
23.1: Asili ya Eukaryotic
- Page ID
- 176530

Ujuzi wa Kuendeleza
- Orodha ya sifa za kuunganisha za eukaryotes
- Eleza kile wanasayansi wanajua kuhusu asili ya eukaryotes kulingana na babu wa mwisho wa kawaida
- Eleza nadharia endosymbiotic
Mambo ya uhai huanguka katika makundi matatu makubwa: Archaea, Bakteria, na Eukarya. Wawili wa kwanza wana seli za prokaryotic, na ya tatu ina eukaryotes zote. Rekodi ndogo ya mafuta inapatikana ili kusaidia kutambua kile wanachama wa kwanza wa kila moja ya mistari hii walionekana kama, hivyo inawezekana kwamba matukio yote ambayo yalisababisha babu wa mwisho wa kawaida wa eukaryotes zilizopo zitabaki haijulikani. Hata hivyo, biolojia ya kulinganisha ya viumbe vilivyopo na rekodi ndogo ya visukuku hutoa ufahamu fulani katika historia ya Eukarya.
Fossils za mwanzo zilizopatikana zinaonekana kuwa Bakteria, uwezekano mkubwa wa cyanobacteria. Wao ni karibu na umri wa miaka bilioni 3.5 na wanatambulika kwa sababu ya muundo wao mgumu na, kwa prokaryotes, seli kubwa. Prokaryotes nyingine nyingi zina seli ndogo, 1 au 2 μm kwa ukubwa, na itakuwa vigumu kuchukua kama fossils. Eukaryotes walio hai zaidi wana seli za kupima 10 μm au zaidi. Miundo ukubwa huu, ambayo inaweza kuwa fossils, kuonekana katika rekodi ya kijiolojia kuhusu 2.1 miaka bilioni iliyopita.
Tabia za Eukaryotes
Takwimu kutoka kwa fossils hizi zimesababisha wanabiolojia wa kulinganisha na hitimisho kwamba eukaryotes wanaoishi wote ni wazao wa babu moja ya kawaida. Ramani sifa zilizopatikana katika makundi yote makubwa ya eukaryotes inaonyesha kwamba sifa zifuatazo lazima ziwepo katika babu wa mwisho wa kawaida, kwa sababu sifa hizi zipo katika angalau baadhi ya wanachama wa kila kizazi kikubwa.
- Viini vilivyo na viini vilivyozungukwa na bahasha ya nyuklia yenye pores Hii ni tabia moja ambayo ni muhimu na ya kutosha kufafanua kiumbe kama eukaryote. Eukaryotes zote zilizopo zina seli zilizo na nuclei.
- Mitochondria. Baadhi ya eukaryotes zilizopo zimepungua sana mabaki ya mitochondria katika seli zao, wakati wanachama wengine wa vizazi vyao wana “kawaida” mitochondria.
- Cytoskeleton iliyo na vipengele vya miundo na motility inayoitwa actin microfilaments na microtubules. Eukaryotes zote zilizopo zina vipengele hivi vya cytoskeletal.
- Flagella na cilia, organelles zinazohusiana na motility ya seli. Baadhi ya eukaryotes zilizopo hazina flagella na/au cilia, lakini zimetoka kwa mababu waliokuwa nao.
- Chromosomes, kila moja yenye molekuli linear DNA coiled kuzunguka msingi (alkali) protini iitwayo histones. Eukaryotes chache zilizo na chromosomes ambazo hazipo histones zimebadilika wazi kutoka kwa mababu waliokuwa nao.
- Mitosis, mchakato wa mgawanyiko wa nyuklia ambayo chromosomes kuigwa ni kugawanywa na kutengwa kwa kutumia mambo ya cytoskeleton. Mitosis iko duniani kote katika eukaryotes.
- Ngono, mchakato wa recombination maumbile kipekee kwa eukaryotes ambapo diploidi viini katika hatua moja ya mzunguko wa maisha hupata meiosis ili kuzalisha viini haploidi na karyogamy inayofuata, hatua ambapo viini viwili vya haploidi huunganisha pamoja ili kuunda kiini cha zygote cha diploidi.
