
20.3: Mitazamo juu ya mti wa Phylogenetic
- Page ID
- 175354

Ujuzi wa Kuendeleza
- Eleza uhamisho wa jeni usio na usawa
- Eleza jinsi prokaryotes na eukaryotes huhamisha jeni kwa usawa
- Tambua mifano ya wavuti na pete ya mahusiano ya phylogenetic na kuelezea jinsi tofauti na dhana ya awali ya mti wa phylogenetic
Dhana za mfano wa phylogenetic zinabadilika mara kwa mara. Ni moja ya mashamba yenye nguvu zaidi ya utafiti katika biolojia yote. Zaidi ya miongo kadhaa iliyopita, utafiti mpya umepinga mawazo ya wanasayansi kuhusu jinsi viumbe vinavyohusiana. Mifano mpya ya mahusiano haya yamependekezwa kwa kuzingatiwa na jumuiya ya kisayansi.
Miti mingi ya phylogenetic imeonyeshwa kama mifano ya uhusiano wa mabadiliko kati ya spishi. Miti ya phylogenetic ilianza na Charles Darwin, ambaye alitengeneza mti wa kwanza wa phylogenetic mwaka 1837 (Kielelezo\(\PageIndex{1}\) a), ambayo ilikuwa kama mfano wa masomo ya baadaye kwa zaidi ya karne. Dhana ya mti phylogenetic yenye shina moja inayowakilisha babu wa kawaida, na matawi yanayowakilisha tofauti ya aina kutoka kwa babu hii, inafaa vizuri na muundo wa miti mingi ya kawaida, kama vile mwaloni (Kielelezo\(\PageIndex{1}\) b). Hata hivyo, ushahidi kutoka kwa uchambuzi wa kisasa wa mlolongo wa DNA na algorithms mpya zilizoendelea za kompyuta umesababisha wasiwasi kuhusu uhalali wa mfano wa mti wa kawaida katika jamii ya kisayansi.

Vikwazo kwa Mfano wa Classic
Classical kufikiri juu ya mageuzi prokaryotic, pamoja na katika mfano classic mti, ni kwamba aina kubadilika clonally. Hiyo ni, huzalisha watoto wenyewe na mabadiliko ya random tu yanayosababisha asili katika aina mbalimbali za aina za kisasa na za mwisho zinazojulikana kwa sayansi. Mtazamo huu ni kiasi fulani ngumu katika eukaryotes zinazozalisha ngono, lakini sheria za jenetiki za Mendelian zinaelezea tofauti katika watoto, tena, kuwa matokeo ya mabadiliko ndani ya spishi. Dhana ya jeni kuhamishwa kati ya spishi zisizohusiana haikuchukuliwa kama uwezekano hadi hivi karibuni. Uhamisho wa jeni usio na usawa (HT), unaojulikana pia kama uhamisho wa jeni lateral, ni uhamisho wa jeni kati ya spishi zisizohusiana. GT imeonyeshwa kuwa jambo la milele-sasa, huku wanafanyabiashara wengi wanaofanya jukumu kubwa kwa mchakato huu katika mageuzi, hivyo kuchanganya mfano wa mti rahisi. Jeni zimeonyeshwa kupitishwa kati ya spishi ambazo zinahusiana mbali tu kwa kutumia phylogeny ya kawaida, hivyo kuongeza safu ya utata kwa uelewa wa mahusiano ya phylogenetic.
Njia mbalimbali ambazo GT hutokea katika prokaryotes ni muhimu kuelewa phylogenies. Ingawa kwa sasa HGT haionekani kama muhimu kwa mageuzi ya eukaryotiki, GT haina kutokea katika uwanja huu pia. Hatimaye, kama mfano wa uhamisho wa mwisho wa jeni, nadharia za fusion ya jenomu kati ya viumbe vya symbiotic au endosymbiotic zimependekezwa kuelezea tukio la umuhimu mkubwa-mageuzi ya kiini cha kwanza cha eukaryotiki, bila ambayo wanadamu hawakuweza kuwepo.
