wa jumla wa strand ya lagging itakuwa 3' hadi 5', na ile ya strand inayoongoza 5' hadi 3'. Protini inayoitwa clamp sliding inashikilia polimerasi ya DNA mahali inapoendelea kuongeza nyukleotidi. Kamba ya kupiga sliding ni protini yenye umbo la pete inayofunga kwa DNA na inashikilia polymerase mahali. Zaidi ya jukumu lake katika uanzishwaji, topoisomerase pia inazuia overwinding ya DNA mara mbili helix mbele ya uma replication kama DNA inafungua; inafanya hivyo kwa kusababisha nicks muda katika helix DNA na kisha kuziba tena. Kama awali inavyoendelea, primers ya RNA hubadilishwa na DNA. Vipindi vinaondolewa na shughuli za exonuclease za DNA polymerase I, na mapungufu yanajazwa. Nicks iliyobaki kati ya DNA mpya synthesized (ambayo badala ya primer RNA) na awali synthesized DNA ni muhuri na enzyme DNA ligase ambayo huchochea malezi ya covalent phosphodiester uhusiano kati ya 3'-OH mwisho wa kipande moja DNA na na 5' phosphate mwisho wa kipande kingine, kuimarisha uti wa mgongo wa sukari-phosphate wa molekuli ya DNA.
Kusitishwa
Mara baada ya kromosomu kamili imepigwa, kusitishwa kwa replication ya DNA lazima kutokea. Ingawa mengi inajulikana kuhusu uanzishwaji wa replication, chini inajulikana kuhusu mchakato wa kusitisha. Kufuatia replication, kusababisha kamili genomes mviringo ya prokaryotes ni concatenated, maana kwamba chromosomes ya DNA mviringo ni interlocked na lazima kutengwa kutoka kwa kila mmoja. Hii inakamilika kupitia shughuli ya topoisomerase IV ya bakteria, ambayo huanzisha mapumziko mara mbili-stranded katika molekuli DNA, kuruhusu yao kujitenga kutoka kwa kila mmoja; enzyme kisha reseals chromosomes mviringo. Azimio la concatemers ni suala la pekee kwa replication ya DNA ya prokaryotiki kwa sababu ya chromosomes zao za mviringo. Kwa sababu wote bakteria DNA gyrase na topoisomerase IV ni tofauti na wenzao eukaryotiki, enzymes hizi hutumika kama malengo ya darasa la dawa za antimicrobial iitwayo quinolones.
| Enzyme au Factor | Kazi |
|---|---|
| DNA POL I | Shughuli ya exonuclease huondoa primer ya RNA na kuibadilisha na DNA iliyopangwa |
| DNA pol III | Kuu enzyme kwamba anaongeza nucleotides katika 5' kwa 3' mwelekeo |
| Helicase | Inafungua helix ya DNA kwa kuvunja vifungo vya hidrojeni kati ya besi za nitrojeni |
| Ligase | Inaweka mapungufu kati ya vipande vya Okazaki kwenye kamba ya kuacha ili kuunda kamba moja ya DNA inayoendelea |
| Primase | Synthesizes RNA primers zinahitajika kuanza replication |
| Protini za kumfunga moja | Funga kwa DNA moja-stranded kuzuia bonding hidrojeni kati ya strands DNA, kurekebisha DNA mbili |
| Sliding clamp | Husaidia kushikilia DNA pol III katika nafasi wakati nyukleotidi ni kuwa aliongeza |
| Topisomerase II (DNA gyrase) | Relaxes kromosomu supercoiled kufanya DNA kupatikana zaidi kwa ajili ya kuanzishwa kwa replication; husaidia kupunguza matatizo ya DNA wakati unwinding, kwa kusababisha mapumziko na kisha resealing DNA |
| Topoisomerase IV | Inaanzisha kuvunja moja-stranded katika chromosomes concatenated kutolewa kutoka kwa kila mmoja, na kisha reseals DNA |
Zoezi\(\PageIndex{2}\)
- Ni enzyme ipi inayovunja vifungo vya hidrojeni vinavyoshikilia vipande viwili vya DNA pamoja ili kuiga kunaweza kutokea?