- Wajumbe wa lineages yote makubwa wana kuta za seli, na inaweza kuwa na busara kuhitimisha kwamba babu ya kawaida ya mwisho inaweza kufanya kuta za seli wakati wa hatua fulani ya mzunguko wa maisha yake. Hata hivyo, haitoshi inajulikana kuhusu kuta za seli za eukaryotes na maendeleo yao kujua ni kiasi gani homolojia ipo kati yao. Ikiwa babu wa kawaida wa mwisho angeweza kufanya kuta za seli, ni wazi kwamba uwezo huu lazima umepotea katika vikundi vingi.
Endosymbiosis na Mageuzi ya Eukaryotes
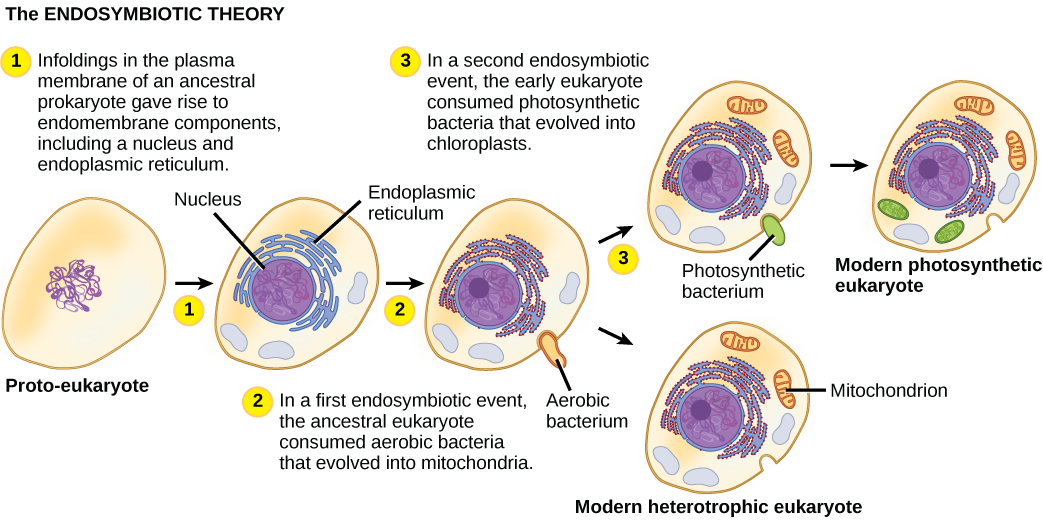
Ili kuelewa viumbe vya eukaryotic kikamilifu, ni muhimu kuelewa kwamba eukaryotes zote zilizopo ni kizazi cha kiumbe cha chimeric ambacho kilikuwa kikundi cha kiini cha jeshi na seli za alpha-proteobacterium ambazo “zilichukua makazi” ndani yake. Mandhari hii kuu katika asili ya eukaryotes inajulikana kama endosymbiosis, kiini kimoja kinachukua mwingine kama kwamba kiini kilichomwagika kinaendelea na seli zote mbili zinafaidika. Zaidi ya vizazi vingi, uhusiano wa ushirikiano unaweza kusababisha viumbe viwili vinavyotegemeana kabisa kwamba wala hakuweza kuishi peke yake. Matukio Endosymbiotic uwezekano imechangia asili ya babu ya mwisho ya kawaida ya eukaryotes leo na baadaye mseto katika lineages fulani ya eukaryotes (Kielelezo\(\PageIndex{4}\)). Kabla ya kuelezea hili zaidi, ni muhimu kuzingatia kimetaboliki katika prokaryotes.
Kimetaboliki ya Prokaryotic
Michakato mingi muhimu ya kimetaboliki iliondoka katika prokaryotes, na baadhi ya haya, kama vile fixation ya nitrojeni, haipatikani kamwe katika eukaryotes. Mchakato wa kupumua kwa aerobic hupatikana katika mistari yote makubwa ya eukaryotes, na ni localized katika mitochondria. Kupumua kwa aerobic pia hupatikana katika mistari mingi ya prokaryotes, lakini haipo katika wote, na aina nyingi za ushahidi zinaonyesha kwamba prokaryotes vile anaerobic kamwe kufanyika kupumua aerobic wala mababu zao.