Uhamisho wa Gene
Uhamisho wa jeni wa usawa (HGT) ni kuanzishwa kwa nyenzo za maumbile kutoka kwa spishi moja hadi aina nyingine kwa njia nyingine isipokuwa maambukizi ya wima kutoka kwa mzazi (wazazi) hadi watoto. Uhamisho huu huruhusu hata spishi zinazohusiana na mbali kugawana jeni, na kushawishi fenotipu zao. Inafikiriwa kuwa HGT imeenea zaidi katika prokaryotes, lakini kwamba tu asilimia 2 ya genome ya prokaryotic inaweza kuhamishwa na mchakato huu. Watafiti wengine wanaamini makadirio hayo ni mapema: umuhimu halisi wa HGT kwa michakato ya mageuzi lazima kutazamwa kama kazi inayoendelea. Kama jambo hilo linachunguzwa vizuri zaidi, linaweza kufunuliwa kuwa la kawaida zaidi. Wanasayansi wengi wanaamini kwamba GT na mutation huonekana kuwa (hasa katika prokaryotes) chanzo kikubwa cha tofauti za maumbile, ambayo ni malighafi kwa mchakato wa uteuzi wa asili. Uhamisho huu unaweza kutokea kati ya aina zozote mbili zinazoshiriki uhusiano wa karibu (Jedwali\(\PageIndex{1}\)).
Jedwali\(\PageIndex{1}\): Muhtasari wa Utaratibu wa Prokaryotic na Eukaryotic GT
| Mfumo | Njia ya Uhamisho | Mfano | |
|---|---|---|---|
| Prokaryotes | mabadiliko | matumizi ya DNA | prokaryotes nyingi |
| transduction | bacteriophage (virusi) | bakteria | |
| kunyambua | pilus | prokaryotes nyingi | |
| mawakala wa uhamisho wa jeni | chembe za page-kama | bakteria zambarau zisizo za | |
| Eukaryotes | kutoka kwa viumbe vya chakula | haijulikani | wadudu |
| kuruka jeni | transposons | mchele na mimea ya mtama | |
| epiphytes/vimelea | haijulikani | Yew mti fungi | |
| kutoka kwa maambukizi ya virusi |
GT katika Prokaryotes
Utaratibu wa GT umeonyeshwa kuwa wa kawaida kabisa katika nyanja za prokaryotiki za Bakteria na Archaea, kwa kiasi kikubwa kubadilisha jinsi mageuzi yao yanavyoonekana. Wengi wa mifano ya mabadiliko, kama vile katika Nadharia ya Endosymbiont, inapendekeza kwamba eukaryotes ilitoka kwa prokaryotes nyingi, ambayo inafanya GT kuwa muhimu zaidi kuelewa mahusiano ya phylogenetic ya aina zote zilizopo na za mwisho.
Ukweli kwamba jeni huhamishwa kati ya bakteria ya kawaida hujulikana kwa wanafunzi wa microbiolojia. Uhamisho huu wa jeni kati ya spishi ni utaratibu mkuu ambapo bakteria hupata upinzani dhidi ya antibiotics. Kwa kawaida, aina hii ya uhamisho imefikiriwa kutokea kwa njia tatu tofauti:
- Mabadiliko: DNA ya uchi inachukuliwa na bakteria
- Transduction: jeni ni kuhamishwa kwa kutumia virusi
- Muungano: matumizi ya tube mashimo iitwayo pilus kuhamisha jeni kati ya viumbe
Hivi karibuni, utaratibu wa nne wa uhamisho wa jeni kati ya prokaryotes umegunduliwa. Small, chembe virusi kama kuitwa gene uhamisho mawakala (GTAs) kuhamisha random genomic makundi kutoka aina moja ya prokaryote hadi nyingine. GTAs zimeonyeshwa kuwa na jukumu la mabadiliko ya maumbile, wakati mwingine kwa mzunguko wa juu sana ikilinganishwa na michakato mingine ya mabadiliko. GTA ya kwanza ilijulikana mwaka 1974 kwa kutumia bakteria zambarau, zisizo za sulfuri. GTA hizi, ambazo zinafikiriwa kuwa bacteriophages ambazo zilipoteza uwezo wa kuzaliana peke yao, hubeba vipande vya DNA random kutoka kiumbe kimoja hadi kingine. Uwezo wa GTAs kutenda kwa mzunguko wa juu umeonyeshwa katika masomo yaliyodhibitiwa kwa kutumia bakteria ya baharini. Matukio ya uhamisho wa jeni katika prokaryotes ya baharini, ama kwa GTA au kwa virusi, yamekadiriwa kuwa ya juu kama 10 13 kwa mwaka katika Bahari ya Mediteranea pekee. GTAs na virusi hufikiriwa kuwa magari ya HGT yenye ufanisi na athari kubwa juu ya mageuzi ya prokaryotic.