- Je, ni strand ya kuacha au strand inayoongoza ambayo inaunganishwa katika mwelekeo kuelekea ufunguzi wa uma ya replication?
- Ambayo enzyme ni wajibu wa kuondoa primers RNA katika wapya replicated bakteria DNA?
Replication DNA katika Eukaryotes
Jenomu za Eukaryotic ni ngumu zaidi na kubwa zaidi kuliko genomes za prokaryotic na huwa na chromosomes nyingi za mstari (Jedwali\(\PageIndex{2}\)). Jenomu ya binadamu, kwa mfano, ina jozi bilioni 3 kwa seti ya haploidi ya chromosomes, na jozi za msingi bilioni 6 zinaingizwa wakati wa kuiga. Kuna asili nyingi za kuiga kwenye kila kromosomu ya eukaryotiki (Kielelezo\(\PageIndex{5}\)); jenome ya binadamu ina asili 30,000 hadi 50,000 ya kuiga. Kiwango cha replication ni takriban nyukleotidi 100 kwa pili-mara 10 polepole kuliko replication prokaryotic.
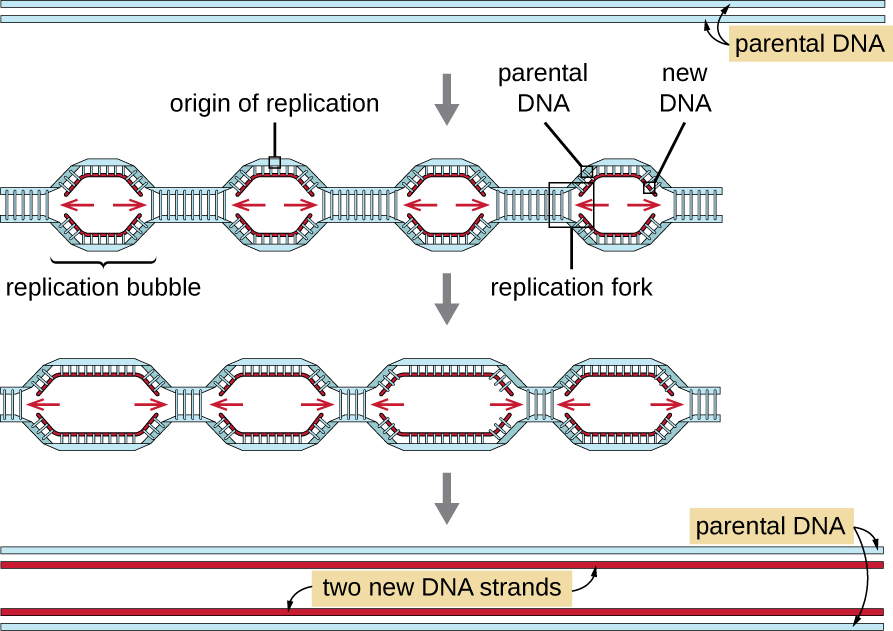
Hatua muhimu za kuiga katika eukaryotes ni sawa na katika prokaryotes. Kabla ya kuiga inaweza kuanza, DNA inapaswa kupatikana kama template. DNA ya Eukaryotic ni yenye supercoiled na vifurushi, ambayo inawezeshwa na protini nyingi, ikiwa ni pamoja na histones (tazama Muundo na Kazi ya Genomes za mkononi). Katika asili ya replication, tata prerelication linajumuisha protini kadhaa, ikiwa ni pamoja na helixase, aina na kuajiri Enzymes nyingine kushiriki katika uanzishwaji wa replication, ikiwa ni pamoja na topoisomerase kupumzika supercoiling, single-stranded kisheria protini, RNA primase, na DNA polymerase. Kufuatia kuanzishwa kwa replication, katika mchakato sawa na ule unaopatikana katika prokaryotes, elongation ni kuwezeshwa na eukaryotic DNA polymerases. Kamba inayoongoza inaendelea kuunganishwa na enzyme ya eukaryotic polymerase pol δ, wakati strand ya kuchelewa inaunganishwa na pol ε. Protini ya kamba ya sliding inashikilia polimerase ya DNA mahali ili iingie DNA. Ribonuclease enzyme H (RNase H), badala ya polymerase ya DNA kama katika bakteria, huondoa primer ya RNA, ambayo inabadilishwa na nucleotides ya DNA. Mapungufu yaliyobaki yanatiwa muhuri na DNA ligase.