Ilhali anga ya leo ni takriban oksijeni ya molekuli moja ya tano (O 2), ushahidi wa kijiolojia unaonyesha kuwa awali ilikosa O 2. Bila oksijeni, kupumua kwa aerobic hakutarajiwa, na vitu vilivyo hai vingetegemea fermentation badala yake. Wakati fulani kabla, karibu miaka bilioni 3.5 iliyopita, baadhi ya prokaryotes ilianza kutumia nishati kutoka jua ili kuimarisha michakato ya anabolic ambayo hupunguza dioksidi kaboni kuunda misombo ya kikaboni. Hiyo ni, walibadilisha uwezo wa photosynthesize. Hidrojeni, inayotokana na vyanzo mbalimbali, ilitekwa kwa kutumia athari zenye nguvu za mwanga ili kupunguza dioksidi kaboni iliyowekwa katika mzunguko wa Calvin. Kundi la bakteria ya Gramu-hasi lililotoa kupanda kwa cyanobacteria lilitumia maji kama chanzo cha hidrojeni na kutolewa O 2 kama bidhaa taka.
Hatimaye, kiasi cha oksijeni ya photosynthetic kilijengwa katika baadhi ya mazingira kwa viwango ambavyo vilikuwa hatari kwa viumbe hai, kwani inaweza kuharibu misombo mingi ya kikaboni. Michakato mbalimbali ya kimetaboliki ilibadilika ambayo inalinda viumbe kutoka oksijeni, moja ambayo, kupumua kwa aerobic, pia ilizalisha viwango vya juu vya ATP. Ilikuwa sasa sana kati ya prokaryotes, ikiwa ni pamoja na katika kikundi tunachoita sasa alpha-proteobacteria. Viumbe ambavyo havikupata kupumua kwa aerobic vilipaswa kubaki katika mazingira yasiyo ya oksijeni. Mwanzoni, mazingira yenye utajiri wa oksijeni yalikuwa yamewekwa karibu na maeneo ambapo cyanobacteria zilikuwa hai, lakini kwa karibu miaka bilioni 2 iliyopita, ushahidi wa kijiolojia unaonyesha kuwa oksijeni ilikuwa inajenga hadi viwango vya juu katika anga. Viwango vya oksijeni vinavyofanana na viwango vya leo viliondoka tu ndani ya miaka milioni 700 iliyopita.
Kumbuka kwamba fossils za kwanza ambazo tunaamini kuwa eukaryotes zimefikia umri wa miaka bilioni 2, hivyo zilionekana kama viwango vya oksijeni viliongezeka. Pia, kumbuka kwamba eukaryotes zote zilizopo zilishuka kutoka kwa babu na mitochondria. Organelles hizi zilionekana kwanza na microscopists nyepesi mwishoni mwa miaka ya 1800, ambapo zilionekana kuwa miundo fulani yenye umbo la minyoo iliyoonekana kuwa inazunguka ndani ya seli. Baadhi ya waangalizi wa mapema walipendekeza kuwa wanaweza kuwa bakteria wanaoishi ndani ya seli za jeshi, lakini nadharia hizi zilibakia haijulikani au kukataliwa katika jamii nyingi za kisayansi.
Theory Endosymbiotic
Kama biolojia ya seli ilivyotengenezwa katika karne ya ishirini, ikawa wazi kuwa mitochondria walikuwa organelles zinazohusika na kuzalisha ATP kwa kutumia kupumua aerobic. Katika miaka ya 1960, mwanabiolojia wa Marekani Lynn Margulis alianzisha nadharia endosymbiotiki, ambayo inasema kwamba eukaryotes huenda ikawa bidhaa ya seli moja inayojitokeza mwingine, moja akiishi ndani ya mwingine, na kubadilika baada ya muda hadi seli tofauti hazikutambulika tena kama vile. Mwaka 1967, Margulis alianzisha kazi mpya juu ya nadharia na kuimarisha matokeo yake kupitia ushahidi wa microbiological. Ingawa kazi ya Margulis awali ilikutana na upinzani, nadharia hii ya mara moja ya mapinduzi sasa imekubaliwa sana (lakini si kabisa), na kazi inaendelea kufunua hatua zinazohusika katika mchakato huu wa mabadiliko na wachezaji muhimu wanaohusika. Mengi bado yanabaki kugunduliwa kuhusu asili ya seli ambazo sasa zinaunda seli katika eukaryotes zote zilizo hai.