Kama matokeo ya uchambuzi huu wa kisasa wa DNA, wazo kwamba eukaryotes ilibadilika moja kwa moja kutoka Archaea imeshuka kwa neema. Ilhali eukaryotes hushirikisha vipengele vingi ambavyo havipo katika bakteria, kama vile sanduku la TATA (linalopatikana katika eneo la promota la jeni nyingi), ugunduzi kwamba baadhi ya jeni za eukaryotiki zilikuwa zenye homologi zaidi na DNA ya bakteria kuliko DNA ya Archaea kulifanya wazo hili liwe na uwezo mdogo. Zaidi ya hayo, fusion ya genomes kutoka Archaea na Bakteria na endosymbiosis imependekezwa kama tukio la mwisho katika mageuzi eukaryotic.
GT katika Eukaryotes
Ingawa ni rahisi kuona jinsi prokaryotes kubadilishana vifaa vya maumbile na GT, awali ilifikiriwa kuwa mchakato huu haukuwepo katika eukaryotes. Baada ya yote, prokaryotes ni seli moja tu zinazoonekana moja kwa moja kwa mazingira yao, wakati seli za ngono za viumbe vya multicellular kawaida hutolewa katika sehemu zilizohifadhiwa za mwili. Inafuata kutokana na wazo hili kwamba uhamisho wa jeni kati ya eukaryotes multicellular lazima iwe ngumu zaidi. Hakika, inadhaniwa kuwa mchakato huu ni wa kawaida katika eukaryotes na una athari ndogo sana ya mabadiliko kuliko katika prokaryotes. Licha ya ukweli huu, HGT kati ya viumbe vinavyohusiana na mbali imeonyeshwa katika aina kadhaa za eukaryotic, na inawezekana kwamba mifano zaidi itagunduliwa baadaye.
Katika mimea, uhamisho wa jeni umeonekana katika spishi ambazo haziwezi kuvuka pollinate kwa njia za kawaida. Transposons au “jenasi za kuruka” zimeonyeshwa kuhamisha kati ya mchele na aina za mimea ya mtama. Zaidi ya hayo, aina ya vimelea kulisha miti yew, ambayo dawa ya kupambana na kansa TAXOL® inatokana na gome, wamepata uwezo wa kufanya taxol wenyewe, mfano wazi wa uhamisho wa jeni.
Katika wanyama, mfano wa kuvutia sana wa GT hutokea ndani ya aina za aphid (Kielelezo\(\PageIndex{2}\)). Nguruwe ni wadudu ambao hutofautiana kwa rangi kulingana na maudhui ya carotenoid. Carotenoids ni rangi iliyofanywa na mimea mbalimbali, fungi, na microbes, na hutumikia kazi mbalimbali kwa wanyama, ambao hupata kemikali hizi kutoka kwa chakula chao. Binadamu zinahitaji carotenoids kuunganisha vitamini A, na tunawapata kwa kula matunda na mboga za machungwa: karoti, apricots, maembe, na viazi vitamu. Kwa upande mwingine, nyuzi za nyuzi zimepata uwezo wa kufanya carotenoids peke yao. Kulingana na uchambuzi wa DNA, uwezo huu ni kutokana na uhamisho wa jeni la vimelea ndani ya wadudu na GT, labda kama wadudu hutumiwa fungi kwa chakula. Enzyme carotenoid aitwaye desaturase ni wajibu wa rangi nyekundu kuonekana katika aphids fulani, na imekuwa zaidi umeonyesha kwamba wakati jeni hii ni inactivated na mutation, aphids kurejea nyuma rangi yao ya kawaida zaidi ya kijani (Kielelezo\(\PageIndex{2}\)).