Kwa sababu kromosomu za eukaryotic ni linear, mtu anaweza kutarajia kuwa replication yao itakuwa zaidi ya moja kwa moja. Kama ilivyo katika prokaryotes, polymerase ya DNA ya eukaryotic inaweza kuongeza nucleotidi tu katika mwelekeo wa 5' hadi 3'. Katika strand inayoongoza, awali inaendelea mpaka kufikia mwisho wa chromosome au njia nyingine ya replication inayoendelea katika mwelekeo kinyume. Juu ya kamba iliyopungua, DNA inatengenezwa kwa muda mfupi, ambayo kila mmoja huanzishwa na primer tofauti. Wakati uma wa replication unafikia mwisho wa kromosomu linear, hakuna nafasi ya kufanya primer kwa kipande cha DNA kunakiliwa mwishoni mwa kromosomu. Hizi mwisho hivyo kubaki unpaired na, baada ya muda, wanaweza kupata kuendelea mfupi kama seli kuendelea kugawanya.
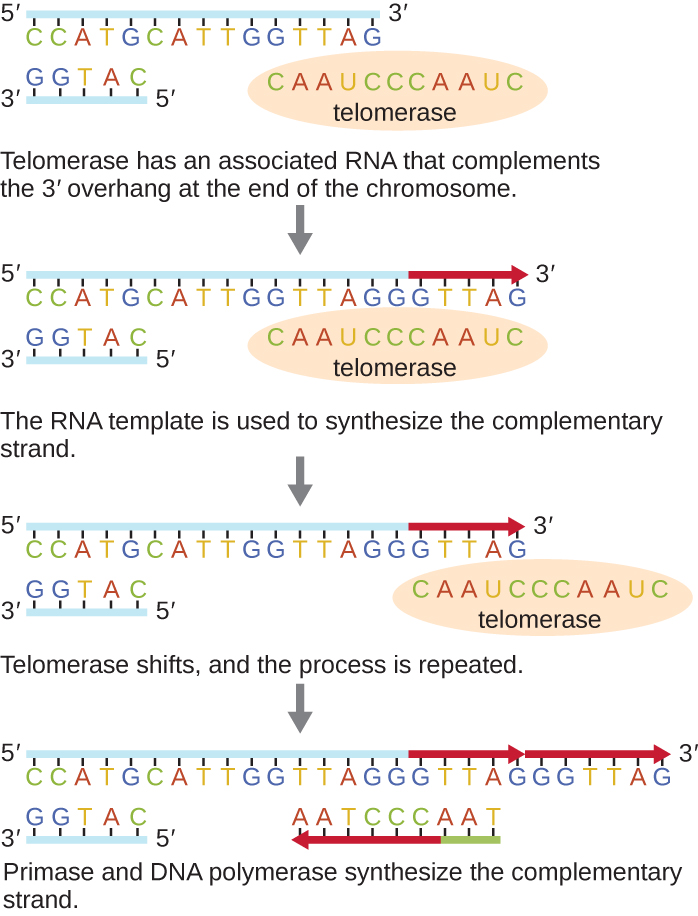
Mwisho wa chromosomes linear hujulikana kama telomeres na hujumuisha utaratibu usio na msimbo wa kurudia. Telomeres kulinda utaratibu wa coding kutoka kupotea kama seli zinaendelea kugawanya. Kwa binadamu, mlolongo sita wa msingi wa jozi, TTAGGG, hurudiwa mara 100 hadi 1000 ili kuunda telomere. Ugunduzi wa telomerase ya enzyme (Kielelezo\(\PageIndex{6}\)) ulifafanua ufahamu wetu wa jinsi mwisho wa chromosome huhifadhiwa. Telomerase ina sehemu ya kichocheo na template iliyojengwa katika RNA. Inaunganisha hadi mwisho wa kromosomu, na misingi ya ziada kwa template ya RNA huongezwa kwenye mwisho wa 3' wa kamba ya DNA. Mara baada ya mwisho wa 3' wa template ya strand ya kuacha ni kutosha vidogo, DNA polymerase inaweza kuongeza nyukleotidi inayoongezea mwisho wa chromosomes. Kwa njia hii, mwisho wa chromosomes huigwa. Kwa binadamu, telomerase ni kawaida kazi katika seli kijidudu na seli shina watu wazima; ni si kazi katika seli watu wazima kuacha za kimwili na inaweza kuhusishwa na kuzeeka ya seli hizi. Viumbe vya Eukaryotiki ikiwa ni pamoja na fungi na protozoans pia huzalisha telomerase ili kudumisha uadilifu wa kromos Kwa ugunduzi wake wa telomerase na hatua yake, Elizabeth Blackburn (1948—) alipokea Tuzo ya Nobel ya Tiba au Fiziolojia mwaka 2009.