Kwa upana, imekuwa wazi kuwa wengi wa jeni zetu za nyuklia na mashine za Masi zinazohusika na kuiga na kujieleza zinaonekana karibu na wale walio katika Archaea. Kwa upande mwingine, organelles za kimetaboliki na jeni zinazohusika na michakato mingi ya kuvuna nishati zilikuwa na asili yao katika bakteria. Mengi bado kufafanuliwa kuhusu jinsi uhusiano huu ulitokea; hii inaendelea kuwa uwanja wa kusisimua wa ugunduzi katika biolojia. Kwa mfano, haijulikani kama tukio endosymbiotiki lililosababisha mitochondria ilitokea kabla au baada ya kiini cha jeshi kilikuwa na kiini. Viumbe vile vitakuwa miongoni mwa watangulizi wa mwisho wa babu wa kawaida wa eukaryotes.
Mitochondria
Moja ya sifa kuu za kutofautisha prokaryotes kutoka eukaryotes ni kuwepo kwa mitochondria. Seli za Eukaryotic zinaweza kuwa na mahali popote kutoka moja hadi mitochondria elfu kadhaa, kulingana na kiwango cha seli cha matumizi ya nishati. Kila mitochondrion hatua 1 hadi 10 au micrometers zaidi kwa urefu na ipo katika kiini kama organelle ambayo inaweza kuwa ovoid kwa minyoo umbo kwa intricately matawi (Kielelezo\(\PageIndex{1}\)). Mitochondria hutoka kutokana na mgawanyo wa mitochondria zilizopo; zinaweza kuunganishwa pamoja; na zinaweza kuhamishwa kuzunguka ndani ya seli kwa mwingiliano na cytoskeleton. Hata hivyo, mitochondria haiwezi kuishi nje ya seli. Kama anga ilikuwa oksijeni na usanisinuru, na kama prokaryotes aerobic mafanikio tolewa, ushahidi unaonyesha kwamba kiini mababu na compartmentalization utando engulfed free-hai aerobic prokaryote, hasa alpha-proteobacterium, na hivyo kutoa kiini jeshi uwezo wa kutumia oksijeni kutolewa nishati kuhifadhiwa katika virutubisho. Alpha-proteobacteria ni kundi kubwa la bakteria linalojumuisha spishi zinazofanana na mimea, viumbe vya magonjwa vinavyoweza kuwaambukiza binadamu kupitia kupe, na spishi nyingi za kuishi bure zinazotumia mwanga kwa nishati. Mistari kadhaa ya ushahidi unaunga mkono kwamba mitochondria inatokana na tukio hili la endosymbiotic. Mitochondria nyingi zina umbo kama alpha-proteobacteria na zimezungukwa na utando mbili, ambayo ingeweza kusababisha wakati kiumbe kimoja kilichofungwa kwa utando kilikuwa kikiingizwa ndani ya utupu na kiumbe kingine kilichofungwa kwa utando. Mbinu ya ndani ya mitochondrial ni pana na inahusisha infoldings kubwa inayoitwa cristae ambayo inafanana na uso wa nje wa alpha-proteobacteria. Matrix na utando wa ndani ni matajiri na enzymes zinazohitajika kwa kupumua kwa aerobic.