Fusion ya Jenome na Mageuzi ya Eukaryotes
Wanasayansi wanaamini mwisho katika GT hutokea kwa njia ya fusion ya genome kati ya aina tofauti za prokaryotes wakati viumbe viwili vya symbiotic vinakuwa endosymbiotic. Hii hutokea wakati spishi moja inachukuliwa ndani ya saitoplazimu ya spishi nyingine, ambayo hatimaye husababisha jenomu yenye jeni kutoka kwa endosymbiont na mwenyeji. Utaratibu huu ni kipengele cha Nadharia ya Endosymbiont, ambayo inakubaliwa na wengi wa wanabiolojia kama utaratibu ambapo seli za eukaryotiki zilipata mitochondria na chloroplasts zao. Hata hivyo, jukumu la endosymbiosis katika maendeleo ya kiini ni zaidi ya utata. DNA ya nyuklia na mitochondrial inadhaniwa kuwa ya asili tofauti (tofauti) ya mabadiliko, huku DNA ya mitochondrial ikitokana na genomes za mviringo za bakteria zilizokuwa zimejaa seli za kale za prokaryotiki. DNA ya Mitochondrial inaweza kuonekana kama kromosomu ndogo zaidi. Kushangaza kutosha, DNA ya mitochondrial inarithi tu kutoka kwa mama. DNA ya mitochondrial inadhoofisha katika mbegu za kiume wakati mbegu inapoharibika katika yai ya mbolea au katika matukio mengine wakati mitochondria iliyoko flagellum ya mbegu inashindwa kuingia yai.
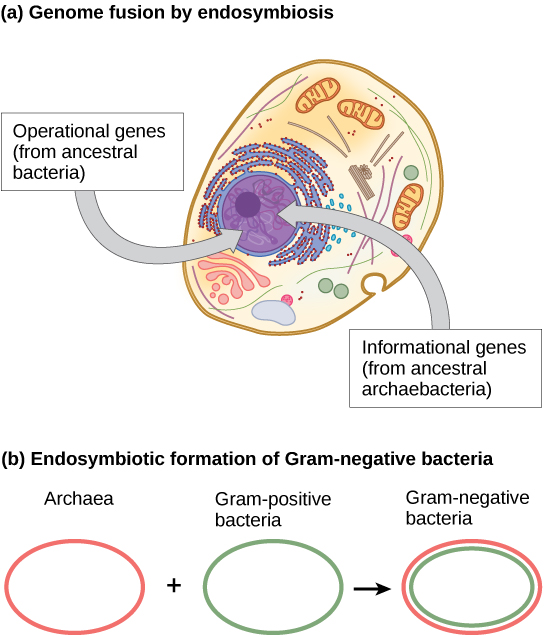
Katika kipindi cha miaka kumi iliyopita, mchakato wa fusion ya genome na endosymbiosis umependekezwa na James Ziwa la Taasisi ya Astrobiolojia ya UCLA/NASA kuwa na jukumu la mageuzi ya seli za kwanza za eukaryotic (Kielelezo\(\PageIndex{3}\) a). Kwa kutumia uchambuzi wa DNA na mpya hisabati algorithm iitwayo conditioned ujenzi (CR), maabara yake alipendekeza kwamba seli eukaryotic maendeleo kutoka endosymbiotic gene fusion kati ya aina mbili, moja Archaea na nyingine Bakteria. Kama ilivyoelezwa, baadhi ya jeni za eukaryotiki zinafanana na zile za Archaea, ilhali nyingine zinafanana na zile za Bakteria. Tukio la fusion endosymbiotic, kama vile Ziwa limependekeza, lingeelezea wazi uchunguzi huu. Kwa upande mwingine, kazi hii ni mpya na algorithm ya CR haipatikani, ambayo husababisha wanasayansi wengi kupinga hypothesis hii.