| Mali | Bakteria | Eukaryotes |
|---|---|---|
| Muundo wa jenomu | Chromosome moja ya mviringo | Chromosomes nyingi za mstari |
| Idadi ya asili kwa chromosome | Single | Multiple |
| Kiwango cha kuiga | Nucleotides 1000 kwa pili | Nucleotides 100 kwa pili |
| Telomerase | Sio sasa | Sasa |
| RNA primer kuondolewa | DNA POL I | RNase H |
| Strand elongation | DNA pol III | pol δ, pol ε |
Zoezi\(\PageIndex{3}\)
- Je! Asili ya replication inatofautiana kati ya eukaryotes na prokaryotes?
- Nini enzymes za polymerase zinawajibika kwa awali ya DNA wakati wa replication ya eukaryotic?
- Ni nini kinachopatikana mwisho wa chromosomes katika eukaryotes na kwa nini?
Replication ya DNA ya Vipengele vya Extrachromosomal: Plasmids na Virusi
Ili kunakili asidi zao za nucleic, plasmidi na virusi mara nyingi hutumia tofauti juu ya muundo wa replication ya DNA iliyoelezwa kwa genomes za prokaryote. Kwa habari zaidi juu ya mikakati mbalimbali ya kuiga virusi, angalia Mzunguko wa Maisha ya Virusi.
Rolling mzunguko replication
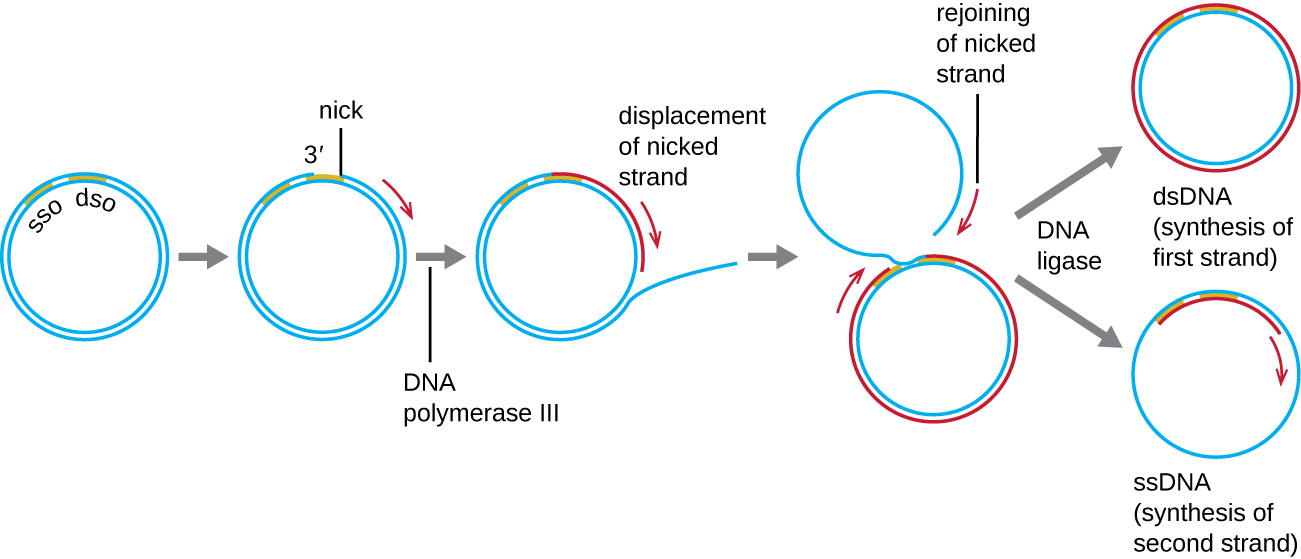
Wakati plasmidi nyingi za bakteria (tazama Tabia za kipekee za seli za Prokaryotiki) zinaiga kwa mchakato sawa na ule uliotumika kunakili kromosomu ya bakteria, plasmidi nyingine, bacteriophages kadhaa, na baadhi ya virusi vya eukaryotes hutumia uigaji wa mduara unaoendelea (Kielelezo\(\PageIndex{7}\)). Hali ya mviringo ya plasmids na circularization ya genomes baadhi ya virusi juu ya maambukizi hufanya hivyo iwezekanavyo. Rolling mduara replication huanza na nicking enzymatic ya strand moja ya molekuli mara mbili stranded mviringo katika asili mbili-stranded (DSO) tovuti. Katika bakteria, DNA polymerase III hufunga kwa kundi la 3'-OH la strand iliyopigwa na huanza kuiga DNA kwa njia moja kwa moja kwa moja kwa kutumia kamba isiyo na nicked kama template, kuhama kamba ya nicked kama inavyofanya hivyo. Kukamilika kwa replication DNA katika tovuti ya matokeo ya awali nick katika makazi yao kamili ya strand nicked, ambayo inaweza kisha recircularize katika moja stranded DNA molekuli. RNA primase kisha synthesizes primer kuanzisha replication DNA katika single-stranded asili (SSO) tovuti ya single-stranded DNA (ssDNA) molekuli, kusababisha mbili-stranded DNA (DSDNA) molekuli kufanana na nyingine mviringo DNA molekuli.