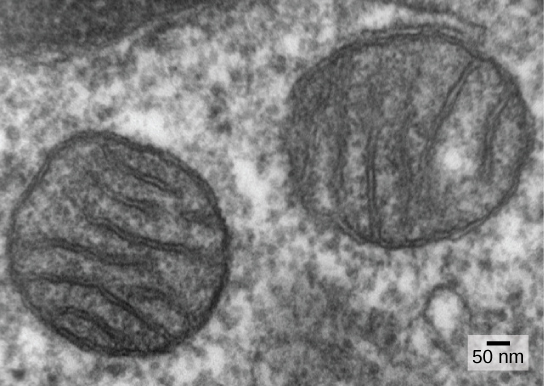
Mitochondria hugawanyika kwa kujitegemea na mchakato unaofanana na fission ya binary katika prokaryotes. Hasa, mitochondria haipatikani kutoka mwanzo (de novo) na seli ya eukaryotiki; huzaa ndani yake na husambazwa kwa saitoplazimu wakati kiini kinagawanyika au seli mbili zinafyuka. Kwa hiyo, ingawa organelles hizi zinaunganishwa sana katika kiini cha eukaryotiki, bado huzaa kana kwamba ni viumbe huru ndani ya seli. Hata hivyo, uzazi wao unafanana na shughuli na mgawanyiko wa seli. Mitochondria huwa na kromosomu yao ya DNA ya mviringo (kwa kawaida) ambayo imetulia na viambatisho kwenye utando wa ndani na hubeba jeni zinazofanana na jeni zilizoelezwa na alpha-proteobacteria. Mitochondria pia ina ribosomu maalum na RNAs za uhamisho zinazofanana na vipengele hivi katika prokaryotes. Hizi zina msaada wote ambao mitochondria walikuwa mara moja prokaryotes ya bure.
Mitochondria ambayo hufanya kupumua aerobic ina genomes yao wenyewe, na jeni sawa na wale walio katika alpha-proteobacteria. Hata hivyo, jeni nyingi za protini za kupumua ziko katika kiini. Wakati jeni hizi zikilinganishwa na zile za viumbe vingine, zinaonekana kuwa za asili ya alpha-proteobacterial. Zaidi ya hayo, katika baadhi ya vikundi vya eukaryotiki, jeni hizo hupatikana katika mitochondria, ambapo katika makundi mengine, hupatikana katika kiini. Hii imetafsiriwa kama ushahidi kwamba jeni zimehamishwa kutoka kromosomu endosymbiont hadi jenomu ya jeshi. Hasara hii ya jeni kwa endosymbiont pengine ni maelezo moja kwa nini mitochondria haiwezi kuishi bila jeshi.
Baadhi ya eukaryotes hai ni anaerobic na hawawezi kuishi mbele ya oksijeni nyingi mno. Baadhi huonekana kukosa organelles ambazo zinaweza kutambuliwa kama mitochondria. Katika miaka ya 1970 hadi mwanzoni mwa miaka ya 1990, wanabiolojia wengi walipendekeza kuwa baadhi ya eukaryotes hizi zilishuka kutoka kwa mababu ambao vizazi vilikuwa vimeachana na ukaryotes iliyo na mitochondrioni kabla endosymbiosis ilitokea. Hata hivyo, matokeo ya baadaye yanaonyesha kwamba organelles zilizopunguzwa zinapatikana katika wengi, ikiwa sio wote, eukaryotes anaerobic, na kwamba eukaryotes zote zinaonekana kubeba jeni fulani katika viini vyao ambavyo vina asili ya mitochondrial. Mbali na kizazi cha aerobic cha ATP, mitochondria ina kazi nyingine za kimetaboliki. Moja ya kazi hizi ni kuzalisha makundi ya chuma na sulfuri ambayo ni cofactors muhimu ya enzymes nyingi. Kazi hizo mara nyingi huhusishwa na organelles zilizopunguzwa za mitochondrion za eukaryotes anaerobic. Kwa hiyo, wanabiolojia wengi wanakubali kwamba babu wa mwisho wa eukaryotes alikuwa na mitochondria.
Plastidi
Makundi mengine ya eukaryotes ni photosynthetic. Seli zao zina, pamoja na organelles ya kawaida ya eukaryotic, aina nyingine ya organelle inayoitwa plastid. Wakati seli hizo zinafanya photosynthesis, plastidi zao zina matajiri katika chlorophyll ya rangi na rangi nyingine mbalimbali, inayoitwa rangi za vifaa, ambazo zinahusika katika kuvuna nishati kutoka mwanga. Plastidi za photosynthetic zinaitwa chloroplasts (Kielelezo\(\PageIndex{2}\)).