Kazi ya hivi karibuni zaidi na Ziwa (Kielelezo\(\PageIndex{3}\) b) inapendekeza kwamba bakteria gramu-hasi, ambayo ni ya kipekee ndani ya uwanja wao kwa kuwa wao vyenye mbili lipid bilayer utando, kwa kweli matokeo ya fusion endosymbiotic ya archaeal na aina ya bakteria. Mbinu mbili itakuwa matokeo ya moja kwa moja ya endosymbiosis, na endosymbiont kuokota utando wa pili kutoka kwa mwenyeji kama ilivyokuwa ndani. Utaratibu huu pia umetumika kuelezea utando wa mara mbili unaopatikana katika mitochondria na chloroplasts. Kazi ya Ziwa haipo na wasiwasi, na mawazo bado yanajadiliwa ndani ya jamii ya sayansi ya kibaiolojia. Mbali na nadharia tete za Ziwa, kuna nadharia nyingine kadhaa zinazoshindana kuhusu asili ya eukaryotes. Kiini cha eukaryotic kilibadilikaje? Nadharia moja ni kwamba seli za prokaryotiki zilizalisha utando wa ziada uliozunguka kromosomu ya bakteria. Baadhi ya bakteria wana DNA iliyoambatanishwa na utando mbili; hata hivyo, hakuna ushahidi wa nucleolus au pores nyuklia. Proteobacteria nyingine pia zina kromosomu zinazofungwa kwa membrane. Ikiwa kiini cha eukaryotic kilibadilika kwa njia hii, tunatarajia moja ya aina mbili za prokaryotes kuwa karibu zaidi na eukaryotes.
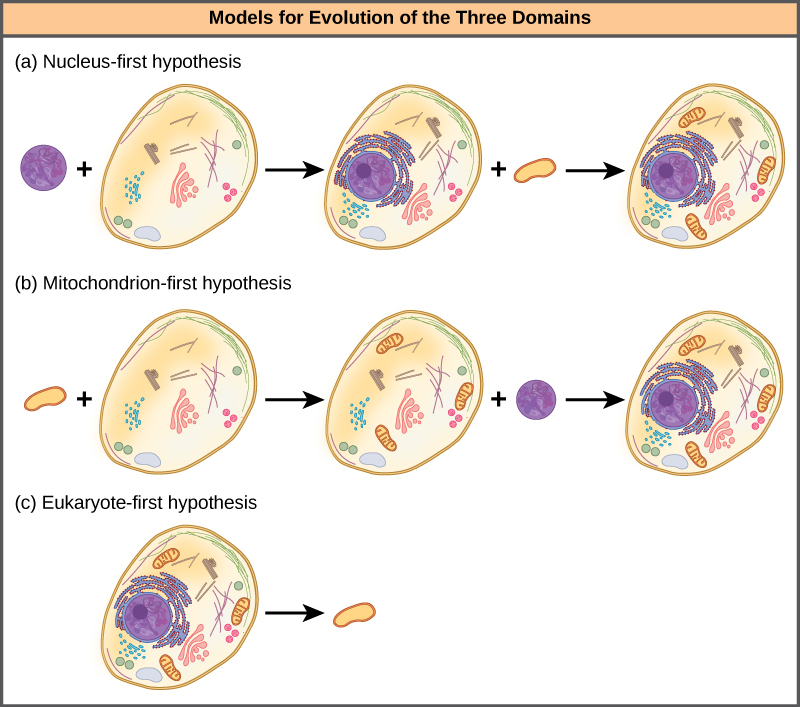
Nucleus-kwanza hypothesis inapendekeza kwamba kiini ilibadilika katika prokaryotes kwanza (Kielelezo\(\PageIndex{4}\) a), ikifuatiwa na fusion baadaye ya eukaryote mpya na bakteria ambayo ikawa mitochondria. Nadharia ya kwanza ya mitochondria inapendekeza kwamba mitochondria ilianzishwa kwanza katika jeshi la prokaryotic (Kielelezo\(\PageIndex{4}\) b), ambayo hatimaye ilipata kiini, kwa fusion au njia nyingine, kuwa kiini cha kwanza cha eukaryotic. Zaidi ya kushangaza, hypothesis eukaryote-kwanza inapendekeza kwamba prokaryotes kweli tolewa kutoka eukaryotes kwa kupoteza jeni na utata (Kielelezo\(\PageIndex{4}\) c). Hadithi hizi zote zinaweza kupimwa. Wakati tu na majaribio zaidi itaamua ambayo hypothesis ni bora mkono na data.