Zoezi\(\PageIndex{4}\)
Je! Kuna kamba iliyopungua katika replication ya mzunguko unaozunguka? Kwa nini au kwa nini?
Dhana muhimu na Muhtasari
- Mchakato wa kuiga DNA ni semiconservative, ambayo husababisha molekuli mbili za DNA, kila mmoja ana kamba moja ya wazazi wa DNA na kamba moja iliyopangwa.
- Katika bakteria, uanzishwaji wa replication hutokea katika asili ya replication, ambapo DNA supercoiled ni wazi na DNA gyrase, alifanya single-stranded na helixase, na amefungwa na single-stranded kisheria protini kudumisha yake moja - hali stranded. Primase huunganisha primer fupi ya RNA, kutoa kikundi cha bure cha 3'-OH ambacho DNA polymerase III inaweza kuongeza nucleotides ya DNA.
- Wakati wa upungufu, kamba inayoongoza ya DNA inatengenezwa kwa kuendelea kutoka kwa primer moja. Kamba ya lagging inatengenezwa kwa muda mfupi katika vipande vidogo vya Okazaki, kila mmoja anayehitaji primer yake mwenyewe. Vipindi vya RNA vinaondolewa na kubadilishwa na nucleotidi za DNA na na bakteria DNA polymerase I, na DNA ligase hufunga mapungufu kati ya vipande hivi.
- Kusitishwa kwa replication katika bakteria kunahusisha azimio la concatemers ya DNA ya mviringo kwa topoisomerase IV ili kutolewa nakala mbili za kromosomu ya mviringo.
- Eukaryotes kawaida huwa na chromosomes nyingi za mstari, kila mmoja na asili nyingi za kuiga. Kwa ujumla, replication katika eukaryotes ni sawa na kwamba katika prokaryotes.
- Hali ya mstari wa chromosomes ya eukaryotic inahitaji telomeres kulinda jeni karibu na mwisho wa chromosomes. Telomerase huongeza telomeres, kuzuia uharibifu wao, katika aina fulani za seli.
- Rolling mduara replication ni aina ya haraka unidirectional DNA awali ya molekuli mviringo DNA kutumika kwa ajili ya replication ya baadhi ya plasmidi.