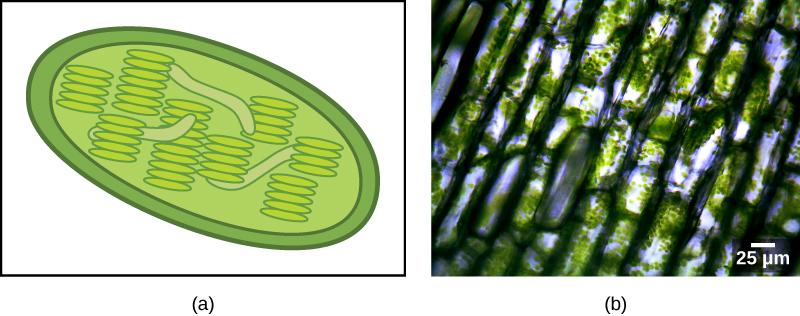
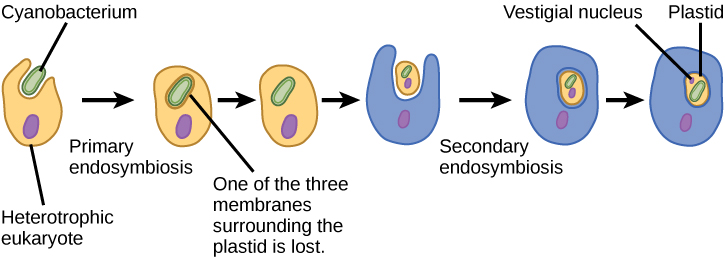
Kama mitochondria, plastids inaonekana kuwa na asili endosymbiotic. Nadharia hii pia ilipigwa na Lynn Margulis. Plastidi zinatokana na cyanobacteria zilizoishi ndani ya seli za eukaryote ya mababu, aerobic, heterotrophic. Hii inaitwa endosymbiosis ya msingi, na plastids ya asili ya msingi imezungukwa na membrane mbili. Ushahidi bora ni kwamba hii imetokea mara mbili katika historia ya eukaryotes. Katika hali moja, babu wa kawaida wa kizazi kikubwa/supergroup Archaeplastida alichukua endosymbiont cyanobacterial; kwa upande mwingine, babu wa taxon ndogo ya amoeboid rhizarian, Paulinella, alichukua endosymbiont tofauti ya cyanobacterial. Karibu eukaryotes zote za photosynthetic zinatoka kwenye tukio la kwanza, na aina kadhaa tu zinatokana na nyingine.
Cyanobacteria ni kundi la bakteria ya Gram-hasi na miundo yote ya kawaida ya kikundi. Hata hivyo, tofauti na prokaryotes nyingi, wana mifuko ya kina, ya ndani ya membrane inayoitwa thylakoids. Chlorophyll ni sehemu ya membrane hizi, kama vile protini nyingi za athari za mwanga za photosynthesis. Cyanobacteria pia ina ukuta wa peptidoglycan na safu ya lipopolysaccharide inayohusishwa na bakteria ya Gram-hasi.
Chloroplasts yenye asili ya msingi ina thylakoidi, kromosomu ya DNA ya mviringo, na ribosomu zinazofanana na zile za cyanobacteria. Kila chloroplast imezungukwa na membrane mbili. Katika kundi la Archaeplastida linaloitwa glaucophytes na katika Paulinella, safu nyembamba ya peptidoglycan iko kati ya utando wa nje na wa ndani wa plastiki. Plastidi nyingine zote hazina ukuta huu wa cyanobacterial wa relictual. Mbinu ya nje inayozunguka plastidi inadhaniwa kuwa imetokana na utupu katika jeshi, na utando wa ndani unadhaniwa kuwa unatokana na utando wa plasma wa symbiont.
Pia kuna, kama ilivyo kwa mitochondria, ushahidi mkubwa kwamba wengi wa jeni za endosymbiont zilihamishiwa kwenye kiini. Plastidi, kama mitochondria, haiwezi kuishi kwa kujitegemea nje ya mwenyeji. Aidha, kama mitochondria, plastidi zinatokana na mgawanyiko wa plastidi nyingine na kamwe hazijengwa kutoka mwanzo. Watafiti wamependekeza kuwa tukio la endosymbiotic ambalo limesababisha Archaeplastida lilitokea miaka bilioni 1 hadi 1.5 iliyopita, angalau miaka milioni mia 5 baada ya rekodi ya mafuta inaonyesha kuwa eukaryotes zilikuwepo.