Mifano ya Mtandao na Mtandao
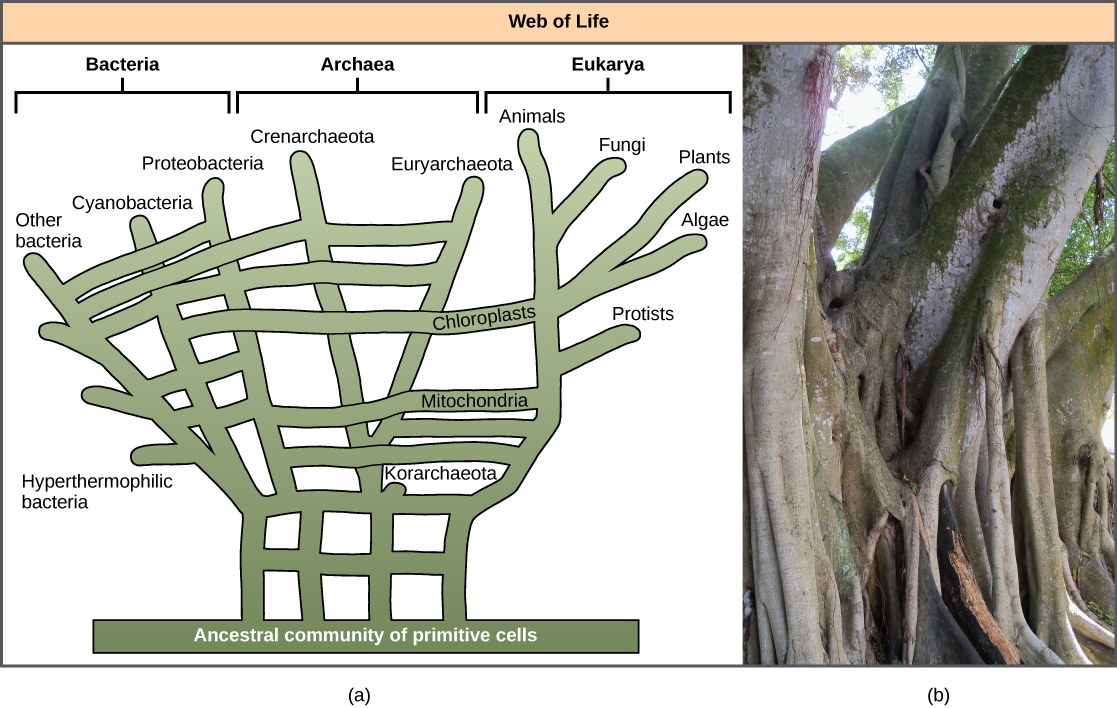
Kutambua umuhimu wa GT, hasa katika mageuzi ya prokaryotes, imesababisha wengine kupendekeza kuacha mfano wa “mti wa uzima” wa kawaida. Mwaka 1999, W. Ford Doolittle alipendekeza mfano wa phylogenetic unaofanana na mtandao au mtandao zaidi ya mti. Nadharia tete ni kwamba eukaryotes hazibadilika kutoka kwa babu mmoja wa prokaryotiki, bali kutoka kwenye bwawa la spishi nyingi zilizokuwa zikishirikiana jeni kwa njia za HT. Kama inavyoonekana katika Kielelezo\(\PageIndex{5}\) a, baadhi prokaryotes mtu binafsi walikuwa na jukumu la kuhamisha bakteria ambayo yalisababisha maendeleo ya mitochondrial kwa eukaryotes mpya, ambapo aina nyingine kuhamishiwa bakteria kwamba alitoa kupanda kwa chloroplasts. Mfano huu mara nyingi huitwa “mtandao wa maisha.” Kwa jitihada za kuokoa mlinganisho wa mti, wengine wamependekeza kutumia mti wa Ficus (Kielelezo\(\PageIndex{5}\) b) na vigogo vyake vingi kama phylogenetic ili kuwakilisha jukumu la mabadiliko ya kupungua kwa GT.