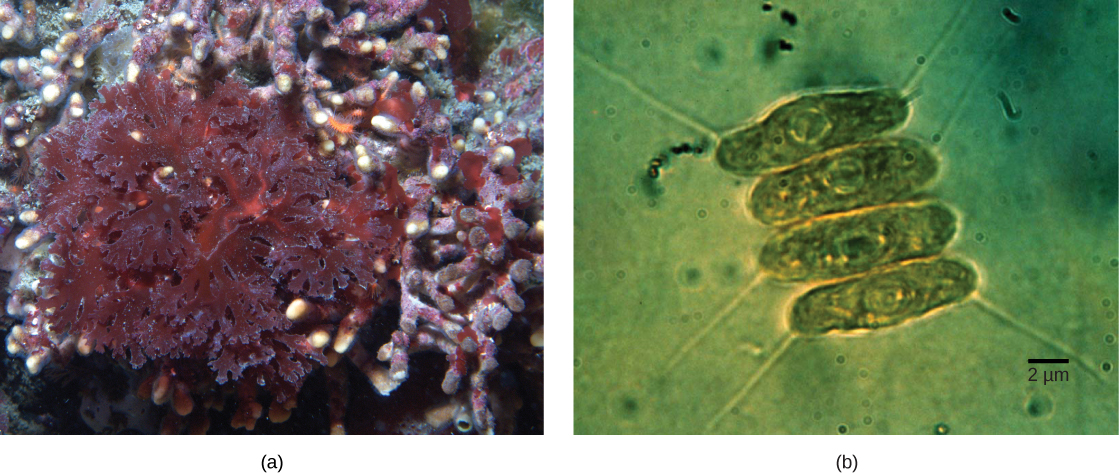
Sio plastidi zote katika eukaryotes zinazotokana moja kwa moja kutoka endosymbiosis ya msingi. Baadhi ya makundi makubwa ya mwani akawa photosynthetic na endosymbiosis ya sekondari, yaani, kwa kuchukua mwani wa kijani au nyekundu mwani (wote kutoka Archaeplastida) kama endosymbionts (Kielelezo\(\PageIndex{3}\)). Masomo mengi ya microscopic na maumbile yamesaidia hitimisho Plastidi ya sekondari imezungukwa na membrane tatu au zaidi, na baadhi ya plastiki za sekondari hata zina mabaki ya wazi ya kiini cha alga endosymbiotic. Wengine si “naendelea” mabaki yoyote. Kuna matukio ambapo matukio ya juu au ya juu-ili endosymbiotic ni maelezo bora kwa plastids katika baadhi ya eukaryotes.
Sanaa Connection
Kuna ushahidi gani kwamba mitochondria iliingizwa katika seli ya eukaryotic ya mababu kabla ya kloroplasts?
Uunganisho wa Mageuzi: Endosymbiosis ya Sekondari katika Chlorarachniophytes
Endosymbiosis inahusisha kiini kimoja kinachochochea mwingine ili kuzalisha, baada ya muda, uhusiano wa pamoja ambao hakuna kiini hakiwezi kuishi peke yake. Chloroplasts ya mwani nyekundu na kijani, kwa mfano, hutolewa kutokana na kuingizwa kwa cyanobacterium ya photosynthetic na prokaryote mapema.
Hii inasababisha swali la uwezekano wa kiini kilicho na endosymbiont yenyewe kinachopigwa, na kusababisha endosymbiosis ya sekondari. Ushahidi wa molekuli na maumbile unaonyesha kwamba protists ya klorarachniophyte yanatokana na tukio la endosymbiotic ya sekondari. Chlorarachniophytes ni wachache wa asili ya bahari ya kitropiki na mchanga ambayo inaweza kuhesabiwa katika supergroup ya rhizarian. Chlorarachniophytes hupanua vipande nyembamba vya cytoplasmic, kuunganisha wenyewe na chlorarachniophytes nyingine, kwenye mtandao wa cytoplasmic. Wafanyabiashara hawa wanafikiriwa kuwa wametokea wakati eukaryote ilipata alga ya kijani, ambayo mwisho ilikuwa tayari imeanzisha uhusiano wa endosymbiotic na cyanobacterium ya photosynthetic (Kielelezo\(\PageIndex{5}\)).
Mistari kadhaa ya ushahidi unaunga mkono kwamba chlorarachniophytes ilibadilika kutoka endosymbiosis ya sekondari. Chloroplasts zilizomo ndani ya endosymbionts ya kijani ya algal bado zina uwezo wa photosynthesis, na kufanya photosynthetes ya chlorarachniophytes. Endosymbiont ya kijani ya algal pia inaonyesha kiini kilichopigwa. Kwa kweli, inaonekana kwamba chlorarachniophytes ni bidhaa za tukio la mwisho la mwisho la mwisho la sekondari. Plastidi ya chlorarachniophytes imezungukwa na utando manne: mbili za kwanza zinahusiana na utando wa ndani na nje wa sainobacterium photosynthetic, ya tatu inalingana na alga ya kijani, na ya nne inalingana na utupu uliozungukwa na alga ya kijani wakati ilipokuwa imefungwa na chlorarachniophyte babu. Katika mistari mingine ambayo ilihusisha endosymbiosis ya sekondari, membrane tatu tu zinaweza kutambuliwa karibu na plastidi. Hii kwa sasa imerekebishwa kama kupoteza kwa usawa wa membrane wakati wa mageuzi.
Mchakato wa endosymbiosis ya sekondari sio ya kipekee kwa chlorarachniophytes. Kwa kweli, sekondari endosymbiosis ya mwani kijani pia imesababisha protists euglenid, wakati endosymbiosis sekondari ya mwani nyekundu imesababisha mageuzi ya dinoflagellates, apicomplexans, stramenopiles.
Muhtasari
Ushahidi wa kale wa mafuta ya eukaryotes ni karibu miaka bilioni 2. Fossils wakubwa kuliko haya yote huonekana kuwa prokaryotes. Inawezekana kwamba eukaryotes ya leo imeshuka kutoka kwa babu ambaye alikuwa na shirika la prokaryotic. Mzee wa mwisho wa kawaida wa Eukarya wa leo alikuwa na sifa kadhaa, ikiwa ni pamoja na seli zilizo na viini ambazo zimegawanyika mitotically na zilizomo kromosomu za mstari ambapo DNA ilihusishwa na histones, mfumo wa cytoskeleton na endometrembrane, na uwezo wa kufanya cilia/flagella wakati wa angalau sehemu ya maisha yake mzunguko. Ilikuwa aerobic kwa sababu ilikuwa na mitochondria iliyokuwa matokeo ya alpha-proteobacterium ya aerobic iliyoishi ndani ya kiini cha jeshi. Ikiwa mwenyeji huyu alikuwa na kiini wakati wa symbiosis ya awali bado haijulikani. Baba wa mwisho wa kawaida anaweza kuwa na ukuta wa seli kwa angalau sehemu ya mzunguko wa maisha yake, lakini data zaidi zinahitajika ili kuthibitisha hypothesis hii. Eukaryotes ya leo ni tofauti sana katika maumbo yao, shirika, mizunguko ya maisha, na idadi ya seli kwa kila mtu.
Sanaa Connections
Kielelezo\(\PageIndex{4}\): Ni ushahidi gani ambao mitochondria iliingizwa katika kiini cha eukaryotic cha mababu kabla ya kloroplasts?
- Jibu
-
Seli zote za eukaryotiki zina mitochondria, lakini si seli zote za eukaryotiki zina kloroplasts.
faharasa
- endosymbiosis
- kuingizwa kwa seli moja ndani ya nyingine kama vile seli iliyojaa huishi, na seli zote mbili zinafaidika; mchakato unaohusika na mageuzi ya mitochondria na chloroplasts katika eukaryotes
- nadharia endosymbiotic
- nadharia kwamba inasema kwamba eukaryotes inaweza kuwa bidhaa ya seli moja engulfing mwingine, moja hai ndani ya mwingine, na kutoa baada ya muda mpaka seli tofauti walikuwa tena kumtambua kama vile
- plastidi
- moja ya kundi la organelles zinazohusiana katika seli za mimea zinazohusika katika uhifadhi wa wanga, mafuta, protini, na rangi