Gonga la Mifano ya Maisha
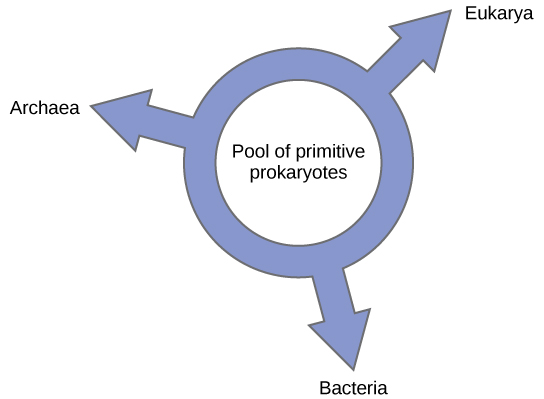
Wengine wamependekeza kuacha mfano wowote wa mti wa phylogeny kwa ajili ya muundo wa pete, kinachojulikana kama “pete ya maisha” (Kielelezo\(\PageIndex{6}\)); mfano wa phylogenetic ambapo nyanja zote tatu za maisha zilibadilishwa kutoka kwenye bwawa la prokaryotes za kale. Ziwa, tena kwa kutumia conditioned ujenzi algorithm, inapendekeza mfano pete-kama ambayo aina ya Domains zote tatu-Archaea, Bakteria, na Eukarya-tolewa kutoka pool moja ya prokaryotes gene-swapping. Maabara yake inapendekeza kwamba muundo huu ni fit bora kwa data kutoka uchambuzi wa kina DNA uliofanywa katika maabara yake, na kwamba mtindo pete ni moja tu kwamba kutosha inachukua HGT na genomic fusion katika akaunti. Hata hivyo, phylogeneticists wengine bado wana wasiwasi juu ya mfano huu.
Kwa muhtasari, mfano wa “mti wa uzima” uliopendekezwa na Darwin lazima ubadilishwe kuwa pamoja na GT. Je! Hii inamaanisha kuacha mfano wa mti kabisa? Hata Ziwa anasema kuwa majaribio yote yanapaswa kufanywa kugundua baadhi ya mabadiliko ya mtindo wa mti ili kuruhusu kwa usahihi kufaa data yake, na tu kutokuwa na uwezo wa kufanya hivyo itakuwa sway watu kuelekea pete pendekezo lake.
Hii haimaanishi mti, wavuti, au pete itahusiana kabisa na maelezo sahihi ya mahusiano ya phylogenetic ya maisha. Matokeo ya mawazo mapya juu ya mifano ya phylogenetic ni wazo kwamba mimba ya awali ya Darwin ya mti wa phylogenetic ni rahisi sana, lakini ilikuwa na maana kulingana na kile kilichojulikana wakati huo. Hata hivyo, utafutaji wa mfano muhimu zaidi unaendelea: kila mfano hutumikia kama nadharia ya kupimwa na uwezekano wa kuendeleza mifano mpya. Hii ni jinsi sayansi inavyoendelea. Mifano hizi hutumiwa kama visualizations kusaidia kujenga mahusiano ya mabadiliko ya nadharia na kuelewa kiasi kikubwa cha data kuwa kuchambuliwa.
Muhtasari
Mti wa phylogenetic, uliotumiwa kwanza na Darwin, ni mfano wa “mti wa uzima” wa kawaida unaoelezea mahusiano ya phylogenetic kati ya aina, na mfano wa kawaida unaotumiwa leo. Mawazo mapya kuhusu GT na fusion ya genome yamesababisha baadhi kupendekeza kurekebisha mfano ili kufanana na webs au pete.
faharasa
- eukaryote-kwanza hypothesis
- pendekezo kwamba prokaryotes tolewa kutoka eukaryotes
- wakala wa uhamisho wa jeni (GTA)
- chembe ya bacteriophage-kama ambayo huhamisha makundi ya genomic ya random kutoka kwa aina moja ya prokaryote hadi nyingine
- fusion ya jenomu
- fusion ya genomes mbili prokaryotic, labda kwa endosymbiosis
- usawa gene uhamisho (HGT)
- (pia, lateral gene uhamisho) uhamisho wa jeni kati ya aina unrelated
- mitochondria-hypothesis ya kwanza
- pendekezo kwamba prokaryotes alipewa mitochondrion kwanza, ikifuatiwa na maendeleo ya nyuklia
- nucleus-kwanza nadharia
- pendekezo kwamba prokaryotes alipewa kiini kwanza, na kisha mitochondrion
- pete ya maisha
- phylogenetic mfano ambapo nyanja zote tatu za maisha tolewa kutoka bwawa la prokaryotes primitive
- mtandao wa maisha
- phylogenetic mfano kwamba majaribio ya kuingiza madhara ya uhamisho usawa gene juu ya mageuzi